Immunmonitoring und Immuntherapie mit HIV-spezifischen mrnas
|
|
|
- Timo Koch
- vor 8 Jahren
- Abrufe
Transkript
1 Immunmonitoring und Immuntherapie mit HIV-spezifischen mrnas Der Naturwissenschaftlichen Fakultät der Friedrich-Alexander-Universität Erlangen-Nürnberg zur Erlangung des Doktorgrades Dr. rer. nat. vorgelegt von Jennifer Etschel aus Ansbach

2 Als Dissertation genehmigt von der Naturwissenschaftlichen Fakultät der Friedrich-Alexander-Universität Erlangen-Nürnberg Tag der mündlichen Prüfung: Vorsitzender der Prüfungskommission: Prof. Dr. Johannes Barth Erstberichterstatter: Prof. Dr. Thomas Winkler Zweitberichterstatter: Prof. Dr. Thomas Harrer

3 Inhaltsverzeichnis 1 ZUSAMMENFASSUNG / SUMMARY... 1 Zusammenfassung... 1 Summary EINLEITUNG Humanes Immundefizienz Virus (HIV) Epidemiologie und Tropismus Pathogenese und Klinik der HIV-1-Infektion Aufbau und Replikationszyklus Immunologische und genetische Faktoren, die die HIV-1-Infektion und Progression beeinflussen Zytotoxische T-Lymphozyten (CTL) HLA-Allele Toll-like Rezeptoren in der angeborenen Immunantwort CD4 + T-Helferzellen Genetische Unterschiede im Gen des Chemokinrezeptor CCR Interaktion von micrornas und Viren Therapie und Behandlungsansätze von HIV Hochaktive antiretrovirale Therapie (HAART) Vakzine Verwendung von mrna als Vakzine ZIELSETZUNG ERGEBNISSE Elektroporation von PBMC mit HIV-1-spezifischen mrnas Erfolgreiche Generierung von HIV-1-spezifischen mrnas Test und Titration von HIV-1-spezifischen Antikörpern in transfizierten HEK293T Zellen Elektroporation der HIV-1-spezifischen mrnas zeigt schnelle und effiziente Proteinexpression PBMC exprimieren HIV-1-Proteine bis zu sechs Tagen nach Elektroporation mrna-kodierte HIV-1-Proteine sind in vitro immunogen und führen zur Freisetzung von γ- IFN I
... 17 2.3.2 Vakzine... 18 2.3.3 Verwendung von mrna als Vakzine... 20 3 ZIELSETZUNG.")
4 Inhaltsverzeichnis Das autologe Virus löst in fünf von zehn Fällen eine stärkere Immunantwort aus als Referenzviren Vergleich der ausgelösten T-Zellantworten zwischen mrna-kodierten HIV-1-Proteinen und Peptiden, Peptid-Pools und Vaccinia-Viren mrna-kodierte gp120-proteine induzieren geringe Immunantworten Geringe Mengen an HIV-1-spezifischer mrna reichen aus, um eine Proteinexpression und T-Zellstimulation zu erhalten Durch mrna-kodierte HIV-1-Proteine aktivieren HIV-1-spezifische CD8 + zytotoxische T-Lymphozyten ohne zusätzliche Effektorzellen Neben γ-ifn induzieren die von HIV-1-spezifischen mrnas-kodierten Proteine die Sekretion von TNF-α Funktionelle Analysen mit HIV-1-spezifischen mrnas mrna-kodierte HIV-1-Proteine stimulieren die Generierung von HIV-1-spezifischen CTL Linien PBMC proliferieren nicht spezifisch nach Elektroporation mit mrna Mit HIV-1-spezifischer mrna-elektroporierte PBMC zeigen nach Kryokonservierung noch schwache Immunantworten mrna-kodierte Nef-Proteine reduzieren die Expression von CD4, CXCR4 und HLA-I Folgen der simultanen Elektroporation von gag und nef mrna Die simultane Elektroporation zweier HIV-1-spezifischer mrnas in B-Zelllinien hat keine Auswirkungen Die simultane Elektroporation zweier HIV-1-spezifischer mrnas in PBMC führt zu einem Verlust der Immunantworten Mit mrna-elektroporierten PBMC kann die CTL-Epitop-Hierarchie studiert werden Kreuzreaktion HIV-1-spezifischer CTLs mit humanen Proteinen Das Nef-Epitop TM9 ähnelt einem Epitop im humanen Protein Fat Breite CTL-Antwort gegen die Epitope Nef TM9 und Fat3 TM9 T/P pfat3 mrna Elektroporation führte trotz effizienter Proteinexpression zu keiner Erkennung durch Nef-spezifische CTLs In silico Analysen des fat3 mrna-konstrukts sagen eine Prozessierung des Epitops TM9 T/P durch das Proteasom mit einer 50%igen Wahrscheinlichkeit voraus Das Nef-Epitop Nef7v1 ist einem Epitop im humanen Enzym D-Aminosäuren-Oxidase (DAO) ähnlich Analyse und Screening von neuen und bekannten CTL-Epitopen Definition von drei neuen Gag und zwei neuen Nef CTL-Epitopen Analyse der neu definierten CTL-Epitope Das Peptid Gag enthält ein HLA-A*02-restringiertes CTL-Epitop Das Peptid Gag07-02 enthält ein HLA-B*41-restringiertes CTL-Epitop II

5 Inhaltsverzeichnis Die restringierenden HLA-Allele für die Peptide Gag07-11 und Gag07-75 konnten nicht bestimmt werden Die Peptide Nef07-08 und Nef07-10 enthalten HLA-B*41-Epitope Das bekannte HLA-C-restringierte Nef-Epitop Nef11T3 ist ein häufig erkanntes Epitop Etablierung alternativer Transfektionsmethoden Chitosan transfiziert DNA, aber keine mrna in HEK293T HIV-1 Tat führt zu keinem Transfer von Nukleinsäuren in Zellen Atelokollagen transfiziert weder DNA noch mrna in HEK293 und HEK293T FuGENE kann neben DNA auch mrna erfolgreich in HEK293T Zellen transfizieren Analyse des Sequenzverlaufs und der Immunantworten eines HIV-1-infizierten Patientenpaares, das mit demselben Virus infiziert worden war Unterschiedliche Erkennung der homologen und heterologen viralen mrna-konstrukten Die viralen autologen Sequenzen erklären die detektierten Immunantworten In Patient 0225 mutierte das Virus im selben Zeitraum deutlich mehr als in Patient Interaktion zwischen HIV-1-spezifischen mrnas und zellulären mirnas Einzelne HIV-1-Gene beeinflussen das zelluläre mirna-profil in CD4 + T-Zellen Durch Deep Sequencing konnten zelluläre mirnas identifiziert werden, deren Expressionsstärke von einzelnen HIV-1-Genen verändert wird Durch HIV-1-Proteine modifizierte mirnas konnten teilweise in einzelnen Proben durch qrt-pcr validiert werden CTL Fluchtmutationen in mirna-binderegionen beeinflussen die Regulation von mirnas DISKUSSION Elektroporation von PBMC mit HIV-1-spezifischen mrnas Immunmonitoring gegen autologe Virusproteine mit HIV-1-spezifischen mrnas HIV-1-spezifische mrnas als therapeutischer Impfstoff Funktionelle Analysen mit HIV-1-spezifischen mrnas Wirksamkeit eines Kombinationsimpfstoffes aus gag und nef mrna Studien zur CTL-Epitop-Hierarchie identifizieren effizient prozessierte Epitope als mögliche Vakzin-Antigene Ausschluss von Nebenwirkungen bei einer Impfung Wechselwirkung zwischen HIV-1-spezifischen CTLs und dem humanen Protein Fat Wechselwirkung zwischen HIV-1-spezifischen CTLs und dem humanen Protein DAO Analyse und Definition bekannter und neuer CTL-Epitope III

6 Inhaltsverzeichnis 5.5 Etablierung alternativer Transfektionsmethoden von mrna Transfektion mit Chitosan Transfektion über das HIV-1 Tat-Protein Transfektion mit Hilfe von Atelokollagen Transfektion mit FuGENE Evolution eines HI-Ausgangsviruses in zwei unterschiedlichen Patienten und die daraus resultierenden Immunantworten Vergleichende Immunantworten auf die mrna-kodierten HIV-1-Proteine Evolution eines HI-Virus in zwei unterschiedlichen Patienten Interaktion von HIV-1-spezifischen mrnas und zellulären mirnas Einfluss der HIV-1 Gene gag und nef auf mirnas in CD4 + T-Zellen Einfluss von CTL Fluchtmutanten in Nef auf die mir-29a-interaktion Ausblick MATERIAL UND METHODEN Material Patienten Verbrauchsmateralien Laborgeräte Chemikalien Puffer und Lösungen Kulturmedien und Zusätze Peptide und Proteine Antikörper Kommerziell erhältliche Kits Enzyme Längenstandards für Gelelektrophoresen Vektoren Bakterien, Viren, Zellen und Zelllinien Oligonukleotidprimer TaqMan MicroRNA Assays Datenbanken Software Methoden Molekularbiologische Methoden Isolation von Nukleinsäuren Konzentrationsbestimmung von Nukleinsäuren IV

7 Inhaltsverzeichnis Polymerasen-Ketten-Reaktion (PCR) Agarosegelelektrophorese Sequenzierung Klonierungsmethoden Generierung von mrna Hochdurchsatzsequenzierung (Deep Sequencing) Quantitative Echtzeit (Real-time) PCR (qrt-pcr) HLA Typisierung Zellbiologische Methoden Allgemeine Hinweise für Zellkulturverfahren Zellselektion Transfektionsmethoden mrna-elektroporation Durchflusszytometrie Untersuchung der Vitalität von Zellen Bestimmung der Zellproliferation Lösen und Verdünnen von Peptide γ-interferon ELISPOT Analyse der Freisetzung von weiteren Zytokine Dualer Luciferase-Reporter-Assay Infektion mit rekombinanten Vaccinia-Vektoren LITERATURVERZEICHNIS ABKÜRZUNGSVERZEICHNIS EIGENE PUBLIKATIONEN, KONFERENZBEITRÄGE UND VORTRÄGE DANKSAGUNG V

8

9 1 Zusammenfassung / Summary Zusammenfassung Ein wesentlicher Schwerpunkt der Promotionsarbeit waren Experimente zur Entwicklung von viralen mrna-konstrukten zum Immunmonitoring von T-Zellen und zur Entwicklung von Vakzinen. In der vorgelegten Dissertation wurden HIV-1-spezifische mrnas generiert und erfolgreich zur Elektroporation in PBMC eingesetzt. Es konnte dabei gezeigt werden, dass die Elektroporation von PBMC mit autologen HIV-1-spezifischen mrnas eine schnelle und effiziente Analyse der T-Zellantworten gegen das eigene HI-Virus erlaubt. Es konnte eine schnelle und effiziente Expression der mrna-kodierten Proteine in allen Subpopulationen der PBMC nachgewiesen werden, wobei die Proteinexpression bis zu sechs Tage nach Elektroporation detektiert werden konnte. Die mrna-kodierten HIV-1- Proteine waren in vitro hoch immunogen und lösten die Sekretion von γ-ifn und TNF-α aus. Dabei zeigte sich, dass die γ-ifn Freisetzung von HIV-1-spezifischen T-Zellen verursacht wird. Neben der Immunogenität wurde auch die Funktionalität der mrnakodierten HIV-1-Proteine belegt. Denn die Expression der Moleküle CD4, CXCR4 und HLA-I auf der Zelloberfläche wurde durch die mrna-kodierten Nef-Proteine deutlich reduziert. Außerdem konnten mrna-kodierte HIV-1-Proteine CD8 + T-Zellen aktivieren. Die mrna-kodierten HIV-1-Proteine können als alternative Antigene für das Monitoring von T-Zellantworten bei HIV-1-infizierten Patienten eingesetzt werden. Sie induzierten stärkere CTL-Antworten wie rekombinante Vaccinia-Viren und ähnliche Immunreaktionen wie Peptid-Pools. Im Vergleich zu einzelnen HIV-1-Peptiden rufen sie jedoch geringere T-Zellantworten hervor. Durch den Einsatz autologer HIV-1-spezifischer mrnas konnten die Immunantworten gegen autologe Virusproteine detektiert und mit denen gegen heterologe Virusproteine verglichen werden. In fünf von zehn Fällen lösten die autologen viralen Proteine stärkere Immunantworten aus als Proteine von HIV-1 Referenzviren. Folglich konnte bewiesen werden, dass die im Monitoring ermittelten Immunreaktionen häufig unterschätzt werden, da mit den Standard-Methoden die CTL- Antworten gegen das eigene Virus nicht quantifiziert werden können. In zwei Fällen wurde eine deutlich geringere T-Zellantwort gegen das autologe Virusprotein gemessen, was auf CTL Fluchtmutanten in der autologen Virussequenz zurückgeführt werden konnte. Demzufolge kann es auch zur Überschätzung der Immunantworten beim Standard- Monitoring kommen. Neben der Etablierung einer neuen, effizienten Methode zum Monitoring von T-Zellantworten gegen autologe Viren diente die mrna-elektroporation auch zur Aufklärung wichtiger Fragestellungen, die den Einsatz HIV-1-spezifischer mrnas als 1

10 1 Zusammenfassung / Summary therapeutische Vakzine betreffen. Es konnte bestätigt werden, dass die mrna-kodierten Proteine Gag und Nef in HIV-1-infizierten Patienten besser erkannt wurden als Env. Allerdings konnte gezeigt werden, dass die gleichzeitige Verabreichung von gag und nef mrna zu einem drastischen Verlust der induzierten Immunantworten in PBMC aufgrund der HLA-I-Reduktion durch das Nef-Protein führt. Zudem wurden klare Hinweise erhalten, aus welchen Epitopen eine optimale mrna-vakzine bestehen soll, da mit dem entwickelten Assay die CTL-Epitop-Hierarchie studiert werden konnte. Außerdem wurde von mir untersucht, ob Kreuzreaktionen von HIV-1-spezifischen CTLs mit humanen Proteinen existieren, die zu Nebenwirkungen bei einer mrna-vakzinierung führen könnten. Dazu wurden Nef-spezifische CTL auf die Erkennung von Peptiden aus den humanen Proteinen DAO und Fat3 getestet. Dabei zeigte sich eine deutliche Erkennung der Peptide, jedoch keine Erkennung von Fat3-exprimierenden Zelllinien. Im Rahmen der Promotion konnte ich mehrere CTL-Epitope neu definieren, darunter drei neue CTL-Epitope in Gag und zwei in Nef. Da HIV-1 Nef HLA-A und B, nicht jedoch HLA-C von der Zelloberfläche herabreguliert und damit die HIV-1-spezifische CTL-Antwort inhibiert, gewinnt die HLA-C-restringierte CTL-Antwort an Bedeutung. Daher wurde eine systematische Analyse der Erkennung eines konservierten HLA-Cw-restringierten CTL in Nef durchgeführt, wobei eine Erkennung des Epitops von ca. der Hälfte der untersuchten HLA-Cw*07-positiven HIV-1-Patienten festgestellt werden konnte. Da bei einer in vivo Applikation die mrna-elektroporation eher ungeeignet ist, wurde nach alternativen Transfektionsmethoden gesucht. Dabei konnte mrna mit Hilfe von FuGENE in die Zelle transfiziert werden, wohingegen Chitosan, Atelokollagen und das HIV-1 Tat-Protein nicht in der Lage waren, mrna in die Zelle einzuschleusen. Um den Einfluss von viralen Proteinen auf die Expression von zellulären mirnas zu erfassen, wurden die HIV-1 gag und nef mrnas in CD4 + T-Zellen elektroporiert und das mirna-profil analysiert. Die gewonnenen Daten weisen darauf hin, dass virale Gene aus verschiedenen HIV-1-Patienten einen unterschiedlichen Einfluss auf zelluläre mirnas haben können. CTL Fluchtmutationen spielen ebenso eine Rolle, da die vorgelegten Ergebnisse zeigen, dass sie die Bindungsaffinität von zellulären mirnas beeinflussen können. Damit konnte in der vorgelegten Arbeit die Elektroporation von HIV-1-spezifischen mrnas in PBMC als neue effiziente Methode zum Immunmonitoring etabliert werden. Die Verwendung autologer viraler mrna erlaubt zusätzlich die Bestimmung der T-Zellantwort gegen das eigene Virus. Darüber hinaus eignet sich die etablierte Technik, um virale mrnas als Vakzine zur Therapie und Prävention der HIV-1-Infektion zu entwickeln. 2

11 1 Zusammenfassung / Summary Summary A major focus of the thesis was the production and characterization of viral mrna constructs for immunomonitoring of T-cells and for the development of RNA-based vaccines. I could demonstrate that the electroporation of PBMC with autologous HIV-1- specific mrna allows a rapid and efficient analysis of the T-cell responses against the autologous virus. Several HIV-1-specific mrnas encoding Gag, Nef and Env were generated and successfully used for electroporation of PBMC in this thesis. Thereby, a rapid and efficient expression of the mrna-encoded proteins in all subpopulations of PBMC was demonstrated whereat the protein expression was detected up to six days after electroporation. The mrna-encoded HIV-1 proteins were highly immunogenic in vitro and triggered the secretion of γ-ifn and TNF-α. It was shown that the release of γ-ifn was induced by HIV-1-specific T-cells. In addition to the immunogenicity the functionality of the mrna-encoded HIV-1 proteins was also demonstrated. The expression of the molecules CD4, CXCR4 and HLA-I on the cell surface was clearly reduced by mrna-encoded Nef proteins. Moreover, the mrna-encoded HIV-1 proteins could activate CD8 + T-cells in vitro. The mrna-encoded HIV-1 proteins can be used as efficient antigens for the monitoring of T-cell responses in HIV-1-infected patients. They induced stronger CTL responses compared to recombinant vaccinia viruses and similar immune reactions as peptide pools. Using autologous HIV-1-specific mrnas the immune response against autologous viral proteins was compared to the immune response against heterologous viral proteins. In five of ten cases, the autologous viral proteins induced stronger immune responses than proteins of HIV-1 reference viruses, demonstrating that conventional monitoring using heterologous antigens frequently underestimates the magnitude of the CTL responses against the autologous virus. In two cases, the T-cell response against the heterologous viral antigen exceeded the response against the autologous virus which could be explained by CTL escape mutations in the autologous viral sequence. This observation underlines the important role of CTL escape and it also demonstrates that the use of heterologous antigens can also overestimate the HIV-1-specific immune responses. Besides the establishment of a novel efficient tool for monitoring the T-cell responses against autologous viruses, mrna-electroporation can also address important issues concerning the use of HIV-1-specific mrnas as therapeutic vaccines. It could be confirmed that the mrna-encoded proteins Gag and Nef induced better immune responses than Env and that they are suitable antigens for vaccination. However, it was shown that combined application of gag and nef mrnas causes a dramatic loss of the 3

12 1 Zusammenfassung / Summary induced immune responses in PBMC due to the HLA-I reduction by the Nef protein. Moreover, the experiments provided interesting insights about the hierarchy of CTL epitopes and the efficiency of epitope presentation which are useful for the design of vaccines. There are several sequence homologies between HIV-1 and the human genome that could potentially lead to autoimmune reactions after vaccination. Therefore, I investigated the cross-recognition of the human proteins DAO and Fat3 by Nef-specific CTL. I observed cross-recognition of DAO and Fat3-derived peptides by Nef-specific CTL, but I could not detect CTL recognition of cell lines expressing the Fat3 protein. This is suggesting that the cross-reactive peptides are not processed from these proteins. During my thesis I could define three new CTL epitopes in Gag and two in Nef. As HIV-1 Nef downregulates HLA-A and B molecules from the cell surface, there is a growing interest in HLA-C-restricted CTL responses. Therefore, I investigated the recognition of a HLA-Cw*07-restricted CTL epitope in Nef in a cohort of HIV-1-infected patients. I could demonstrate that this epitope is targeted by approximately half of the HLA-Cw*07-positive patients. As the mrna-electroporation is not suitable for in vivo application, I investigated alternative transfection methods. The mrna could be transferred into cells by FuGENE, but not by chitosan, atelocollagen or the HIV-1 protein Tat. To assess the influence of viral proteins on the expression of cellular mirnas, HIV-1 nef and gag mrnas were transfected into purified CD4 + T-cells and the mirna profile was analyzed by sequencing. It could be shown that viral genes from various HIV-1-patients can differently influence cellular mirnas. CTL escape mutations play also a role not only for CTL recognition, but they can also influence the binding affinity of cellular mirnas. In my thesis, the electroporation of HIV-1-specific mrnas in PBMC could be established as a novel, efficient tool for immunomonitoring. The use of autologous viral mrna allows also the determination of the T-cell response against the autologous virus and the analysis of function of the viral proteins. In addition, the results provide useful information for the development of efficient mrna-based vaccines for therapy and prevention of HIV-1-infection. 4

13 2 Einleitung Im Jahre 1983 gelang es Mitarbeitern um Luc Montagnier am Pasteur-Institut in Paris erstmals das Virus aus einem Patienten zu isolieren, welches für das erworbene Immunschwächesyndrom (acquired immunodeficiency syndrome, AIDS) verantwortlich ist (1). Robert Gallo und seiner Gruppe am National Cancer Institute in Bethesda war dies kurze Zeit später möglich. Seit 1986 wird dieses Pathogen humanes Immundefizienz Virus (HIV) genannt und ersetzt die zuvor verwendeten Bezeichnungen wie Lymphadenopathie-assoziiertes Retrovirus (LAV) oder humanes T-Zell Leukämie Virus Typ III (HTLV-III) (2). Luc Montagnier und seine Mitarbeiterin Françoise Barré-Sinoussi wurden 2008 für diese Arbeit mit dem Nobelpreis für Medizin oder Physiologie ausgezeichnet. Seitdem bemühen sich Wissenschaftler darum, die Wirkungsweise des Virus aufzuklären und eine erfolgreiche Therapie zur Bekämpfung des Virus zu entwickeln. Dabei wurde ein komplexes Zusammenspiel zwischen Virus und Wirt entschlüsselt. Das menschliche Immunsystem besitzt viele Mechanismen zur Erkennung und Eindämmung von Viren. Neben CD4 + T-Helferzellen, neutralisierenden Antikörper und zytotoxischen T-Lymphozyten, welche HIV-1-infizierte Zellen erkennen und eliminieren, spielen auch genetische Faktoren des Wirts, wie HLA-Allele und antivirale Faktoren eine wichtige Rolle. Letztendlich gelingt es unserem Immunsystem aber nicht, HIV-1 vollständig zu eliminieren, weshalb eine wirkungsvolle Therapie entwickelt wurde. Obwohl HIV-1 eines der beststudierten Viren ist und dadurch auch eine erfolgreiche Behandlung hervorgebracht wurde, ist die Heilung einer HIV-1-Infektion immer noch nicht möglich. 2.1 Humanes Immundefizienz Virus (HIV) Epidemiologie und Tropismus Weltweit zählt HIV mit ungefähr 34 Millionen infizierten Menschen (Stand 2010) immer noch zu den häufigsten Infektionskrankheiten. In Zentraleuropa lag die Anzahl der Erkrankten bei ca ( ), in Deutschland bei ca ( ) (3). Seit 2006 kann ein Rückgang der Neuinfektionen konstatiert werden (2011: ca ), während vor 2006 ein Anstieg zu verzeichnen war (3, 4). Die Übertragung des Virus erfolgt hauptsächlich sexuell. Die parenterale Übertragung durch kontaminierte Blutprodukte oder Nadeln bei Drogenmissbrauch sowie die vertikale Übertragung während der Schwangerschaft kommen in Deutschland kaum mehr vor. 5
 (2). Luc Montagnier und seine Mitarbeiterin Françoise Barré-Sinoussi wurden 2008 für diese Arbeit mit dem Nobelpreis für Medizin oder Physiologie ausgezeichnet.")
14 2 Einleitung Man unterscheidet zwischen HIV-1 und HIV-2, die sich in ihrer Sequenz bis zu 50% unterscheiden. HIV-2 zeigt ein AIDS-ähnliches Krankheitsbild, die Virulenz ist jedoch deutlich geringer. HIV-2 kommt vor allem in Westafrika und in Indien vor, wohingegen HIV-1 weltweit vertreten ist und sich aufgrund von Sequenzunterschieden in die drei Subtypen M, O und N spaltet (5-7). Die meisten vorkommenden Viren gehören dem M- Subtyp (major) an, der sich weiter in die Genotypen A-I unterteilt. Der Genotyp B hat dabei die größte Bedeutung in den westlichen Industrienationen (8). Subtyp O bezeichnet Ausreißer (outlier) und der N-Subtyp umfasst neue Viren (new). Des Weiteren wird HIV-1 in R5- und X4-Stämmen differenziert. Erstere verwenden den Chemokinrezeptor CCR5 als Ko-Rezeptor und infizieren darüber primär Makrophagen und Lymphozyten. Sie kommen insbesondere in den frühen Stadien der Erkrankung vor und bilden keine Synzytien. Daher werden sie auch als NSI (non-syncytium-inducing)- bzw. M (Makrophagen)-tropische Stämme bezeichnet. Im Gegensatz dazu kommen X4- Stämme, welche den CXC-Motiv-Chemokinrezeptor CXCR4 als Ko-Rezeptor binden, vor allem in den späteren Krankheitsstadien vor und infizieren transformierte CD4 + T-Zelllinien und primäre Lymphozyten. Da sie Synzytien bilden können, werden sie auch als SI (syncytium-inducing)- oder T (T-Zellen)-tropische Stämme bezeichnet. Überdies finden sich auch dual-trope Stämme, die die Eigenschaften beider Stämme vereinen. HIV-1 ist zudem in der Lage durch Mutationen den Tropismus während eines Krankheitsverlaufs zu wechseln (6, 7, 9) Pathogenese und Klinik der HIV-1-Infektion Die akute Infektion mit HIV-1 erfolgt oft unbemerkt, da Symptome bei nur ca % der Infizierten auftreten und sehr unspezifisch wie auch variabel sind. Meist ähneln die Symptome einem grippalen Infekt und machen sich in Form von Fieber, Lymphknotenschwellung oder Erkältungsbeschwerden bemerkbar. In dieser Phase weisen die Patienten eine hohe Virämie mit 10 6 bis 10 8 Viruspartikel/ml Blut auf. Kurzfristig sinkt in dieser Phase die Zahl der CD4 + T-Zellen unter 500 Zellen/µl. Nach dieser kurzen, initialen Virämie, welche meist nur wenige Wochen andauert, sinkt die Viruslast und die Zahl der CD4 + T-Zellen normalisiert sich. Es folgt eine symptomfreie Latenzzeit, die mehrere Jahre anhält. Die Anzahl der CD4 + T-Zellen reduziert sich währenddessen stetig (Abbildung 2.1). CD4 + T-Lymphozyten, Makrophagen und dendritsche Zellen (dendritic cells; DCs) werden durch das Virus immer mehr zerstört, so dass es nach und nach zu einem Ausfall der immunologischen Funktionen dieser Zellen 6
 und der N-Subtyp umfasst neue Viren (new). Des Weiteren wird HIV-1 in R5- und X4-Stämmen differenziert.")

15 2 Einleitung kommt. Fällt die Zahl der CD4 + T-Zellen unter den Schwellenwert von 200 Zellen/µl Blut, kann das Immunsystem freigesetzte Viren nicht mehr abfangen und eliminieren. Abbildung 2.1 Typische Verlaufskurve einer HIV-1-Infektion. Die Infektion gliedert sich in die drei Stadien der Primärinfektion, der klinischen Latenz und dem Endstadium AIDS. Die rote Linie beschreibt den Verlauf der Viruslast im Plasma über die Zeit, die blaue Linie zeigt die Anzahl der CD4 + T-Zellen. Detaillierte Erklärung siehe Text. Abbildung von Wikipedia: (10). Es folgt das Endstadium AIDS. Dieses ist charakterisiert durch wiederholte Infekte mit opportunistischen Erregern und das Auftreten von malignen Tumoren, vor allem von Lymphomen oder von Karposi-Sarkomen. Oft werden auch schwere neurologische Symptome wie Demenz diagnostiziert (7, 11). Nach der CDC-Klassifikation (U.S. Centers for Disease Control and Prevention) lässt sich die HIV-1-Infektion in drei klinische Kategorien einteilen. Kategorie A umfasst das asymptomatische Stadium der Infektion mit >500 CD4 + T-Zellen/µl. Kategorie B beschreibt die symptomatischen Erkrankten ohne AIDS und CD4 + T-Zellen/µl und zur letzten Kategorie C gehören Patienten mit Vollbild AIDS und weniger als 200 CD4 + T-Zellen/µl (12) Aufbau und Replikationszyklus HIV-1 ist der Familie der Retroviren zuzuordnen und gehört der Gattung der Lentiviren an. Sein Genom ist etwa Basen groß und liegt im Viruskapsid in zwei Kopien als lineares, einzelsträngiges, plus-orientiertes RNA-Molekül vor. Die virale RNA besitzt eine 7
. Es folgt das Endstadium AIDS.")
16 2 Einleitung cap-struktur am 5`-Ende und eine Poly-Adenylierung am 3`-Ende, so dass sie mit der mrna (messenger RNA) von Eukaryonten vergleichbar ist. Neben den Genen gag, pol und env, welche in allen replikationskompetenten Retroviren vorkommen, besitzt HIV-1 sechs zusätzliche, sogenannte akzessorische Gene (vif, vpu, vpr, tat, rev und nef). Alle neun Gene sind von langen terminalen Sequenzwiederholungen (long terminal repeats, LTR) umgegeben, welche zur Integration des Virus in das Wirtsgenom benötigt werden (Abbildung 2.2) (13). Abbildung 2.2 Schematischer Aufbau des HIV-1-Genoms. Die Gene gag (orange), pol (rot) und env (blau), kodieren für die Strukturproteine, die Enzyme und die Hüllproteine des Virus. Zusätzlich besitzt HIV-1 noch sechs akzessorische Gene (grün). Erklärung siehe Text. Adaptiert von Costin, 2007 (13). Das Gen gag (group antigen) trägt die Information für die Proteine p17, p24 und p7, welche die Matrix, das Kapsid und das Nukleokapsid bilden, sowie das Linkerprotein p6. Die Glykoproteine gp120 und gp41 für die Virushülle werden von env (envelope) kodiert. Die vom Virus benötigten Enzyme zur Virusduplikation und -integration wie Protease, Reverse Transkriptase und Integrase werden von pol (polymerase) kodiert. Die akzessorischen Gene vif, vpu, vpr, tat, rev und nef enthalten die Information für Proteine, welche die Replikation wie auch die Infektion des Virus entscheidend beeinflussen können. Das Vif-Protein beispielsweise inaktiviert den zellulären, antiviralen Restriktionsfaktor APOBEC3G. Das Nef-Protein hat unter den akzessorischen Proteinen eine zentrale Rolle, da es mit vielen verschiedenen Wirtsproteinen interagieren kann und zudem entscheidend zur Pathogenität des Virus beiträgt. So wurde gezeigt, dass 8
, pol (rot) und env (blau), kodieren für die Strukturproteine, die Enzyme und die Hüllproteine des Virus.")
17 2 Einleitung Rhesusaffen, die mit nef deletierten Viren infiziert sind, einen stark abgeschwächten Krankheitsverlauf aufweisen. Zudem zeigen Sequenzanalysen von Langzeit- Nichtprogressoren (longterm non-progressors, LTNP) höchst mutierte Nef Sequenzen, die unter anderem große Deletionen im nef Gen aufweisen (7, 11, 14-16). Abbildung 2.3 Schematischer Überblick über den Replikationszyklus von HIV-1. Erklärung siehe Text. Abbildung von Wikipdia: (17) Der Lebenszyklus von HIV-1 ist durch mehrere Schritte gekennzeichnet (Abbildung 2.3). Der erste Schritt beschreibt den Eintritt des Virus in die Zielzelle, was über Fusion der viralen Membran mit der Membran der Zielzelle erfolgt. Primär findet diese Verschmelzung bei HIV-1 über Rezeptoren statt, wobei die Oberflächendomäne gp120 des Env-Proteins eine zentrale Rolle spielt. Zunächst bindet gp120 an das CD4-Molekül auf der Oberfläche der Zielzellen, zu welchen CD4 + T-Helferzellen, Makrophagen und Monozyten gehören. Diese Bindung verursacht eine Konformationsänderung in gp120, wodurch die Bindestelle für den Ko-Rezeptor frei wird. Je nach Tropismus des Virus erfolgt die Bindung mit den zellulären Ko-Rezeptor CCR5 oder CXCR4. Dies führt zu einer erneuten Konformationsänderung, welche eine 6-Helices-Struktur ausbildet, die die Fusion der zwei Membranen ermöglicht. Nach Eintritt des Nukleokapsids in das Zytoplasma wird das Nukleokapsid entpackt und die virale RNA wird mit Hilfe der Reversen Transkriptase in eine provirale DNA umgewandelt. Diese wandert in Form eines Präintegrationskomplexes bestehend aus zellulären sowie viralen Faktoren in den Zellkern und nach Aktivierung der Zelle wird die provirale DNA durch die Integrase in die 9
 Der Lebenszyklus von HIV-1 ist durch mehrere Schritte gekennzeichnet (Abbildung 2.3).")
18 2 Einleitung Wirts-DNA eingefügt. Die Integration der proviralen DNA ist Voraussetzung für die Transkription der viralen Gene. Zelluläre Transkriptionsfaktoren wie NF-κB (nuclear factor 'kappa-light-chain-enhancer' of activated B-cells) können die Expression der viralen Gene initiieren, indem sie an den LTR-Bereich der integrierten, proviralen DNA binden. In diesem Bereich befinden sich der Promotor sowie Bindestellen für Transkriptionsfaktoren. Da zunächst regulatorische Proteine wie Tat und Rev exprimiert werden, kann die Expression der viralen Gene in eine frühe und späte Phase unterteilt werden. Rev und Tat stimulieren dabei die weitere Transkription als auch die Ausbildung von langen RNA- Transkripten. Rev fördert zudem die Expression von strukturellen und enzymatischen Genen, welche zur Bildung von viralen Partikeln benötigt werden. Nach Transkription der viralen Gene im Zellkern findet die Translation an den Ribosomen im Zytoplasma, sowie Spaltung von Vorläuferproteinen durch die Protease oder Glykosilierung der Proteine am endoplasmatischen Retikulum statt. Die Produkte von pol und gag sowie die virale RNA lagern sich zusammen und bilden das Nukleokapsid. Die Produkte von env bilden später die spikes auf der Oberfläche der Viren. Im Anschluss werden die Partikel über den Golgi-Apparat zur Zelloberfläche transportiert, können sich dort von der Zelle abschnüren und als neue, infektiöse Viren weitere Zellen infizieren (7, 11, 18). 2.2 Immunologische und genetische Faktoren, die die HIV-1-Infektion und Progression beeinflussen Zytotoxische T-Lymphozyten (CTL) Neben B-Zellen gehören zu den Hauptzellen des adaptiven Immunsystems die T-Zellen, welche nach Entstehung im Knochenmark im Thymus heranreifen. Sie gelangen von dort aus in die Blutbahn als reife, naive T-Zellen, welche noch nicht aktiviert sind. Je nach Oberflächenrezeptor unterscheidet man zwischen zwei Subpopulationen, den CD4 + T-Zellen, auch als T-Helferzellen bezeichnet, und den CD8 + T-Zellen, die auch T-Killerzellen oder zytotoxische T-Lymphozyten (cytotoxic T lymphocytes; CTL) genannt werden. Die T-Helferzellen spalten sich noch in weitere Populationen und können die Aktivierung und Proliferation von CTLs wesentlich beeinflussen (19). Die zytotoxische T-Zelle differenziert erst nach Kontakt mit ihrem Antigen zu einer aktiven Effektorzelle. Der Kontakt findet dabei über den spezifischen T-Zell-Rezeptor (T cell receptor; TCR) der CTL und das antigenbeladene HLA Klasse I-Molekül statt, welches antigenpräsentierende Zellen (antigen presenting cells; APC) wie dendritsche Zellen, Makrophagen und B-Zellen 10

19 2 Einleitung den CD8 + T-Zellen präsentieren. Die aktivierte CTL eliminiert nun Virus-infizierte Zellen, wobei mehrere Mechanismen möglich sind. Zum einen können CTLs Zytokine sezernieren, die antivirale Eigenschaften aufweisen, wie γ-interferon (γ-ifn) oder Tumor- Nekrosis-Faktor-α (TNF-α). Zum anderen können CTLs über ihren Fas-Liganden an den Fas-Rezeptor der Zielzelle binden und so die Apoptose der Zielzelle induzieren (20). Oft nutzen sie auch den TRAIL-Liganden und binden den dazugehörigen Rezeptor auf der Oberfläche der Zielzelle, den death receptor 4 oder 5, was ebenfalls zur Apoptose der Zielzelle führt (21). In den meisten Fällen erfolgt die zytolytische Aktivität von CTLs über die Sekretion von lytischen Granula, die Perforin enthalten und damit die Zielzelle lysieren können (20, 22). HIV-1-spezifische CTLs spielen eine wichtige Rolle in der Immunantwort gegen HIV-1. Bereits im Tiermodell mit Rhesusmakaken, die mit dem simianen Immundefizienz Virus (SIV) infiziert sind, konnte gezeigt werden, dass während der Primärinfektion die virale Replikation ansteigt, wenn CD8 + T-Zellen depletiert werden (23). Gleichermaßen ist die CTL-Aktivität im Menschen invers korreliert mit der Virämie und dem Krankheitsverlauf (24). Die antigenspezifische Zelllyse, welche von den CTLs ausgelöst wird, ist sehr effizient und zudem schnell genug, um HIV-1-infizierte Lymphozyten abzutöten bevor sie eine maximale Virusproduktion erreichen. Eine erfolgreiche Suppression der viralen Replikation findet man insbesondere bei Gag-spezifischen CTLs, CTLs gegen andere HIV-1-Proteine sind oft weniger effektiv (25). HIV-1-spezifische CTLs treten sehr früh im Verlauf der Infektion auf, meist wenn der initiale Peak der Virämie sinkt. Die erste CD8 + T-Zell-Antwort ist jedoch nur gegen wenige Epitope gerichtet (26). In der chronischen Phase der Infektion nehmen die Bandbreite und Stärke dieser Antwort zu, dennoch kann durch CTLs die virale Replikation nur in wenigen Patienten, wie LTNP, kontrolliert werden. Geht es in das Stadium AIDS über, sinken die CTL-Antworten ab (27). Neben der Frequenz der CTL-Antwort ändert sich auch die CTL-Spezifität im Laufe einer Infektion. Einige CTLs finden sich nur in der akuten Phase der Infektion, während andere CTLs einzig in der chronischen Phase auftauchen (28). Durch Mutationen in der viralen Sequenz können CTL-Epitope entweder verloren gehen, nicht mehr prozessiert werden oder nicht mehr an das HLA-Molekül binden. Auch eine fehlende Erkennung durch den TCR kann Folge einer Mutation sein (27). Diese sogenannten CTL Fluchtmutationen treten sehr häufig auf. Da das Immunsystem nachreift und neue TCR mit anderer Spezifität bildet, kommt es anschließend zu einer zweiten und eventuell auch dritten CTL- Antwort. Aufgrund der nachgereiften CTLs können nun verschiedene Virusvarianten, auch Fluchtmutanten, kreuzreaktiv erkannt werden. Patienten, die ein größeres Repertoire an kreuzreaktiven CTLs besitzen, kontrollieren das Virus in der akuten Phase besser. Daher könnte die Effektivität einer CTL-Antwort nicht nur durch die antivirale zytotoxische 11
.")
20 2 Einleitung Aktivität beeinflusst werden, sondern eben auch durch ihre kreuzreaktiven Eigenschaften (28). Des Weiteren wurde festgestellt, dass Patienten, die HIV-1 gut kontrollieren, CD8 + T-Zellen mit einer höheren Polyfunktionalität besitzen. Diese können mehrere Zytokine wie γ-ifn, TNF-α, Interleukin-2 (IL-2) sowie MIP-1β sekretieren (29, 30). Demzufolge spielen HIV-1-spezifische CTLs eine entscheidende Rolle zur Eliminierung des Virus und insbesondere LTNP weisen meist eine hohe CTL-Aktivität auf HLA-Allele Eine Ansammlung von Genen, die auf dem humanen Chromosom 6 liegen, werden als Haupthistokompatibilitätskomplex (major histocompatibility complex, MHC) oder als humanes Leukozytenantigen-System (humane leukocyte antigen, HLA) bezeichnet. Sie kodieren für Proteinkomplexe, die eine entscheidende Rolle in der Immunantwort und der individuellen, zellulären Immunität spielen. So sind diese Proteinkomplexe unter anderem an der Differenzierung zwischen fremden und eigenen Proteinkomponenten beteiligt. Es werden zwei Klassen von Molekülen unterschieden. HLA Klasse I-Moleküle (HLA-I) präsentieren endogene, acht bis neun Aminosäuren lange Peptide den CD8 + T-Zellen, welche das peptidbeladene HLA-Molekül über einen spezifischen TCR erkennen. Dabei werden zunächst Proteine im Zytoplasma, die entweder fehlgefaltet sind oder von einem Virus oder einem Tumor stammen, von der Protease in Peptide gespalten. Anschließend werden im endoplasmatischen Retikulum neusynthetisierte HLA-Moleküle mit passenden Peptiden beladen und über den Golgi-Apparat an die Zelloberfläche transportiert (siehe Abbildung 2.4) (27, 31, 32). Dagegen präsentieren HLA Klasse II-Moleküle (HLA-II) exogene, Aminosäuren lange Peptide den CD4 + T-Zellen. Die Aufnahme der Peptide erfolgt über Endozytose. Jeder Mensch besitzt jeweils zwei der HLA-I-Moleküle der drei Hauptgruppen A, B und C. Von den HLA-II-Molekülen werden ebenfalls jeweils zwei Moleküle der Gruppe DQ, DR und DP exprimiert. Die verschiedenen Gruppen differenzieren sich in ihren Peptidbindungseigenschaften. Die Nomenklatur für HLA-Allele setzt sich stets aus der Abkürzung HLA, dem Genort und einer Zahl zusammen. So bezeichnet bei HLA-B*1301 der Buchstabe B den Genort, die Zahl 13 ist die Nummer der Gruppe für das spezifische Antigen und 01 die Subgruppe (33). Assoziationsstudien haben gezeigt, dass es einen Zusammenhang zwischen dem individuellen HLA-Typ und der unterschiedlich starken Virämie bei HIV-1-Patienten gibt (34). Insbesondere konnte dieser Zusammenhang für das HLA-B-Allel beobachtet werden. Die HLA-Allele B*57, B*58, B*62, B*27 und B*1302 sind alle mit einer guten Kontrolle von HIV-1 assoziiert (35). Dabei zeigen HLA-B*5701 und B*5703 den 12

21 2 Einleitung stärksten, protektiven Effekt unter Kaukasiern und Afrikanern (36). Diese HLA-Allele binden bestimmte virale Epitope, die Fluchtmutationen nicht tolerieren können, da sonst die Fitness und Replikationsfähigkeit des Virus erheblicher eingeschränkt wird (37). Dagegen gibt es auch HLA-I-Allele, die mit einem schlechteren Krankheitsverlauf sowie einer deutlich schnelleren Progression der Erkrankung assoziiert sind, wie z.b. einige Subtypen von HLA-B*35 oder der Haplotyp A*01-B*08-DR*03 (38-41). Des Weiteren weisen Patienten mit homozygoten HLA-Allelen einen deutlich schlechteren Krankheitsverlauf auf als Patienten mit heterozygoten HLA-Allelen. Dies liegt daran, dass Patienten mit heterozygoten HLA-Allelen ein größeres Repertoire an viralen Epitopen präsentieren können (42, 43). Die Immunantwort hängt folglich sehr stark vom individuellen HLA-Typ ab. Man schätzt, dass allein der HLA-I-Typ der Patienten ungefähr für 6-15% der Variationen in den Viruslasten verantwortlich ist (37). Abbildung 2.4 Schematische Darstellung des HLA Klasse I Signalweges und die Erkennung durch CTL. In einer Virus-infizierten Zelle wird die integrierte provirale DNA zu mrna transkripiert, wandert aus dem Zellkern in das Zytosol und wird dort zu einem viralen Protein translatiert. Dieses wird unter anderem durch das Proteasom zu kleinen Peptidfragmenten gespalten, welche mit Hilfe von TAP (transporter associated with antigen presenting) in das Endoplasmatische Retikulum gelangen. Einige der Peptide können nun an MHC-I- Moleküle binden, woraufhin sie an die Zelloberfläche transportiert werden. Der TCR einer CTL kann spezifisch bestimmte MHC-I-Moleküle mit ihrem gebunden Peptid erkennen und dadurch eine Immunreaktion auslösen. Der funktionsfähige TCR ist ein Heterodimer aus einer α- und einer β-kette, die beide jeweils aus einer konstanten (C) und einer variablen (V) Domäne bestehen. Das MHC-I Molekül besteht aus einer α-kette aus drei Domänen und verbindet sich mit dem extrazellulären β2-mikroglobulin. Adaptiert von Sewell, 2000 (27). 13
22 2 Einleitung Toll-like Rezeptoren in der angeborenen Immunantwort Die erste Immunantwort gegen eine HIV-1-Infektion ist die angeborene Immunantwort. Dazu gehören unter anderem auch Toll-like Rezeptoren (TLR), die von Viren und Bakterien bestimmte Motive erkennen und eine direkte Immunaktivierung auslösen. Die einzelsträngige RNA von HIV-1 wird von TLR 7 und 8 erkannt, was zur Produktion von antiviralen Zytokinen führt (44). In erster Linie wird Interferon-α (IFN-α) freigesetzt, welches die Expression von intrinsischen Restriktionsfaktoren erhöht und andere Immunzellen wie B- und T-Zellen reguliert (44, 45). Allerdings spielen auch genetische Faktoren des Wirts eine Rolle. So wurde eine erhöhte Produktion von IFN-α bei Frauen gefunden, was wahrscheinlich auf eine erhöhte Expression von TLR 7 zurückzuführen ist, da sich der Genlokus für TLR 7 auf dem X-Chromosom befindet (46). Darüber hinaus kommen in dem Gen für TLR 7 Einzelpolymorphismen (single nucleotide polymorphism, SNP) vor, welche zu einer geringeren Freisetzung von IFN-α führen. Dies resultiert letztendlich auch in einem schnelleren Krankheitsverlauf bei HIV-1 (47) CD4 + T-Helferzellen Nachdem lange Zeit die Rolle der CD4 + T-Zellen für den Krankheitsverlauf bei HIV-1 umstritten war, weiß man nun, dass nicht nur CD8 + T-Zellen einen Einfluss auf den Verlauf einer HIV-1-Infektion haben, sondern auch die CD4 + T-Helferzellen sehr wichtig sind. Eine starke HIV-1-spezifische CD4 + T-Zellantwort ist mit einer besseren Kontrolle der Virusreplikation assoziiert. HIV-2, welches eine geringere Pathogenität im Gegensatz zu HIV-1 besitzt, geht stets mit robusten und polyfunktionalen CD4 + T-Zellen einher (48, 49). Zum einen sind CD4 + T-Zellen von Bedeutung, da sie IL-2 produzieren, welches das wichtigste Proliferationssignal für CTLs ist. Zum anderen erkennen CD4 + T-Zellen Epitope, welche von HLA Klasse II präsentiert werden. Dass die daraufhin von den CD4 + T-Zellen ausgelöste Immunantwort eine wichtige Rolle in der Kontrolle der Virusreplikation spielt, zeigen Fluchtmutationen in den HLA Klasse II Epitopen. CD4 + T-Zellen verschwinden zwar nach der initialen Virämie, dennoch üben sie genug Druck aus, dass das Virus Mutationen in diese Epitope einbaut, um den Druck des Immunsystems zu entkommen (50). Außerdem konnten nachgewiesen werden, dass HIV-1-Patienten, die das Virus sehr gut kontrollieren, eine Population von Gedächtnis CD4 + T-Zellen besitzen, die eine hohe Avidität gegen das Gag-Protein aufweisen (51). Obwohl CD4 + T-Zellen bevorzugt von HIV-1 infiziert werden, bleiben die meisten CD4 + T-Zellen auch bei einer hohen Virämie virusfrei (52). 14
23 2 Einleitung Genetische Unterschiede im Gen des Chemokinrezeptor CCR5 Neben den bereits genannten Aspekten tragen noch weitere genetische Faktoren des Wirts zum Verlauf einer HIV-1-Infektion bei. So wurde für den Chemokinrezeptor CCR5 eine natürlich auftretende Deletion von 32 Basen beschrieben, die bei ca. 1% der Kaukasier zu finden ist. Diese CCR5Δ32 Mutation resultiert in einem verfrühten Stoppcodon und somit in einem nicht-funktionsfähigen Rezeptor. Da CCR5 der Ko- Rezeptor für den Eintritt der Viren in Zellen ist, können R5- bzw. M-trope Viren Zellen nicht mehr infizieren, X4- bzw. T-trope und dual-trope HIV-Stämme indessen schon (53, 54). Homozygote Merkmalsträger für CCR5Δ32 sind damit resistent gegenüber einer HIV-1-Infektion von R5-tropen Viren. Heterozygote Merkmalsträger zeigen einen langsameren Krankheitsverlauf mit einer Verzögerung in das Stadium AIDS von 2-4 Jahren (55, 56). Wie wichtig dieser genetischer Faktor für die Entwicklung einer Gentherapie bei HIV-1 ist, zeigt der Fall des sog. Berlin-Patienten. Dieser konnte mit einer Stammzell-Transplantation eines Spenders, der Merkmalsträger für CCR5Δ32 ist, von HIV-1 geheilt werden. Die Stammzell-Transplantation bei diesem HIV-1-infizierten Patienten aus Berlin wurde durchgeführt, da dieser an Leukämie erkrankt war und die Chemotherapie keinen Erfolg zeigte (57). Seit der Operation im Februar 2007 konnten bei ihm keine Viren mehr nachgewiesen werden (58). Der Erfolg dieser Therapie wird wahrscheinlich ein Einzelfall bleiben. Dennoch konnte aus diesem Fall viel gelernt werden und es öffneten sich daraus neue Perspektiven einer Gentherapie für die Behandlung von HIV Interaktion von micrornas und Viren MicroRNAs (mirna) sind einzelsträngige, Nukleotid lange nicht-kodierende RNAs, welche die Genexpression post-transkriptionell regulieren. Zur Zeit sind beim Menschen verschiedene mirnas bekannt, die neben der Regulierung von zellulären Prozessen wie der Entwicklung des Immunsystems (59, 60), der Signaltransduktion (61) oder der Zellproliferation auch eine Rolle in der Tumorgenese (62), der Entstehung inflammatorischer Erkrankungen (63) sowie bei der Replikation von Viren spielen (64). Die Regulation von zellulären mrnas erfolgt dabei über die Bindung von mirnas an teilweise komplementäre Basensequenzen meist in der 3 -untranslatierten Region (UTR) oder seltener im offenen Leseraster, was schließlich den Abbau der mrna oder eine Inhibierung der Translation zur Folge hat (65). 15
24 2 Einleitung Abbildung 2.5 Die Biogenese von mirnas. MicroRNAs sind entweder in eigenen Genen kodiert oder aber werden aus Introns gebildet. Dabei werden zunächst die mirna-gene zu primären mirnas (pri-mirnas) transkripiert. Durch Drosha, eine nukleäre RNase III, werden die pri-mirnas zu precursor mirnas (premirnas) gespalten, welche anschließend über Exportin aus dem Zellkern in das Zytoplasma gelangen. Durch Dicer wird die precursor mirna zu einer reifen, doppelsträngigen mirna prozessiert. Dieser Duplex wird in den RISC Komplex, welcher unter anderem aus Argonaut-Proteinen entsteht, aufgenommen. Darin trennt sich der Duplex auf. Bindet die mirna an eine komplementäre mrna-sequenz mit einer vollständigen Übereinstimmung, so kann die mrna direkt abgebaut werden. Falls die Bindung zwischen mirna und mrna einige Fehlpaarungen enthält, wird die Translation der mrna inhibiert, die mrna jedoch nicht degradiert. Abbildung adaptiert von Wikipedia: (66). Einen ersten Hinweis auf eine Interaktion zwischen HIV-1 und mirnas lieferte die Entdeckung, dass HIV-1 mrna mit Effektoren des mirna-stoffwechselweges in zellulären p-body -Strukturen ko-lokalisiert (67). Mehrere Mechanismen sind für eine mögliche Modulation der Wirt-Virus-Interaktion durch mirnas denkbar (67). Durch virale Infektionen kann das zelluläre mirna-milieu verändert werden (68, 69). Eine Virusinfektion induziert den antiviralen Verteidigungsmechanismus einer Zelle. Dabei werden spezifische mirnas exprimiert, um die Replikation des Virus zu inhibieren. So reduziert z.b. mir-122 spezifisch die Replikation des Hepatitis C-Virus in der Leber (70) und mir-29a inhibiert die Nef-Expression und damit die HIV-1-Replikation in Jurkat T-Zellen durch Bindung der Zielsequenz im nef Gen (71). Eine weitere Möglichkeit wäre, dass Virusinfektionen durch die Induktion von mirnas die Expression von verschiedenen zellulären Genen verringern, wobei das Virus das mirna-muster zu seinen Gunsten verändert. Viren können auch selbst mirnas kodieren, um die Expression von WirtsmRNAs zu inhibieren. Die Hemmung von viralen mrnas durch Virus-kodierte mirnas 16
25 2 Einleitung stellt eine vierte Möglichkeit dar, wodurch verschiedene Schritte des viralen Lebenszyklus reguliert werden können, wie z.b. der Übergang in die Latenzphase (72, 73). Ein Beispiel bildet die von HIV-1 im nef Gen kodierte mir-n367, welche die HIV-1-Transkription vermindert (72). Außerdem wurde bereits gezeigt, dass das regulatorische HIV-1-Protein Tat eine zentrale Rolle bei der HIV-1-Replikation spielt und einige mirnas ein verändertes Expressionsmuster nach Tat-Behandlung aufweisen (74). Da das HI-Virus aufgrund seiner hohen Sequenzdiversität sehr variabel ist, kann es der Inhibierung mittels mirnas durch Mutationen in der Zielsequenz der mirna entkommen (65). 2.3 Therapie und Behandlungsansätze von HIV Hochaktive antiretrovirale Therapie (HAART) Die hochaktive antiretrovirale Therapie (highly active antiretroviral therapy, HAART) wird seit 1996 zur Behandlung einer HIV-1-Infektion eingesetzt. Die verschiedenen Medikamente werden in fünf Gruppen unterteilt: Nukleosidische Reverse-Transkriptase- Inhibitoren (NRTI), Nicht-Nukleosidische Reverse-Transkriptase-Inhibitoren (NNRTI), Protease-Inhibitoren (PI), Integrase-Inhibitoren und Eintritts- bzw. Fusionshemmer (75). Darunter waren NRTI die ersten HIV-1-Medikamente, für welche es bereits 1987 eine Zulassung gab. Sie werden von der Zelle direkt aufgenommen und entfalten ihre Wirkung erst nach intrazellulärer Phosphorylierung (11). Sie bestehen aus alternativen Substraten für die Reverse Transkriptase, so dass diese mit den physiologischen Nukleosiden konkurrieren. Werden diese Nukleosidanaloga in den DNA-Strang während der cdna- Synthese eingebaut, kommt es zum Abbruch der reversen Transkription (76). Die Gruppe der NNRTI hingegen, welche das erste Mal 1990 beschrieben wurde, bindet direkt und nicht-kompetitiv an die Reverse Transkriptase (77). Bei den Protease-Inhibitoren handelt es sich um Peptidanaloga, die an das aktive Zentrum der Protease binden und so die Prozessierung von viralen Proteinen verhindern (78). Seit 2007 sind auch Integrase- Inhibitoren auf dem Markt, welche die virale Integrase hemmen und dadurch den Einbau der viralen cdna in die Wirts-DNA unterbinden (79). Ebenfalls erst seit kurzem gibt es Eintritts- bzw. Fusionshemmer, welche den Eintritt der Viren in die Zelle blockieren. Diese bestehen aus Ko-Rezeptor-Antagonisten (CCR5 bzw. CXCR4) und Fusionsinhibitoren, welche die Verschmelzung zwischen Virus und Zellen verhindern (80). Bei der antiretroviralen Therapie handelt es sich um eine Kombinationstherapie aus bis zu fünf verschiedenen Medikamenten. Mit den Jahren hat sich die Kombination von drei 17
26 2 Einleitung Medikamenten etabliert, wobei meist zwei Reverse-Transkriptase-Inhibitoren und ein Protease-Inhibitor verabreicht werden (81). Die Verwendung von mehreren Medikamenten soll sicherstellen, dass die Wahrscheinlichkeit für die Entwicklung von Resistenzmutationen stark verringert wird (7). Ziel der HAART ist die Unterdrückung der viralen Replikation und die Erhöhung der CD4 + T-Zellen, so dass die Viruslasten im Idealfall unter der Nachweisgrenze von <20 Kopien/ml liegen. Damit kann eine Progression in das Stadium AIDS verhindert werden. Dank der HAART kann heute wahrscheinlich eine weitgehend normale Lebenserwartung trotz HIV-1-Infektion möglich werden, eine Heilung jedoch nicht. Die HAART ist somit sehr effektiv, allerdings können dabei auch starke Nebenwirkungen auftreten. Lipodystrophie, mitochondriale Toxizität, Leberschäden und Herzinfarkte können Folgen der HAART sein (11). Des Weiteren können bei Einnahmefehlern sehr schnell resistente Viren auftreten, die einen Therapiewechsel erfordern (82). Ein weiterer Nachteil der HAART sind die immensen Kosten der Therapie, die sich in Deutschland auf ca pro Jahr und pro Patient belaufen (83). Aufgrund dieser hohen Kosten ist die Therapie gerade in Entwicklungsländern nicht immer, und wenn nur eingeschränkt, verfügbar (84) Vakzine Aufgrund der unter beschriebenen Nachteile der HAART ist die Suche nach wirkungsvollen, therapeutischen Impfstoffen als alternative Therapie immer noch im vollen Gange. Dies zeigen auch die geleisteten finanziellen Investitionen. Im Jahr 2008 wurden beispielsweise weltweit 868 Millionen USD für Forschung und Entwicklung von HIV-1- Vakzinen ausgegeben (85). Über die Jahre wurden unterschiedliche Ansätze verfolgt, jedoch gibt es bis heute noch keinen zugelassenen Impfstoff. Das Hauptproblem in der Entwicklung einer erfolgreichen Vakzine liegt in der antigenen Plastizität des Virus und der Fähigkeit sich dem Immunsystem immer wieder entziehen zu können (86, 87). Bei den meisten viralen Infektionen handelt es sich bei den Impfstoffen um attenuierte, tote oder fragmentierte Pathogene, welche eine Antikörper-Antwort induzieren. Diese Antikörper können die Infektion weiterer Zellen verhindern und zusätzlich die zelluläre Immunantwort aktivieren, um Virus-infizierte Zellen zu eliminieren. Neutralisierende Antikörper erkennen und binden Proteine auf der Oberfläche von Viren. Auf der Suche nach neutralisierenden Antikörpern gegen HIV-1 sind neben der Hypervariabilität des Virus, die Instabilität der Env-Moleküle, welche Ziel der neutralisierenden Antikörper sind, und die Unzulänglichkeit der konservierten Zielbereiche entscheidende Hindernisse (85). Der erste neutralisierende Antikörper gegen HIV-1 mit dem Namen b12 wurde
27 2 Einleitung isoliert und ist gegen die CD4-Bindestelle von gp120 gerichtet (88). Es folgten weitere Antikörper, die jedoch alle nicht zur therapeutischen Anwendung kamen. Nachdem sich die Suche nach neutralisierenden Antikörpern zur Vakzinierung als schwierig erwies, richtete sich der Fokus verstärkt auf HIV-1-Vakzine, welche eine T-Zell- Antwort, insbesondere CTL-Antwort, induzieren können. Jedoch können CTLs nur bereits infizierte Zellen erkennen und eliminieren. Nachdem in HIV-1-exponierten, nicht-infizierten Menschen HIV-1-spezifische CTLs nachgewiesen wurden, keimte Hoffnung auf, dass man durch eine T-Zell-basierte Vakzine erste, kleine Infektionsherde eindämmen könnte (89). Da sich CTLs dem Immunsystem durch CTL Fluchtmutationen (siehe 2.2.1) entziehen können, ist es für eine effiziente Vakzine wichtig, möglichst hoch konservierte CTL-Epitope zu integrieren. Um CTLs zu aktivieren, stehen verschiedene Techniken zur Verfügung. Man verwendet unter anderem DNA, welche HIV-1-Antigene kodiert. Dazu wird Plasmid-DNA intradermal, subkutan oder intramuskulär verabreicht. Nach Transkription und Translation zu Protein werden die entsprechenden Antigene von den APC präsentiert, wodurch T-Zellen aktiviert werden (90). Allerdings scheiterten erste Studien mit DNA-Vakzinen aufgrund ihrer mangelnden Immunogenität. Dieses Problem wurde versucht zu umgehen, indem rekombinante Viren oder Proteine zur Verbesserung der Immunantworten eingesetzt wurden. Eine wichtige Plattform ist dabei das modifizierte Vaccinia-Virus Ankara (MVA), welches mit HIV-1-Antigenen wie z.b. nef verwendet wurde. Dieses war sehr gut verträglich und induzierte zudem eine erhöhte CTL-Antwort (91). Ein weiterer Vektor, der die Immunogenität von DNA-Vakzinen steigert, ist das humane Adenovirus Serotyp 5 (Ad5). Dieser Vektor wurde in einer klinischen Studie Phase II von Merck, der STEP-Studie, verwendet. Dabei wurden drei Dosen des Vektors mit den HIV-1-Genen gag, pol und nef Personen verabreicht. Die Studie wurde allerdings abgebrochen, da es zu einer erhöhten Anzahl von HIV-1-Infektionen in der immunisierten Gruppe kam (87, 92). Eine andere klinische Studie namens RV144 mit Freiwilligen wurde in Thailand durchgeführt und verwendete ein rekombinantes Kanarienpockenvirus, welches als Vektor für die HIV-1-Gene gag, pol und zwei Molekülen env gp120 eingesetzt wurde. Nach dreieinhalb Jahren konstatierte man 31% weniger HIV-1-Infektionen in der immunisierten Gruppe im Vergleich zur Placebo-Gruppe (93). Die RV144-Studie ist bis jetzt die einzige, großangelegte Impfstudie im Menschen, die eine moderate Effizienz zeigte. Vorteile der DNA-Vakzine sind ihre Flexibilität, Stabilität, einfache Lagerung und Sicherheit, da sie im Gegensatz zu normalen Impfstoffen keine lebenden oder abgetöteten Viren enthalten. Diese Impfstoffe wären aufgrund von Sicherheitsgründen auch nicht akzeptabel. Auf der anderen Seite gehen DNA-Vakzine auch mit einigen Nachteilen einher. So kann es zum Einbau der eingebrachten DNA in die zelluläre DNA 19
28 2 Einleitung kommen, wodurch Onkogene aktiviert werden oder Chromosomenbrüche auftreten können. Außerdem kann es zur Entwicklung von Autoimmunitäten und antibiotischen Resistenzen beitragen (90). Alle bisherigen Studien mit Ausnahme der RV144-Studie schlugen fehl, was vermutlich an der immer noch zu geringen Immunogenität der verwendeten Impfstoffe liegt Verwendung von mrna als Vakzine Aufgrund der Nachteile der DNA-Vakzinierung wurde mrna als alternative Nukleinsäure zur Impfung interessant (94). Da es sich um eine Nukleinsäure handelt, die nur transient in den eingebrachten Zellen exprimiert wird, wird die genetische Information nicht dauerhaft in den Organismus integriert. Demzufolge kommt es zu keiner Integration in die Wirts-DNA, wie es beim Einsatz von DNA der Fall ist. Die in vitro generierte mrna ist hoch immunogen und benötigt keine weiteren Vektoren wie Vaccinia-Viren. Sowohl in vitro als auch in Tiermodellen konnte die Induktion von CTLs und Antikörpern durch mrna nachgewiesen werden (95-97). Neben der guten Immunogenität von mrna wurde in vitro eine starke Stimulierung der TLR 3, 7 und 8 durch RNA detektiert. Diese Ko- Stimulierung könnte dabei als eine Art Adjuvans wirken (44, ). Die Wirkungsweise funktioniert wie bei DNA. Die mrna, die ein bestimmtes Protein als Antigen kodiert, wird von der Zelle aufgenommen, geht allerdings nicht in den Zellkern, sondern es findet sogleich die Proteintranslation im Zytoplasma statt. Die Antigene werden dann über die HLA-Moleküle den T-Zellen präsentiert, was eine Aktivierung von CTLs zur Folge hat. Der Einsatz von mrna begann 1995 in der Behandlung von Tumorerkrankungen, was viele klinische Studien nach sich zog (101). Heute findet man die Therapie mit mrna vorwiegend immer noch im Bereich der Tumorforschung, wobei es nun aber auch erste Studien für deren Einsatz bei Infektionskrankheiten gibt (102, 103). Das größte Problem bei der Verwendung von mrna stellt die geringe Halbwertszeit dar, da mrna sehr instabil ist und leicht von RNasen degradiert werden kann (104). Mehrere Methoden wurden zur Stabilisierung der mrna getestet. Eine Verlängerung der Poly-A- Sequenz am 3`-Ende der mrna sowie ein antireverses Cap-Analog (ARCA) tragen zur Stabilitätsverbesserung bei (105, 106). Da RNasen ubiquitär vorhanden sind, muss auch bei Verabreichung der Vakzine die mrna geschützt werden. Dazu gab es Studien mit mrna, welche mit Substanzen wie Protamin oder Liposomen geschützt wurde, und erfolgreich CTLs in Mäusen induzieren konnte (107, 108). Die Anwendung von mrna erfolgt in den meisten klinischen Studien intradermal. Die mrna kann dem Patienten aber auch subkutan, intravenös oder intranodal zugeführt werden. Zusätzlich zu den direkten 20
29 2 Einleitung Injektionsarten von mrna besteht auch die Möglichkeit dendritische Zellen des Patienten zur Therapie zu entnehmen, in vitro mrna in diese Zellen einzubringen und anschließend dem Patienten wieder zu verabreichen (109). Die neuste Entwicklung im Bereich der mrna-therapie sind autologe mrna-vakzine ( ). Diese umgehen das Problem der hohen Sequenzvariabilität der Antigene, da das Antigen direkt aus dem Patienten isoliert und zur Generierung von mrna verwendet wird. Aufgrund der immer noch fehlenden Vakzine zur Therapie einer HIV-1-Infektion, aber den Fortschritten im Bereich der mrna-technologie, rückte die Generierung und Analyse einer autologen, therapeutischen mrna-vakzine in den Fokus dieser Arbeit. Vor allem deren Anwendung in PBMC (peripheral blood mononuclear cells; mononukleäre Zellen des peripheren Blutes) würde eine schnelle und kostengünstige Methode erlauben. Da es bisher keine effiziente Strategie für das Monitoring von CTL-Antworten gegen autologe Virusproteine gibt, sollten autologe mrnas zudem für diesen Einsatz getestet werden. 21
30 3 Zielsetzung Das primäre Ziel der vorgelegten Dissertation war die Generierung von mrnas der HIV-1-Gene gag, nef und env und deren in vitro Testung auf Funktionalität und Immunogenität. Dabei sollten sowohl mrnas aus autologen Virussequenzen von HIV-1- infizierten Patienten sowie mrnas aus HIV-1 Referenzstämmen hergestellt werden. Anschließend sollten diese mrnas im Hinblick auf eine mögliche Anwendung als therapeutischer Impfstoff präklinisch untersucht und zudem als neue Methode zum Monitoring der Immunantworten von HIV-1-Patienten etabliert werden. Zunächst sollte die mrna-elektroporation von PBMC etabliert werden, da die HIV-1- spezifischen mrnas in PBMC exprimiert werden sollten. Um die mrnas auf ihre Funktionalität und Immunogenität zu untersuchen, sollten zum einen die Expression der mrna-kodierten Proteine in den Zellen detektiert und zum anderen die transfizierten Zellen in immunologische Assays eingesetzt werden. Da die hohe Sequenzvariabilität von HIV-1 eines der großen Probleme in der Vakzinentwicklung ist, sollten die Immunantworten, welche durch autologe mrna-kodierte HIV-1-Proteine ausgelöst werden, mit denen verglichen werden, welche durch mrna-kodierte Proteine von Referenzviren induziert werden. Durch diese Analysen sollte geklärt werden, ob eine therapeutische Vakzine optimaler Weise aus autologen oder heterologen Virussequenzen bestehen soll. Des Weiteren sollte das Risiko von Nebenwirkungen bei einer Impfung möglichst gering sein. Daher sollte ausgeschlossen werden, dass HIV-1-spezifische CTLs, die durch mrna-kodierte HIV-1-Proteine aktiviert werden, mit humanen Proteinen kreuzreagieren und dadurch auch nicht-infizierte Zellen zerstören. Aus diesem Grunde sollten HIV-1-spezifische CTL Linien generiert und auf ihre Kreuzreaktivität zu humanen, homologen Epitopen überprüft werden. Außerdem ist die Auslösung einer breit gefächerten Immunreaktion ein weiterer, wichtiger Aspekt bei einer Impfung, so dass eine Vakzinierung mit HIV-1-spezifischen mrnas eine Kombination aus verschiedenen mrnas sein wird. In diesem Zusammenhang sollte ermittelt werden, wie sich die Verabreichung von zwei unterschiedlichen HIV-1-spezifischen mrnas auf deren Effektivität und Immunogenität auswirkt. Das Immunmonitoring der HIV-1-spezifischen T-Zellantwort von HIV-1-Patienten ist sehr wichtig für das Verständnis der Pathophysiologie der HIV-1-Infektion. Bisherige Methoden, welche Peptide, Peptid-Pools oder rekombinante Vaccinia-Vektoren verwenden, detektieren jedoch nur die T-Zellantwort gegen HIV-1 Referenzstämme und nicht gegen das autologe Virus. Deswegen sollten die generierten, autologen HIV-1- spezifischen mrnas auch für den Einsatz im Immunmonitoring getestet und mit den Standard-Techniken verglichen werden. Damit könnte eine umfassende Analyse der 22
31 3 Zielsetzung Quantität der T-Zellantwort gegen das autologe HI-Virus ermöglicht werden. Im Zuge des Immunmonitorings sollten auch neue CTL-Epitope definiert werden. Ein weiterer Ansatzpunkt in der vorgelegten Dissertation sollte die Erprobung von alternativen Transfektionsmethoden sein, um mrna sicher und gut verträglich in Zellen transportieren zu können. Da die Elektroporation zwar eine gute Vorgehensweise für in vitro Studien repräsentiert, jedoch in vivo nicht zugelassen ist, ist die Suche nach alternativen Transfektionsreagenzien für eine in vivo Applikation unumgänglich. Ferner sollte mit Hilfe der generierten HIV-1-spezifischen mrnas der Einfluss einzelner HIV-1-Gene auf das zelluläre mirna-profil von CD4 + T-Zellen studiert werden. Zwar ist bekannt, dass eine HIV-1-Infektion das zelluläre mirna-muster verändert, allerdings wurden in den bisherigen Studien nur komplette HI-Viren eingesetzt. Daher ist keine Aussage möglich, welche HIV-1-Gene welche mirnas speziell beeinflussen. Die Analyse von CD4 + T-Zellen ist dabei besonders interessant, da es sich um die Zielzellen der HIV-1-Infektion handelt. 23
32 4 Ergebnisse 4.1 Elektroporation von PBMC mit HIV-1-spezifischen mrnas Erfolgreiche Generierung von HIV-1-spezifischen mrnas Für die Herstellung von HIV-1-spezifischer mrna wurde zunächst die PCR zur Amplifikation der Gene gag, nef und env etabliert. Nach PCR-Aufreinigung und Sequenzierung wurden die Fragmente in den Vektor pgem4z64a bzw. pgemsigsurvivindclamp kloniert und in vitro zu mrna transkripiert. Auf diese Weise wurden insgesamt sechs autologe gag mrnas, sieben autologe nef mrnas und zwei autologe env gp120 mrnas generiert. Von Patient 0143 existierte bereits zu Beginn meiner Arbeit autologe nef mrna. Von fünf Patienten stehen nun autologe gag und nef mrnas zur Verfügung, von vier Patienten ist nur jeweils eines der beiden Gene als autologe mrna vorhanden. Der Grund hierfür liegt in der Amplifikation der PCR- Produkte. Trotz Verwendung von verschiedenen Primern zur spezifischen cdna- Synthese und PCR konnte kein oder kein ausreichendes Produkt zur Klonierung gewonnen werden. Auch Abwandlungen der PCR-Bedingungen wie Zyklenanzahl oder Annealing-Temperatur und die Veränderung der Salzkonzentration in der PCR-Reaktion führten zu keinem Erfolg. Vermutlich können die verschiedenen Primer nicht binden, da sich in deren Binderegion Mutationen in der Virussequenz des Patienten befinden. Eine zu geringe Ausgangskonzentration der isolierten, viralen RNA könnte ebenfalls eine Rolle spielen. Die Generierung von autologer env gp120 mrna führte aufgrund der hohen Sequenzvariabilität in env nur in zwei Fällen zum Erfolg. Neben autologen HIV-1- spezifischen mrnas wurden zudem von den HIV-1 Referenzstämmen NL4-3 und SF2 mrnas generiert. Dies erfolgte über PCR der Plasmide pnl4-3 und pdsf2, welche HIV-1-Gene enthalten. Eine Übersicht der generierten Produkte ist in Tabelle 4.1 dargestellt. 24
33 4 Ergebnisse Tabelle 4.1 Übersicht über die erfolgreich generierten HIV-1-spezifischen mrnas Virusisolat gag nef env (gp120) NL4-3 SF * *Nicht in dieser Arbeit generiert, sondern von Fr. Dr. K. Maurer Test und Titration von HIV-1-spezifischen Antikörpern in transfizierten HEK293T Zellen Zur Bestimmung der Proteinexpression der mrna-elektroporierten Zellen sollten intrazelluläre Färbungen mit anschließender Analyse am Durchflusszytometer durchgeführt werden. Da die vorhandenen HIV-1-spezifischen Antikörper noch nicht für die Durchflusszytometrie getestet wurden, sollte dies vorher anhand transfizierter HEK293T Zellen erfolgen. Dazu wurden die HIV-1 Gene gag, nef und env von den Referenzviren SF2 und NL4-3 in den Vektor pegfp-c1 kloniert, so dass sie ein Fusionsprotein mit GFP (green fluorescence protein) bilden. Nach erfolgreicher Generierung der Plasmide wurden diese in HEK293T Zellen mit FuGENE transfiziert. Da die HIV-1 Gene als Fusionsprotein mit GFP gebildet werden, kann anhand der Fluoreszenz von GFP zum einen die erfolgreiche Transfektion detektiert werden und zum anderen, ob der zu untersuchende Antikörper bindet. Daher wurden die Zellen 48 Stunden nach Transfektion zunächst unter einem Fluoreszenzmikroskop betrachtet. Konnte man eine GFP-Fluoreszenz feststellen, wurden die Zellen permeabilisiert und intrazellulär mit den entsprechenden HIV-1-spezifischen Antikörpern und einem PEgekoppelten, sekundären Antikörper gefärbt. Bei der Analyse am Durchflusszytometer konnte anhand der transfizierten, doppelt fluoreszierenden Zellen erkannt werden, ob der Antikörper gebunden hat (Abbildung 4.1). 25
34 4 Ergebnisse Abbildung 4.1 Der Nef-Antikörper EH1 bindet spezifisch an HEK293T Zellen, die mit dem Plasmid pegfp-c1 SF2 Nef transfiziert wurden. SF2 Nef liegt als Fusionsprotein mit GFP vor. HEK293T wurden mit FuGENE transfiziert und bei positiver Fluoreszenz des GFP 48 Stunden nach Transfektion permeabilisiert und intrazellulär mit dem Nef-spezifischen Antikörper gefärbt. Somit konnte detektiert werden, ob der Antikörper spezifisch bindet und für Analysen am Durchflusszytometer geeignet ist. Der linke Dot Plot zeigt, dass sich in beinah allen transfizierten Zellen aufgrund der Färbung mit dem Nef-Antikörper eine Nef-Expression (y-achse) nachweisen lässt. Die Kontrollfärbungen auf untransfizierte HEK293T (rechts) und Zellen, die mit dem Vektor pegfp-c1 transfiziert wurden (Mitte), demonstrieren keine Nef-Expression. Für Gag- und Env-spezifische Antikörper wurde ebenso verfahren. Insgesamt konnte für Nef ein Antikörper (Nef EH1), für Gag zwei Antikörper (p24 AG3.0, p24 183/H12) und für Env gp120 drei Antikörper (gp120 AD3, gp D, gp Chessie 6) ermittelt werden, die spezifisch an die jeweiligen Proteine binden und für Analysen am Durchflusszytometer genutzt werden können. Durch Titration konnte ich zudem die geeignete Antikörperkonzentration bestimmen. Alle Antikörper und deren zu verwendende Konzentration sind in der Tabelle 6.22 zusammengefasst Elektroporation der HIV-1-spezifischen mrnas zeigt schnelle und effiziente Proteinexpression Nach Elektroporation der PBMC mit HIV-1-spezifischen mrnas und GFP mrna als Kontrolle wurden die Zellen für eine intrazelluläre Färbung mit HIV-1-spezifischen Antikörpern und anschließender Durchflusszytometrie verwendet. Hiermit sollte überprüft werden, ob die Elektroporation erfolgreich war und die mrna-kodierten Proteine in der Zelle exprimiert werden. Dabei konnte bereits nach vier Stunden eine deutliche Expression der Proteine GFP, Gag und Nef detektiert werden (Abbildung 4.2 A). Betrachtet man die Mediane sind 90% der PBMC positiv für GFP (range:67% -93%), 65% der Zellen exprimieren das Gag-Protein (range: 4%-88%) und 32% das Nef-Protein (range: 1%-89%). Hierbei fällt auf, dass in den meisten Versuchen die Expression des 26
35 4 Ergebnisse Nef-Proteins deutlich geringer ist als die Expression des Gag-Proteins. GFP zeigte konstant hohe Expressionswerte. Da PBMC aus vielen Subpopulationen bestehen, ist neben der Elektroporationseffizienz der Gesamt-PBMC auch die Effizienz in den verschiedenen Zelltypen innerhalb der PBMC interessant. Auf Grund dessen analysierte ich die Expression der Proteine GFP und SF2 Gag in CD4 + T-Lymphozyten, CD8 + T-Lymphozyten, CD19 + B-Zellen und CD14 + Monozyten. Alle Subpopulationen waren etwa zu 90% positiv für GFP, wohingegen die Expression des Gag-Proteins in allen Populationen mit Ausnahme der CD14 + Monozyten geringer war (Abbildung 4.2 B). Die Elektroporationseffizienz mit mrna ist in T-Lymphozyten und Monozyten sehr gut. Für B-Zellen wurde eine gute Transfektionseffizienz von GFP beobachtetet, vergleichbar zu T-Zellen, während Gag- Protein in B-Zellen deutlich schlechter exprimiert wurde. Abbildung 4.2 Expression der mrna-kodierten Proteine GFP, HIV-1 Gag und HIV-1 Nef in PBMC von Normalspendern und HIV-1-infizierten Patienten. Die Expression wurde 4 Stunden nach Elektroporation mittels intrazellulärer Färbung und Durchflusszytometrie bestimmt. A: Repräsentative Histogramme zeigen die Fluoreszenzintensität in PBMC, die mit GFP-, Gag- oder Nef-kodierender mrna elektoporiert wurden (schwarz) im Gegensatz zur Mock-Elektroporation (grau). B: Expression von GFP und SF2 Gag in den unterschiedlichen Subpopulationen der PBMC. PBMC wurden mit der entsprechenden mrna elektroporiert, und nach 4 Stunden mit einem Gag-spezifischen Antikörper und verschiedenen CD-Markern gefärbt. Dargestellt sind die prozentualen Frequenzen der GFP- bzw. Gag-exprimierenden Zellen innerhalb der verschiedenen Subpopulationen. Die Grafik zeigt den Median aus drei Experimenten mit den Interquartilen. C: Die Verteilung der PBMC-Subpopulationen wird durch die Elektroporation nicht verändert. Dargestellt sind die Median-Werte aus drei Experimenten mit den Interquartilen. E-PBMC= elektroporierte PBMC. 27
36 4 Ergebnisse Des Weiteren ist wichtig, dass sich innerhalb der PBMC die Verteilung der Subpopulation nicht durch die Elektroporation verändert. Daher wurden unbehandelte, mockelektroporierte und SF2 gag mrna-elektroporierte PBMC 24 Stunden nach Elektroporation mit verschiedenen CD-Antikörpern gefärbt und am Durchflusszytometer vermessen. Wie in Abbildung 4.2 C dargestellt, wiesen die Frequenzen der PBMC- Subpopulationen keine Veränderungen nach Elektroporation auf. Somit kann konstatiert werden, dass keine der Subpopulationen selektiv durch die Elektroporation getötet wird PBMC exprimieren HIV-1-Proteine bis zu sechs Tagen nach Elektroporation Nachdem eine deutliche Proteinexpression bereits vier Stunden nach Elektroporation detektiert werden konnte, stellte sich die Frage, wie lange die exogenen Proteine in den elektroporierten PBMC vorhanden sind. Deswegen wurden PBMC mit mrna elektroporiert und anschließend die Proteinexpression zu bestimmten Zeitpunkten intrazellulär mit spezifischen Antikörpern bestimmt. In Abbildung 4.3 ist ein repräsentatives Beispiel aus drei Kinetikversuchen mit ähnlichen Ergebnissen dargestellt. Die GFP-Expression ist sehr stabil. Auch sechs Tage nach Elektroporation waren noch über 80% der Zellen positiv für GFP. Für das HIV-1 Gag-Protein ließ sich die höchste Proteinexpression nach 24 Stunden messen. Zu diesem Zeitpunkt exprimierten 61% der PBMC Gag. Anschließend sank die Gag-Proteinmenge kontinuierlich, dennoch exprimierten 140 Stunden nach Elektroporation noch 15% der PBMC das Gag-Protein. Die HIV-1 Nef-Proteinmenge erreichte ein Expressionsmaximum nach 48 Stunden. Danach fiel die Expression sehr schnell ab, so dass nach 140 Stunden kein Nef-Protein mehr nachweisbar war. Diese Ergebnisse demonstrieren, dass mrna-kodierte Proteine sehr schnell in den Zellen synthetisiert werden und für mehrere Tage nach der Elektroporation in PBMC vorhanden sind. 28
37 4 Ergebnisse Abbildung 4.3 Expressionskinetik der mrna-kodierten Proteine GFP, HIV-1 Gag und HIV-1 Nef. PBMC wurden mit den entsprechenden mrnas elektroporiert. Zu den angegebenen Zeitpunkten wurden die Zellen intrazellulär mit Gag- bzw. Nef-spezifischen Antikörpern gefärbt und durchflusszytometrisch bestimmt. Die gezeigte Darstellung repräsentiert eins aus drei Experimenten mit ähnlichen Ergebnissen mrna-kodierte HIV-1-Proteine sind in vitro immunogen und führen zur Freisetzung von γ-ifn PBMC von HIV-1-infizierten Spendern wurden mit mrna elektroporiert und vier Stunden später in einen γ-ifn ELISPOT Assay eingesetzt, um die Immunogenität der mrnakodierten Proteine zu testen. Bei den meisten Patienten konnte nach Elektroporation mit HIV-1-spezifischer mrna eine deutliche Sekretion von γ-ifn beobachtet werden (Abbildung Abbildung 4.9). Durchgeführte Kontrollen wie die Elektroporation ohne mrna (Mock) und mit GFP mrna induzierten in allen durchgeführten Experimenten keine Zytokinfreisetzung, so dass die Immunaktivierung durch die mrna-kodierten HIV-1-Proteine spezifisch ausgelöst wird und nicht durch unspezifische Signale, die bei der mrna-elektroporation entstehen. Jedoch führte bei einigen Patienten die Elektroporation mit HIV-1-spezifischer mrna trotz nachgewiesener Proteinexpression zu keiner oder nur einer geringen γ-ifn Produktion. Dies war bei den Patienten 0011, 0545 und 0899 der Fall, bei welchen jeweils autologe nef mrna und SF2 nef mrna getestet wurden. Für die Patienten 0545 und 0899 konnte im γ-ifn ELISPOT Assay auch keine Reaktion gemessen werden, wenn PBMC mit HIV-1-Peptiden inkubiert wurden, wohingegen 0011 eine gute Reaktion auf das Nef-Peptid FL8 zeigte. 29
38 4 Ergebnisse Das autologe Virus löst in fünf von zehn Fällen eine stärkere Immunantwort aus als Referenzviren PBMC von HIV-1-Patienten, von welchen eine autologe HIV-1 mrna vorhanden ist (siehe Tabelle 4.1), wurden sowohl mit autologer gag oder nef mrna sowie mit mrna von HIV-1 Referenzviren elektroporiert. Nach vier Stunden wurden 2x10 5 Zellen pro Well im γ-ifn ELISPOT Assay analysiert, um die T-Zellantwort gegen autologe virale Proteine mit der T-Zellantwort gegen heterologe virale Proteine zu vergleichen. Zudem wurde stets die Expression der mrna-kodierten Proteine bestimmt. Eine Übersicht zu den Patientendaten ist in Tabelle 4.2 gegeben. Tabelle 4.2 Patientendaten, zu welchen die T-Zellantwort gegen das autologe Virus bestimmt wurde. Die Daten beziehen sich auf den Zeitpunkt des dargestellten γ-ifn ELISPOT Assays. Patienten ID 0011 M 0143 W 0225 M 0284 M 0335 W 0347 M 0545 M 0899 M 0925 M Geschlecht HLA Klasse I ART A*01, A*24, B*08, B*61,Cw*07, Cw*02 A*02, A*03, B*07, B*60, Cw*03, Cw*07 A*02, A*03, B*07, B*18, Cw*05, Cw*07 A*02, A*11, B*39, B*57,Cw*06,Cw*07 A*02, A*03, B*62, B*60, Cw*03, Cw*03 A*02, A*24, B*13, B*18, Cw*06, Cw*07 A*02, A*02, B*75, B*62, Cw*03, Cw*08 A*01, A*02, B*51, B*52, Cw*07, Cw*12 A*01, A*66, B*41, B*44, Cw*16, Cw*17 ATV, 3TC/ AZT Dauer der ART [Jahren] Viruslast [Kopien/ml] CD4/µl 8 < keine / EFV, TDF, FTC keine / keine / TDF, FTC, RAL TDF/ FTC, EFV TDF/ FTC, EFV 9 < ,5 < < keine / ART: Antiretrovirale Therapie, ATV: Atazanavir, EFV: Efavirenz, RAL: Raltegravir, TDF: Tenofovir DF, FTC: Emtricitabin, AZT: Zidovudin, 3TC: Lamivudin Patient 0011 Da für diesen Patienten nur autologe nef mrna vorhanden war, wurde die Nefspezifische T-Zellantwort nach Elektroporation mit autologer nef und SF2 nef mrna miteinander verglichen. Allerdings wurde nach Elektroporation der nef mrnas trotz 30
39 4 Ergebnisse nachgewiesener Proteinexpression kaum eine messbare T-Zellantwort quantifiziert (SF2 Nef: 20%; autologes Nef: 40%). Da die Immunantworten so gering waren, konnte kein Vergleich der beiden Nef-Proteine durchgeführt werden (Abbildung 4.4 A). Auch in wiederholten Versuchen konnte keine γ-ifn Freisetzung detektiert werden. Die Inkubation der PBMC mit dem Peptid FL8 (FLKEKGGL) resultierte hingegen in einer guten Nefspezifischen T-Zellantwort (30 SFU=spot forming units) (Abbildung 4.4 A). Das Peptid entspricht der Sequenz eines HLA-B*08 Epitops (113). Der Patient wurde bereits seit mehreren Jahren antiretroviral therapiert, was in der Regel mit einer Reduktion der Zahl an zirkulierenden HIV-1-spezifischen CTLs einhergeht (114). Patient 0143 Trotz großer Sequenzunterschiede zwischen dem autologen Nef von Patient 0143 und SF2 Nef (Abbildung 4.10 B) konnte nur eine geringe Divergenz in der T-Zellantwort auf die beiden Nef-Proteine festgestellt werden (Abbildung 4.4 B). SF2 Nef induzierte 14 SFU, autologes Nef 21 SFU (Abbildung 4.4 B). Allerdings wurden die beiden Nef-Proteine unterschiedlich exprimiert. So exprimierten nur 11% SF2 Nef, 28% der Zellen hingegen exprimierten das autologe Nef-Protein. Dadurch kann die etwas bessere Immunantwort gegen das autologe Nef-Protein von der stärkeren Expression stammen. Das Peptid Nef KL9 (KEKGGLEGL), ein HLA-B*60-restringiertes Epitop (115), zeigte mit 68 SFU eine klare Nef-spezifische T-Zellantwort (Abbildung 4.4 B). 31
40 4 Ergebnisse Abbildung 4.4 Vergleich der T-Zellantwort der Patienten 0011 und 0143 gegen autologe und heterologe HIV-1-Proteine im γ-ifn ELISPOT Assay. PBMC von HIV-1-infizierten Patienten wurden mit autologer und heterologer HIV-1-spezifischer mrna elektroporiert und nach vier Stunden in einem ELISPOT Assay analysiert. A: Die Nef-spezifische T-Zellantwort von Patient 0011 war sehr gering, so dass eine vergleichende Aussage nicht möglich war. B: Patient 0143 zeigte eine etwas stärkere T-Zellantwort gegen das autologe Nef- Protein. SFU=spot forming units Patient 0225 Nach Elektroporation der PBMC dieses antiretroviral therapierten Patienten mit gag mrna konnte eine starke T-Zellantwort mit 57 SFU auf das autologe Gag-Protein im ELISPOT detektiert werden. SF2 Gag löste mit 16 SFU eine deutlich schwächere Immunantwort aus (Abbildung 4.5 A). Beide Proteine wurden von den Zellen gleich stark exprimiert (SF2 Gag: 79%, autologes Gag: 82%). Gegen das p17 KL14 Peptid (KIRLRPGGKKKQKL) konnte eine gute Immunstimulation gezeigt werden (Abbildung 4.5 A). Dieses enthält die beiden HLA-A*03-restringierten Epitope RK9 (RLRPGGKKK) und KK9 (KIRLRPGGK) (116, 117). Da die Reaktion gegen das Peptid p17 KL14 stärker als die Summe der Reaktionen gegen RK9 und KK9 war, kann vermutet werden, dass das p17 KL14 Peptid möglicherweise weitere, nicht definierte CTL-Epitope bzw. auch HLA-II- Epitope enthält. 32
41 4 Ergebnisse Abbildung 4.5 Vergleich der T-Zellantworten des Patienten 0225 gegen autologe und heterologe HIV-1- Proteine im γ-ifn ELISPOT Assay. A: Die Gag-spezifische Immunantwort des Patienten war deutlich stärker gegen das autologe Gag-Protein. B: Die Nef-spezifische Immunantwort des Patienten war geringer auf das autologe Nef-Protein. Im Hinblick auf die Nef-spezifische T-Zellantwort trat genau das Gegenteil auf. Das autologe Nef aktivierte weniger T-Zellen (8 SFU) als SF2 Nef (37 SFU) (Abbildung 4.5 B), obwohl SF2 Nef von weniger Zellen exprimiert wurde (25%) als das autologe Nef (57%). In Abbildung 4.10 ist ein Sequenzalignment der Aminosäuren der verschiedenen Nef- Proteine dargestellt. In der autologen Nef Sequenz von Patient 0225 findet man in nur einem relevanten CTL-Epitop eine Mutation. Das HLA-A*03-restringierte Epitop QK10 (QVPLRPMTYK) (118) hat an Position acht einen Threonin zu Asparagin Austausch, der zum Verlust der Erkennung führt. Dies wurde sowohl durch die Inkubation der PBMC des Patienten mit den verschiedenen Peptiden nachgewiesen (Abbildung 4.5 B) als auch mit QK10-spezifischen CTL Linien des Patienten (Abbildung 4.6 A). In beiden Fällen resultierte die Stimulation mit dem Peptid QK10 T/N, welches die Mutation des autologen Virus enthält, in einer fehlenden oder schwächeren T-Zellantwort als die Stimulation mit dem Wildtyp Peptid. Dies zeigt zum einen, dass es sich hierbei um eine CTL Fluchtmutante handelt und zum anderen, dass diese Mutation Grund der schlechteren Erkennung des autologen Nef-Proteins ist. 33
42 4 Ergebnisse Das Immunsystem des Patienten ist allerdings nachgereift, um sich an die veränderten Verhältnisse anzupassen. So konnten drei Jahre nach Plasmaentnahme, welches zur Isolation der viralen RNA und Generierung der autologen mrna diente, CTLs gegen die mutierte Variante Nef QK10 T/N detektiert werden. Sowohl die Inkubation mit dem Wildtyp Peptid QK10 als auch mit dem mutierten Peptid QK10 T/N führten zur Stimulation von spezifischen CTL Linien (Abbildung 4.6 B). Da nur die QK10 T/N-spezifische CTL Linie eine geringe Kreuzreaktivität zu QK10 zeigte, kann man davon ausgehen, dass es sich um zwei verschiedene TCRs handelt und das Immunsystem von 0225 einen neuen TCR gegen das mutierte Epitop gebildet hat (Abbildung 4.6). Abbildung 4.6 Nef-spezifische CTL Linien des Patienten 0225 zeigten zunächst nur die Erkennung des Wildtyp Peptids von QK10, zu einem späteren Zeitpunkt konnte das Immunsystem nachreifen und es ließen sich CTLs gegen das mutierte Epitop QK10 T/N detektieren. QK10-spezifische (schwarze Balken) bzw. QK10 T/N-spezifische (graue Balken) CTL Linien wurden durch Inkubation der PBMC von Patient 0225 mit den jeweiligen Peptiden generiert. Das Peptid Nef QK10 T/N entspricht der autologen Nef Sequenz des Patienten und stellt eine CTL Fluchtmutante dar. Gezeigt ist hier das Ergebnis einer γ-ifn ELISPOT Analyse mit 1x10 5 Zellen pro Well. A: Ende des Jahres 2010 konnten nur CTLs gegen das Wildtyp Peptid QK10 quantifiziert werden, die keine Kreuzreaktion zu der mutierten Form zeigten. Das autologe Epitop QK10 T/N enthält eine CTL Fluchtmutante und wurde nicht erkannt. B: Anfang des Jahres 2011 wurden CTLs gegen das mutierte Epitop detektiert. Zudem zeigte die QK10 T/N-spezifische CTL Linie eine mäßige Kreuzerkennung des QK10 Peptids. Das Immunsystem ist somit nachgereift und hat einen neuen TCR für das mutierte Epitop gebildet. Patient 0284 Für den noch unbehandelten Patient 0284 konnte autologe gag und nef mrna verwendet werden. Die T-Zellantwort gegen autologes Gag war mit 275 SFU mehr als doppelt so stark wie gegen SF2 Gag mit 109 SFU (Abbildung 4.7 A). Die Expression der beiden Proteine war gleich. SF2 Gag wurde von 59% der PBMC exprimiert, autologes Gag von 64% der Zellen. Die eingesetzten Gag-Peptide induzierten eine sehr starke 34
43 4 Ergebnisse Immunantwort, insbesondere das Peptid Gag07-41, welches das bekannte HLA-B*57- restringierte CTL-Epitop KF11 (KAFSPEVIPMF) enthält (119). Die summierten Frequenzen der Gag-spezifischen T-Zellen, welche durch die Inkubation der PBMC mit diversen Gag-Peptiden beobachtet wurden, überstiegen die Anzahl der Gag-spezifischen T-Zellen, die durch das Protein Gag stimuliert wurden. Dies lässt auf eine bessere Präsentation der exogen zugefügten Peptide schließen. Die Nef-spezifische T-Zellantwort von diesem Patient fiel schwächer aus als die Gagspezifische T-Zellantwort, was vermutlich durch eine geringe Nef-Proteinexpression in den nef mrna-elektroporierten PBMC verursacht wurde. SF2 Nef wurde von 7% der Zellen exprimiert, autologes Nef von 9% der Zellen. Trotz dieser ähnlichen, sehr niedrigen Proteinexpression induzierte autologes Nef eine zweimal stärker T-Zellantwort als SF2 Nef (Abbildung 4.7 B). Die Stimulation der PBMC mit Nef-Peptiden zeigte eine sehr gute Immunantwort (Abbildung 4.7 B). Abbildung 4.7 Vergleich der T-Zellantworten des Patienten 0284 gegen autologe und heterologe HIV-1- Proteine im γ-ifn ELISPOT Assay. Sowohl die Gag-spezifische (A) als auch die Nef-spezifische (B) T-Zellantwort war stärker gegen das autologe Virus. 35
44 4 Ergebnisse Patient 0335 PBMC des Patienten 0335 zeigten mit 77 SFU eine stärkere Antwort gegen das autologe Gag-Protein als gegen das SF2 Gag-Protein mit 54 SFU (Abbildung 4.8 A). Dabei exprimierten 63% der Zellen das autologe Gag-Protein und 55% das SF2 Gag. Durch die Verwendung von Gag-Peptidpools, die aus jeweils fünf Peptiden bestehen und SF2 entsprechen, und weiteren kürzeren Peptiden konnte das HLA-A*02-restringierte Epitop YL9 (YVDRFYKTL) (120) als einziges CTL-Epitop in SF2 Gag identifiziert werden, welches von diesem Patienten erkannt wurde (Abbildung 4.8 A). Dieses Epitop ist zwischen dem autologen und SF2 Gag-Protein konserviert (Abbildung 4.10 A). Folglich könnte die stärkere T-Zellantwort gegen das autologe Gag ein Resultat von weiteren CTL- Epitopen sein, die in SF2 Gag nicht enthalten sind, oder aber das Resultat einer besseren Prozessierung und Präsentation des YL9-Epitops im autologen Kontext. Beide Nef-Proteine stimulierten eine sehr geringe spezifische T-Zellantwort in Patient Die von SF2 Nef induzierte Antwort (9 SFU) war dabei stärker als die von dem autologen Nef (3 SFU) (Abbildung 4.8 B), wobei die Expression des SF2 Nef-Proteins (20%) etwas höher war als die Expression des autologen Nef (12%). Der Grund für diese sehr geringe Immunantwort nach nef mrna-elektroporation könnte die schwache Expression der Nef-Proteine sein. Da das Peptid AK9 (AVDLSHFLK) für das HLA-B*62- restringierte Epitop (121) eine sehr starke T-Zellantwort mit 220 SFU induzierte (Abbildung 4.8 B), kann ausgeschlossen werden, dass der Patient generell nur eine sehr schwache Immunantwort gegen Nef aufweist. In dem Epitop findet sich sowohl in der autologen als auch in der SF2 Sequenz ein V-zu-L Austausch an Position 2 (Abbildung 4.10 B), was bei früheren Analysen der Arbeitsgruppe nur zu einer leichten Verminderung der Erkennung durch AK9-spezifische, HLA-B*62-restringierte CTL Linien führte, was die schwache Immunantwort auf die beiden Nef-Proteine daher alleine vermutlich nicht erklärt. Leider konnten hierzu keine weiteren Untersuchungen durchgeführt werden, da der Patient im Verlauf des Projektes verstarb. 36
45 4 Ergebnisse Abbildung 4.8 Vergleich der T-Zellantworten des Patienten 0335 gegen autologe und heterologe HIV-1- Proteine im γ-ifn ELISPOT Assay. A: Die Gag-spezifische Immunantwort des Patienten war stärker gegen das autologe Gag-Protein. B: Die Nef-spezifische Immunantwort des Patienten war geringer auf das autologe Nef-Protein, jedoch fiel die Immunantwort auf das SF2 Nef-Protein ebenfalls sehr gering aus. Patient 0347 Die spezifische T-Zellantwort des Patienten 0347 war mit 10 SFU sehr schwach gegen das autologe Gag-Protein. Hingegen führte SF2 Gag zu einer deutlich größeren Freisetzung von γ-ifn mit 34 SFU (Abbildung 4.9). Die Proteinexpression war beinahe gleich, da 81% der PBMC das autologe Gag exprimierten und 76% SF2 Gag. Der Vergleich der Aminosäurensequenzen der beiden Proteine offenbarte neben anderen Mutationen eine CTL Fluchtmutation in einem HLA-B*13-Epitop namens p15 RI9 (RQANFLGKI) (122). In der autologen Gag Sequenz befindet sich eine Lysin (K) zu Arginin (R) Substitution an Position acht (Abbildung 4.10 A). Die Inkubation von PBMC des Patienten mit dem Wildtyp Peptid p15 RI9 und dem Peptid p15 RI9 K/R, welches die Mutation des autologen Virus enthält, zeigte eine starke Abnahme der T-Zellantwort gegen das Peptid p15 RI9 K/R (Abbildung 4.9 A). CTL Linien des Patienten bestätigten diese Beobachtung. Nur die Inkubation mit dem Wildtyp Peptid p15 RI9 führte zur Generierung einer positiven CTL Linie (Daten nicht gezeigt). Die Inkubation mit dem 37
46 4 Ergebnisse Peptid p15 RI9 K/R zeigte keine Reaktion, so dass es sich hierbei um eine CTL Fluchtmutante handelt. Die Mutation p15 RI9 K/R in der autologen Gag Sequenz von 0347 könnte somit ein Grund für die schlechtere T-Zellantwort gegen das von der mrnakodierte autologe Gag-Protein sein. Patienten 0545 und 0899 Wie bereits unter erwähnt, konnte für diese beiden Patienten keine T-Zellantwort im γ-ifn ELISPOT Assay detektiert werden. PBMC wurden in beiden Fällen mit autologer nef und SF2 nef mrna elektroporiert. Trotz nachgewiesener Proteinexpression führten die mrna-kodierten HIV-1-Proteine zu keiner Sekretion von γ-ifn. Jedoch induzierten auch die verwendeten Nef-Peptide keine oder nur eine sehr geringe T-Zellantwort mit maximal 13 SFU. Beide Patienten wurden seit längerem bereits mit einer effektiven antiretroviralen Therapie behandelt. Patient 0925 Die beobachtete Gag-spezifische T-Zellantwort in PBMC des Patienten 0925 fiel relativ schwach aus. Zudem induzierten autologes Gag und SF2 Gag eine identische Immunantwort mit 18 SFU (Abbildung 4.9 B). In 60% der Zellen wurde das autologe Gag- Protein exprimiert, in 50% das SF2 Gag-Protein. Durch den Einsatz von Peptiden konnte die Erkennung von zwei CTL-Epitopen in Gag nachgewiesen werden: das Peptid Gag07-2, welches das HLA-B*40 Epitop GELDRWEKI (120) enthält, und das Peptid p24/3 mit dem HLA-B*44 Epitop EEKAFSPEV (123) (Abbildung 4.9 B). Obwohl autologe nef mrna des Patienten vorhanden war, konnte die T-Zellantwort gegen das autologe Nef im Rahmen dieser Arbeit nicht mehr analysiert werden, da der Patient mit HAART behandelt wurde und in Folge dessen keine Immunantworten mehr ohne Vorstimulation der Zellen im γ-ifn ELISPOT zeigte. 38
47 4 Ergebnisse Abbildung 4.9 Vergleich der T-Zellantworten der Patienten 0347 und 0925 gegen autologe und heterologe HIV-1-Proteine im γ-ifn ELISPOT Assay. A: Die Gag-spezifische Immunantwort des Patienten 0347 war geringer gegen das autologe Gag-Protein. B: Die Gag-spezifische Immunantwort des Patienten 0925 war auf autologes und heterologes Gag-Protein gleich. Reproduzierbarkeit und Zusammenfassung Für jeden Patienten wurden die Versuche mindestens zu zwei verschiedenen Zeitpunkten durchgeführt. Das Erkennungsmuster der autologen mrna gegenüber der heterologen mrna konnte in allen Fällen reproduziert werden. Die Unterschiede bezüglich der Proteinexpression zwischen den Zeitpunkten lagen in der Mehrzahl der Experimente (>80%) unter 10% Abweichung. Dies weist auf eine gute Reproduzierbarkeit des Assays hin. Bei den Patienten 0545 und 0899 wurde der Versuch aufgrund der komplett fehlenden T-Zellantwort nicht wiederholt. Die Analyse der T-Zellantwort gegen das autologe Nef-Protein des Patienten 0335 konnte nicht wiederholt werden, da der Patient verstarb. Insgesamt konnten mit PBMC, welche mit autologer HIV-1-spezifischer mrna elektroporiert waren, sehr effizient die CD8 + T-Zellantwort gegen autologe Virusproteine analysiert werden. Für fünf Patienten konnte eine stärkere T-Zellantwort gegen autologe virale Proteine im Vergleich zur T-Zellantwort gegen Proteine des HIV-1 Referenzstamms SF2 beobachtet werden (Nef von Patient 0143, Gag und Nef von Patient 0284, Gag von 39
.")
48 4 Ergebnisse Patient 0225 und Gag von Patient 0335). Eine geringere T-Zellantwort gegen autologes Virusprotein konnte in zwei Patienten mit Fluchtmutationen in CTL-Epitopen assoziiert werden (Nef 0225 und Gag 0347). Patient 0335 wies ebenfalls eine geringere Immunantwort gegen das autologe Nef-Protein auf, jedoch konnte hierfür keine Erklärung gefunden werden. Die weiteren Patienten zeigten entweder keine T-Zellreaktion auf die durch mrna-elektroporation in die Zellen eingebrachten HIV-1-Proteine (0545 und 0899) oder es konnte kein Unterschied in der Erkennung zwischen autologen und heterologen Proteinen festgestellt werden (Nef 0011 und Gag 0925). 40
49 4 Ergebnisse Abbildung 4.10 Alignment der Aminosäurensequenzen der autologen Gag- (A) und Nef- (B) Proteine. Grau unterlegte Aminosäuren unterscheiden sich von der Consensus Sequenz. Rot eingerahmte Sequenzbereiche sind CTL-Epitope, welche für die Immunantworten der Patienten eine entscheidende Rolle spielen. Die Consensus Sequenz wurde von der Vector NTI Software generiert Vergleich der ausgelösten T-Zellantworten zwischen mrnakodierten HIV-1-Proteinen und Peptiden, Peptid-Pools und Vaccinia-Viren Zum Monitoring der T-Zellantworten von HIV-1-infizierten Patienten werden üblicherweise HIV-1-Peptide, Peptid-Pools oder rekombinante Vaccinia-Viren verwendet. Da mrnakodierte HIV-1-Proteine als alternative Antigene zum Immunmonitoring eingesetzt werden könnten, sollte die Kapazität der mrna-kodierten Proteine zur Stimulation von T-Zellen mit der von Standardantigenen verglichen werden. Dazu wurden PBMC von HIV-1- infizierten Patienten mit gag und nef mrna elektroporiert und in γ-ifn ELISPOT Assays ohne weitere Effektorzellen eingesetzt, um die Aktivierung von Gag- oder Nefspezifischen T-Zellen in den elektroporierten PBMC zu analysieren. Gleichzeitig wurden PBMC des Patienten auch mit einzelnen Peptiden, Peptid-Pools oder rekombinanten Vaccinia-Viren im ELISPOT Assay inkubiert. Die Elektroporation der PBMC mit HIV-1- spezifischer mrna führte zu einer deutlichen Stimulation der HIV-1-spezifischen T-Zellen und ist vergleichbar mit der T-Zellstimulation nach Inkubation mit Peptid-Pools (Abbildung 4.4 A). Jedoch zeigten einzelne Peptide eine höhere Kapazität HIV-1-spezifische T-Zellen zu stimulieren. Auch die Summe der T-Zellantworten gegen mehrere Peptide lag über der T-Zellantwort gegen HIV-1 mrna-kodierte Proteine (Abbildung 4.5 -Abbildung 4.9). Die 41
50 4 Ergebnisse Stimulation von T-Zellen nach Elektroporation mit HIV-1-spezifischer mrna ist deutlich stärker als die nach Infektion der PBMC mit rekombinanten Vaccinia-Viren (Abbildung 4.5 A). Weder mock-elektroporierte PBMC noch GFP mrna-elektroporierte PBMC bewirkten eine Freisetzung von γ-ifn, daher ist die T-Zellantwort auf gag oder nef mrna eine spezifische Immunantwort. Da die Infektion von PBMC mit Vaccinia-Viren weniger effizient ist als die von EBVtransformierten lymphoiden B-Zelllinien (B-LCL), haben wir die T-Zell-Erkennung von mrna-elektroporierten B-LCL mit der Erkennung von Vaccinia-infizierten B-LCL verglichen. Dafür wurde eine HLA-A*03-positive B-LCL mit SF2 gag mrna elektroporiert oder mit einem rekombinanten Vaccinia-Virus infiziert, welches SF2 Gag exprimiert. Daraufhin wurden die Zellen im γ-ifn ELISPOT Assay mit CTL Linien von verschiedenen HIV-1-Patienten, die positiv für das HLA-A*03-restringierte p17 Epitop KK9 (KIRLRPGGK) sind, inkubiert. Somit konnte die T-Zell-Erkennung durch die CTL Linien verglichen werden. Die verwendeten Negativkontrollen wie unbehandelte B-LCL, mockelektroporierte B-LCL, GFP mrna-elektroporierte B-LCL und B-LCL, die mit einem Kontroll Vaccinia-Virus (vsc8) infiziert wurden, lösten durch die Ko-Inkubation eine geringe T-Zellaktivierung aus und stellen Hintergrundwerte dar (Abbildung 4.11 A). Als Positivkontrolle dienten die Peptid-inkubierte CTL Linie und die CTL Linie, welche mit Peptid-inkubierten B-LCL ko-inkubiert wurden. Diese zeigten jeweils eine deutliche γ-ifn Freisetzung, die mit einem Anti-CD8-Antikörper geblockt werden konnte (Abbildung 4.11 A). SF2 gag mrna elektroporierte B-LCL wurden von beiden KK9-spezifischen CTL Linien merklich besser erkannt als SF2 Gag-Vaccinia infizierte B-LCL (Abbildung 4.11 A). Zwar könnte diese Differenz in der Erkennung auch ein Resultat der unterschiedlich starken Proteinexpression von SF2 Gag sein, da 70% der mrna-elektroporierten B-LCL positiv für das Gag-Protein waren und nur 50% der Vaccinia-infizierten B-LCL (Abbildung 4.11 B). Durch die Erhöhung der Infektionsdosis der Vaccinia-Viren wurde die Proteinexpression jedoch nicht erhöht, die Viabilität der Zellen allerdings deutlich vermindert. Insgesamt demonstrieren diese Daten, dass mrnas, die HIV-1-Proteine kodieren, als alternatives Antigen zum Immunmonitoring verwendet werden können, da sie gleiche Mengen an γ-ifn wie Peptidpools freisetzten und besser erkannt wurden als rekombinante Vaccinia-Viren. 42
51 4 Ergebnisse Abbildung 4.11 SF2 gag mrna-elektroporierte B-LCL wurden im Gegensatz zu SF2 Gag Vacciniainfizierten B-LCL besser von Gag-spezifischen CTL Linien erkannt. PBMC von zwei verschiedenen HIV- 1-infizierten Patienten wurden mit dem Peptid p17 KK9 stimuliert und für 10 Tage inkubiert. Ausgewachsene CTL Linien wurden mit dem Peptid p17 KK9 im γ-ifn ELISPOT Assay getestet. B-LCL wurden mock elektroporiert oder mit GFP bzw. SF2 gag mrna elektroporiert. Zudem wurden B-LCL mit einem SF2 Gag exprimierenden Vaccinia-Virus oder einem Kontroll Vaccinia-Virus (vsc8) infiziert (MOI=5). Vier Stunden nach Elektroporation bzw. 24 Stunden nach Infektion wurden die Zellen zur Analyse in einen γ-ifn ELISPOT Assay und zur intrazellulären Färbung verwendet. A: γ-ifn ELISPOT Assay, um die Erkennung der SF2 gag mrnaelektroporierten B-Zellen mit der Erkennung der SF2 Gag-Vaccinia infizierten B-LCL durch Gag-spezifische CTL Linien zu vergleichen. Es wurden 5x10 4 Zellen der CTL Linie mit 5x10 4 der B-LCL inkubiert. In diesem Beispiel wurden CTL Linien von zwei verschiedenen Patienten verwendet (schwarze bzw. weiße Balken). B: Intrazelluläre Färbung mit einem spezifischen p24 Antikörper und Durchflusszytometrie der elektroporierten und Vaccinia-infizierten B-LCL. Die Gag Expression ist auf der y-achse angegeben. Sowohl gag mrnaelektroporierte als auch Gag-Vaccinia infizierte B-LCL exprimierten das SF2 Gag-Protein, jedoch war die Expression in den gag mrna-elektroporierten B-LCL um 20% höher. B-LCL: EBV-transformierte B-lymphoblastoiden Zelllinien 43
52 4 Ergebnisse mrna-kodierte gp120-proteine induzieren geringe Immunantworten Es sollten ebenfalls autologe HIV-1 env gp120 Gene amplifiziert und für die mrna- Synthese kloniert werden. Allerdings gelang dies nur bei zwei HIV-1-infizierten Patienten (0225 und 0347, Tabelle 4.1). Trotz mehrfacher Abänderung des PCR- Amplifikationsprotokolls und der Verwendung von diversen PCR-Primern konnte kein weiteres Fragment erfolgreich amplifiziert werden. Ob und in welchem Ausmaß die autologen Env gp120-proteine T-Zellantworten induzieren, sollte in γ-ifn ELISPOT Assays gemessen werden. Dazu wurden PBMC von HIV-1-infizierten Patienten mit 0225 gp120 mrna elektroporiert und anschließend im ELISPOT inkubiert. Es konnte in allen durchgeführten Versuchen kaum eine Expression des gp120-proteins detektiert werden (n=4; gp120 exprimierende Zellen 0%-7,6%). Jedoch konnte in drei Patienten eine Immunantwort gegen das gp120-protein von 0225 Env nachgewiesen werden, so dass die fehlende Detektion der Expression eventuell auf die Variabilität des gp120-proteins (Abbildung 4.13) und eine dadurch fehlende Erkennung durch den Antikörper zurückzuführen ist. Patient 0566 zeigte dabei die geringste Reaktion mit 15 SFU/2x10 5 PBMC. Patient 0284 wies eine T-Zellantwort von 30 SFU/2x10 5 PBMC auf, was sogar einer stärkeren Reaktion entspricht als die CTL-Antwort, welche bei Patient 0225 selbst quantifiziert wurde (20 SFU/2x10 5 PBMC). Somit induzierte das autologe gp120 bei 0225 eine mäßige Immunantwort, wohingegen bei seinem Partner mit demselben Ausgangsvirus eine etwas bessere Reaktion beobachtet wurde. Autologes gp120 von 0284 stand nicht zur Verfügung. Das gp120-protein von 0566 ist nicht mit dem gp120-protein von 0225 verwandt. Daher zeigte 0566 nur geringe T-Zellantworten gegen das mrna-kodierte gp120 von Dass dennoch Immunreaktionen auftreten, lässt vermuten, dass CTL-Epitope, die über die HLA-Allele von 0566 restringiert sind, in gp120 von 0225 nicht mutiert sind und daher noch durch CTLs von 0566 erkannt werden können. 44
53 4 Ergebnisse Abbildung 4.12 Env gp120 löste nur geringe Immunantworten aus, sowohl nach Peptidstimulation als auch nach Elektroporation mit gp120-spezifischer mrna. PBMC von HIV-1-infizierten Patienten wurden zum einen mit Env-Peptiden im γ-ifn ELISPOT Assay inkubiert und zum anderen mit env gp120-spezifischer mrna elektroporiert und anschließend im γ-ifn ELISPOT Assay inkubiert. Als Kontrollen dienten mock- und GFP mrna-elektroporierte PBMC. Es wurden stets 2x10 5 PBMC im ELISPOT eingesetzt und die autologe gp120 mrna von 0225 zur Elektroporation verwendet. A: Patient 0225 zeigte auf sein autologes gp120 eine geringe T-Zellreaktion mit 20 SFU. Die Inkubation mit zwei Env-Peptiden löste keine Reaktion aus. B: Patient 0566 zeigte ebenfalls keine Immunantwort nach Peptidinkubation, aber eine leichte T-Zellantwort gegen das gp120 des Patienten 0225 mit 15 SFU. C: Patient 0284 zeigte von den drei Patienten die stärkste T-Zellantwort gegen das gp120 von Es handelt sich dabei um das gp120 seines Partners, was somit vom selben Ausgangsvirus stammte. Die Inkubation der PBMC mit ausgewählten Env-Peptiden induzierte keine bis schwache Reaktionen (max. 8 SFU/2x10 5 PBMC). Die Env-Peptide entsprechen der Consensus B bzw. HXB2 Sequenz und wurden anhand der HLA-Allele des Patienten ausgewählt, da sie potentielle CTL-Epitope für diese HLA-Allele enthielten. Aufgrund der schlechten Nachweisbarkeit des Env-Proteins mit diversen Antikörpern und der geringen Immunogenität wurden keine weiteren Analysen dazu durchgeführt. 45

54 4 Ergebnisse Abbildung 4.13 Alignment der Aminosäurensequenzen des gp120-proteins von Env zeigen die hohe Variabilität des Gens. Die gp120 Sequenzen der Referenzviren NL4-3 und SF2 wurden mit den autologen gp120 Sequenzen von 0225 und 0347 verglichen. Es wurde die Software Vector NTI verwendet, welche auch die angegebene Consensus-Sequenz generierte. Grau unterlegt sind Aminosäuren oder Abschnitte, in welchen die Sequenzen nicht übereinstimmen. Bereits der Vergleich von NL4-3 und SF2 offenbart die hohe Variabilität Geringe Mengen an HIV-1-spezifischer mrna reichen aus, um eine Proteinexpression und T-Zellstimulation zu erhalten Welche Mindestmenge an mrna zur Elektroporation benötigt wird, um eine Proteinexpression zu detektieren und eine T-Zellantwort zu erhalten, sollten Titrationsversuche klären. Dazu wurden PBMC von HIV-negativen und HIV-positiven Spendern mit verschiedenen Mengen an mrna elektroporiert, nach vier Stunden intrazellulär gefärbt und durchflusszytometrisch analysiert. Eine Fluoreszenz für GFP konnte in über 80% der PBMC bis zu einer Menge von 2,5 µg nachgewiesen werden. Bei Verwendung von 0,5 µg GFP mrna fluoreszierten immerhin noch 51% der Zellen (Abbildung 4.14 A). Die Frequenz der SF2 Gag-exprimierenden PBMC fällt von 62% bei der Verwendung von 10 µg mrna auf 16% bei der Verwendung von 1 µg mrna (Abbildung 4.14 A). Da in den meisten Experimenten weniger Nef-Protein als Gag-Protein nachweisbar war und ausgeschlossen werden sollte, dass es sich hierbei eventuell um eine unterschiedliche Sensitivität der Antikörper handelt, kamen in der Titration der nef mrna zusätzlich noch steigende Mengen zum Einsatz. Dabei zeigte sich, dass der 46
55 4 Ergebnisse Einsatz von mehr mrna auch in einer höheren Proteinexpression resultierte. Während 15 µg SF2 nef mrna zu 27% Nef-exprimierenden Zellen führten, wurde durch die Verwendung von 30 µg mrna die Frequenz der Nef-exprimierenden Zellen mit 50% verdoppelt (Abbildung 4.14 B). Dies würde eher gegen eine geringere Sensitivität des Nef-Antikörpers sprechen. Abbildung 4.14 Proteinexpression von mrna-elektroporierten PBMC in Titrationsversuchen. PBMC von Normalspendern wurden mit unterschiedlichen Mengen an GFP, SF2 gag und SF2 nef mrna elektroporiert. Anschließend wurde die Expression nach Färbung mit spezifischen Antikörpern am Durchflusszytometer analysiert. A: Proteinexpression der Proteine GFP und SF2 Gag in PBMC nach Elektroporation mit verschiedenen Mengen an mrna. B: Proteinexpression von SF2 Nef nach Elektroporation mit unterschiedlicher Menge an mrna. Des Weiteren sollte bestimmt werden, wie viel mrna in die Zellen eingebracht werden muss, um eine ausreichende Proteinexpression für die Aktivierung von T-Zellen zu erhalten. Da die T-Zellantwort gegen das Gag-Protein deutlich stärker ausfällt und dadurch besser messbar ist, wurden von einem Patienten, von welchem eine gute Gag- Antwort bekannt war, PBMC mit verschiedenen Mengen an SF2 gag mrna elektroporiert und nach vier Stunden die γ-ifn Freisetzung im ELISPOT überprüft. Wie erwartet, konnte ein kontinuierlicher Abfall der SFU mit sinkender mrna-menge im ELISPOT beobachtet werden. Dennoch reichte 1 µg SF2 gag mrna aus, um nach Proteinexpression in den Zellen eine T-Zellaktivierung von 12 SFU zu erhalten (Abbildung 4.15). Diese Ergebnisse belegen, dass geringe Mengen an mrna zur Elektroporation ausreichend sind, um eine Proteinexpression detektieren zu können. Dazu reichen 0,5 µg GFP mrna und 1 µg gag mrna aus. Außerdem genügen 1 µg gag mrna, um durch die anschließende Proteinexpression in den Zellen eine messbare T-Zellaktivierung zu erhalten. 47
56 4 Ergebnisse Abbildung 4.15 Sekretion von γ-ifn nach Elektroporation von PBMC eines HIV-1-infizierten Patienten mit unterschiedlicher Menge an SF2 gag mrna. PBMC wurden mit 1 µg, 5 µg und 10 µg SF2 gag mrna elektroporiert. Nach 4 Stunden wurden 2x10 5 PBMC pro Well auf einen γ-ifn ELISPOT Assay gegeben, um die Induktion der γ-ifn Antwort zu messen Durch mrna-kodierte HIV-1-Proteine aktivieren HIV-1- spezifische CD8 + zytotoxische T-Lymphozyten ohne zusätzliche Effektorzellen Nachdem eine gute Immunogenität der HIV-1-spezifischen mrnas in vitro festgestellt wurde, wollten wir wissen, ob die Stimulation von HIV-1-spezifischen T-Zellen durch mrna-elektroporierte PBMC allein ausreicht oder ob noch zusätzliche Effektorzellen benötigt werden. Daher wurden PBMC eines HIV-1-positiven Spenders (0609) mit heterologer nef mrna elektroporiert. Diese elektroporierten PBMC wurden einmal mit autologen, unbehandelten PBMC und einmal ohne autologe, unbehandelte PBMC in einem γ-ifn ELISPOT Assay inkubiert. Als Kontrolle wurden Zellen mit dem Peptid Nef RI9 (RQEILDLWI) inkubiert, gegen welches der Patient eine bekannte T-Zellantwort besaß. Die Stimulation von PBMC mit dem Peptid Nef RI9 löste mit 609 SFU eine starke Immunreaktion aus. Wie erwartet resultierte die Verdoppelung der Zellzahl auf mit 1009 SFU etwa in der Verdopplung der Immunantwort (Abbildung 4.16). Eine starke T-Zellantwort mit einer deutlichen γ-ifn Freisetzung wurde auch durch die nef mrna-elektroporierten PBMC induziert (212 SFU). Während die Zugabe von unbehandelten PBMC zu den nef mrna-elektroporierten PBMC die T-Zellantwort verdoppelte, wurde die Immunreaktion mit 634 SFU verdreifacht, wenn man nef mrna-elektroporierte PBMC ohne die Zugabe von Effektorzellen einsetzte 48
57 4 Ergebnisse (Abbildung 4.16). Folglich werden keine zusätzlichen Effektorzellen benötigt, um eine T-Zellantwort zu induzieren. PBMC allein, welche mit HIV-1-spezifischer mrna elektroporiert wurden, sind zur Aktivierung der CTL-Antwort ausreichend und eine Steigerung der Sensitivität kann am besten durch die Erhöhung der Zellzahl an elektroporierten Zellen erreicht werden. Abbildung 4.16 γ-ifn-elispot Assay mit variierenden Zellzahlen zeigte, dass keine zusätzlichen Effektorzellen benötigt werden. PBMC eines HIV-1-infizierten Patienten (0609) wurden mock, mit GFP mrna oder mit heterologer nef mrna von Patient 0143 elektroporiert. Die Frequenz der Nef-spezifischen T-Zellen wurde vier Stunden nach Elektroporation im ELISPOT detektiert, indem unterschiedliche Zellzahlen eingesetzt wurden. Die Verwendung von 2x10 5 nef mrna-elektroporierten PBMC führte zu einer größeren T-Zellantwort als die Inkubation von 1x10 5 nef mrna-elektroporierten PBMC mit unbehandelten PBMC als Effektorzellen. Des Weiteren wurden PBMC von vier HIV-negativen Spendern mit HIV-1-spezifischen mrnas elektroporiert und im Anschluss auf Freisetzung von γ-ifn im ELISPOT untersucht. Im Gegensatz zu den PBMC der HIV-positiven Spender führten HIV-1- spezifische mrnas in PBMC von HIV-negativen Spendern zu keiner Produktion von γ-ifn. Dies beweist, dass die Immunreaktion, die in Zellen von Patienten ausgelöst wird, durch HIV-1-spezifische T-Zellen induziert wird und nicht durch eine unspezifische Aktivierung von T-Zellen. Daraufhin stellte sich die Frage, ob es sich bei den HIV-1-spezifischen T-Zellen, welche die Immunantworten auf die Antigen-kodierende mrna hervorrufen, um CD8 + zytotoxische T-Lymphozyten oder um CD4 + T-Lymphozyten handelt. Daher wurden PBMC von HIV-1-Patienten mit HIV-1-spezifischer mrna elektroporiert und anschließend 49
58 4 Ergebnisse im ELISPOT Assay mit blockierenden Antikörpern inkubiert. Es konnte eine starke Reduktion der ausgelösten Immunantworten festgestellt werden, wenn die Zellen mit einem Anti-CD8 Antikörper geblockt wurden (Abbildung 4.17). Die Inkubation der Zellen mit einem Anti-CD4 Antikörper zeigte einen geringen inhibitorischen Effekt. Dies konnte in zwei Patienten nachgewiesen werden. Somit lässt sich konstatieren, dass die meisten Immunantworten gegen Antigene, die durch HIV-1-spezifische mrnas kodiert werden, durch HIV-1-spezifische CTLs induziert werden. Abbildung 4.17 Durch die Elektroporation mit HIV-1-spezifischen mrnas wurden CD8 + HIV-1- spezifische T-Zellen stimuliert. 2x10 5 unbehandelte PBMC von HIV-1-positiven Spendern (hier: 0284) wurden mit 2x10 5 PBMC im γ-ifn ELISPOT Assay inkubiert, die entweder mit SF2 gag mrna oder autologer gag mrna elektroporiert wurden. Anschließend wurden blockierende Anti-CD4- bzw. Anti-CD8-Antikörper direkt zu den Zellen gegeben. Die Reduktion der Frequenz der SFU durch diese Antikörper zeigte, dass sowohl SF2 gag mrna elektroporierte PBMC sowie auch autologe gag mrna elektroporierte PBMC hauptsächlich von CD8 + T-Zellen erkannt werden und nur ein kleiner Anteil von CD4 + T-Zellen Neben γ-ifn induzieren die von HIV-1-spezifischen mrnas-kodierten Proteine die Sekretion von TNF-α Der Cytometric Bead Array (CBA) Human Th1/Th2 Cytokine Kit erlaubt die Messung mehrerer Zytokine, die in den Überstand der Zellen abgegeben werden können. Daher wurden vier bzw. 24 Stunden nach Elektroporation mit HIV-1-spezifischer mrna 50µl aus dem Überstand abgenommen und anschließend mit dem oben genannten Kit die Freisetzung der Zytokine IL-2, IL-4, IL-6, IL-10, TNF-α sowie γ-ifn analysiert. Allerdings konnte nur eine deutliche Freisetzung von TNF-α nach Elektroporation mit HIV-1- spezifischen mrnas gemessen werden, die anderen Zytokine zeigten keine messbare 50
59 4 Ergebnisse Produktion. Die Elektroporation mit gag oder nef mrna induzierte in allen Patienten eine Abgabe von TNF-α, wobei die Stärke der TNF-α Freisetzung zwischen den Patienten variierte. Außerdem wiesen die verschiedenen mrna-konstrukte eine unterschiedliche Kapazität zur Stimulation von TNF-α auf. In Abbildung 4.18 sind repräsentative Beispiele dargestellt. So rief SF2 nef mrna eine fast dreifach höhere Produktion von TNF-α hervor als SF2 gag (Abbildung 4.18 D). Die autologe gag mrna in Patient 0284 induzierte eine dreimal höhere Freisetzung von TNF-α als SF2 gag mrna (Abbildung 4.18 A), wohingegen die autologe gag mrna von Patient 0347 eine geringere Produktion im Vergleich zu SF2 gag mrna bewirkte (Abbildung 4.18 C). Die autologe nef mrna von Patient 0143 stimulierte eine ähnliche TNF-α-Sekretion wie SF2 nef mrna (Abbildung 4.18 B). Überdies wurden höhere Zytokinlevel vier Stunden nach Elektroporation als nach 24 Stunden detektiert. Unbehandelte PBMC, mock-elektroporierte sowie GFP mrnaelektroporierte PBMC stellen mit geringen Werten an TNF-α Hintergrundwerte dar. Eine Sekretion von TNF-α konnte zudem auch in PBMC von HIV-negativen Spendern nach Elektroporation mit HIV-1-spezifischen mrnas beobachtet werden (Abbildung 4.18 D). Daher kann man annehmen, dass die TNF-α Produktion zumindest in den HIVnegativen Spendern nicht durch Antigen-spezifische T-Zellen angeregt wird, sondern durch die Triggerung der Toll-like Rezeptoren TLR7 und TLR8. Diese beiden TLRs, die in diversen Zelltypen innerhalb der PBMC exprimiert sind, erkennen einzelsträngige RNA- Moleküle, welche bestimmte Nukleotidsequenzen aufweisen. 51
60 4 Ergebnisse Abbildung 4.18 Sekretion von TNF-α nach Elektroporation der PBMC mit HIV-1-spezifischer mrna. Vier Stunden nach Elektroporation wurde der Überstand der Zellen auf die Freisetzung von Zytokinen analysiert. Die Abbildungen A-D stellen repräsentative Beispiele dar. A: Patient 0284 B: Patient 0143 C: Patient 0347 D: Normalspender
61 4 Ergebnisse 4.2 Funktionelle Analysen mit HIV-1-spezifischen mrnas mrna-kodierte HIV-1-Proteine stimulieren die Generierung von HIV-1-spezifischen CTL Linien Standardmäßig werden HIV-1-spezifische CTL Linien in vitro generiert, indem PBMC von HIV-1-Patienten mit HIV-1-spezifischen Peptiden, die CTL-Epitope enthalten, inkubiert werden. Nun sollte getestet werden, ob auch durch die Elektroporation von PBMC mit HIV-1-spezifischer mrna CTL zur Proliferation induziert werden können. Daher wurden PBMC von HIV-1-Patienten mit HIV-1-spezifischer mrna elektroporiert. Nach vier Stunden wurden unbehandelte PBMC des Patienten mit den elektroporierten PBMC stimuliert. Die Zellen wurden zehn Tage kultiviert und anschließend wurde die CTL- Aktivität im γ-ifn ELISPOT analysiert. Dazu wurden 1x10 5 Zellen pro Well auf die ELISPOT-Platte gegeben und mit einem HIV-1-Peptid getestet. Wie exemplarisch in Abbildung 4.19 dargestellt, konnte eine Stimulation der Zellen durch die mrna-kodierten HIV-1-Proteine erreicht werden. SF2 gag mrna sowie autologe gag mrna des Patienten führten zu einer klaren Stimulation von HIV-1-spezifischen CTL Linien, wohingegen GFP mrna-elektroporierte PBMC keine CTL Linien induzieren konnten. Als Kontrolle wurden PBMC des Patienten mit dem Peptid Gag07-41 inkubiert. Abbildung 4.19 PBMC, welche mit gag mrna elektroporiert wurden, stimulierten HIV-1-spezifische CTL Linien. PBMC eines HIV-1-Patienten wurden mit mrna elektroporiert. Nach 4 Stunden wurden unbehandelte PBMC mit den mrna-elektroporierten PBMC stimuliert. Nach 10-tägiger Kultivierung wurden die Zellen im γ-ifn ELISPOT getestet. Als read-out wurde das Peptid Gag07-41 verwendet. GFP mrnaelektroporierte PBMC zeigten keine Stimulation von Gag-spezifischen CTL Linien. 53
62 4 Ergebnisse PBMC proliferieren nicht spezifisch nach Elektroporation mit mrna Nachdem festgestellt wurde, dass mrna-elektroporierte PBMC HIV-1-spezifische CTL Linien stimulieren können (siehe 4.2.1), sollte geklärt werden, ob die mrnaelektroporierten PBMC proliferieren. Dazu wurden autologe PBMC mit mrnaelektroporierten PBMC vier Stunden nach Transfektion stimuliert und anschließend mit CFSE (Carboxyfluoreszein-Succinimidyl Ester) markiert. Dabei diffundiert CFSE passiv in die Zellen und nach Abspaltung einer Acetatgruppe entsteht ein Fluoreszenzmolekül. Als Kontrolle wurden PBMC mit PHA oder einem HIV-1-Peptid stimuliert. Die Proliferation der Zellen wurde durch die Detektion der zellteilungsbedingten Abnahme der CFSE- Fluoreszenz über einen Verlauf von mehreren Tagen im Durchflusszytometer bestimmt. Wie in Abbildung 4.20 exemplarisch dargestellt, konnte nach einer Kultivierung von fünf Tagen eine CFSE-Abnahme festgestellt werden. Allerdings wurde diese CFSE-Abnahme nicht nur wie erwartet bei den nef mrna-elektroporierten PBMC detektiert, sondern auch in den unbehandelten PBMC und den mock-elektroporierten PBMC. Daher konnte man mit dieser Methode keine spezifische Induktion der Proliferation der mrnaelektroporierten PBMC erreichen. Abbildung 4.20 Analyse der Proliferation nach mrna-elektroporation von PBMC. PBMC eines HIV-1- infizierten Patienten wurden mock- bzw. mit HIV-1 nef mrna elektroporiert. Vier Stunden nach Elektroporation wurden die elektroporierten Zellen als Stimulationszellen für unbehandelte PBMC verwendet und mit CFSE markiert. Die Proliferation der Zellen wurde durch die proliferationsbedingte Abnahme der CFSE-Fluoreszenz am Durchflusszytometer gemessen. Dabei wurde die CFSE-Fluoreszenz am Tag 5 (grau) mit der am Tag 1 (schwarze Linie) verglichen. In allen drei Ansätzen konnte eine geringe Proliferation der Zellen detektiert werden, jedoch kein Unterschied zwischen den unbehandelten PBMC (links), den mockelektroporierten PBMC (Mitte) und den nef mrna-elektroporierten PBMC festgestellt werden. Dargestellt ist ein repräsentatives Experiment aus drei Versuchen. 54
63 4 Ergebnisse Mit HIV-1-spezifischer mrna-elektroporierte PBMC zeigen nach Kryokonservierung noch schwache Immunantworten Als nächstes sollte untersucht werden, ob kryokonservierte und wieder aufgetaute PBMC, die mit HIV-1-spezifischer mrna elektroporiert wurden, noch T-Zellreaktionen im γ-ifn ELISPOT auslösen können. Dies ist wichtig, wenn mrna-elektroporierte PBMC für das Immunmonitoring verwendet oder als Impfstoff wiederholt verabreicht werden sollen. Daher wurden PBMC von HIV-1-infizierten Patienten 24 Stunden nach Elektroporation nach Standardprotokoll (siehe Methoden) kryokonserviert. Bei der nächsten Routineuntersuchung des Patienten wurden frische PBMC aus Citratblut isoliert und diese als Effektorzellen zusammen mit den aufgetauten, mrna-elektroporierten PBMC im ELISPOT Assay inkubiert. Dabei zeigte sich, dass die kryokonservierten und wieder aufgetauten PBMC noch Immunantworten induzieren können, jedoch die Stärke der induzierten Reaktion deutlich schwächer ist als zum Zeitpunkt vor Kryokonservierung (Abbildung 4.21). Es wurden stärkere T-Zellreaktionen erzielt, wenn zu den aufgetauten, mrna-elektroporierten PBMC frische, autologe PBMC als Effektorzellen zugegeben wurden (Abbildung 4.21). Im Vergleich zu den ursprünglich ausgelösten Immunantworten vor Kryokonservierung lag die Stärke der Immunantworten nach der Kryokonservierung bei durchschnittlich 19% ohne Effektorzellen und bei 30% mit Effektoren. Insgesamt wurde der Versuch dreimal durchgeführt. In einem Fall konnte überhaupt keine Reaktion im ELISPOT quantifiziert werden. 55
64 4 Ergebnisse Abbildung 4.21 Kryokonservierte, aufgetaute PBMC, die mit HIV-1-spezifischer mrna elektroporiert wurden, konnten noch T-Zellantworten induzieren, die jedoch deutlich schwächer sind. PBMC von HIV-1-infizierten Patienten (hier exemplarisch 0225) wurden mit HIV-1-spezifischer mrna elektroporiert und vier Stunden nach Elektroporation im γ-ifn ELISPOT Assay inkubiert (schwarze Balken). Nach 24 Stunden wurden die Zellen kryokonserviert. Nach Auftauen wurden die kryokonservierten Zellen sogleich in einem ELISPOT Assay eingesetzt (graue Balken). Die induzierten T-Zellreaktionen der aufgetauten PBMC sind deutlich besser, wenn man frische, autologe PBMC des Patienten als Effektorzellen hinzufügt mrna-kodierte Nef-Proteine reduzieren die Expression von CD4, CXCR4 und HLA-I Nachdem eine schnelle, effiziente Expression der mrna-kodierten Proteine und eine gute Immunogenität der HIV-1-Proteine in vitro nachweisbar war, stellte sich die Frage, ob die durch mrna-elektroporation in PBMC eingebrachten HIV-1-Proteine funktionell aktiv sind. Für das HIV-1-Protein Nef sind viele Funktionen bekannt. Unter anderem reduziert Nef die Moleküle HLA-I, CD4 sowie CXCR4 auf der Oberfläche von infizierten Zellen ( ). Daher sollte nun untersucht werden, ob sich nach Elektroporation von PBMC mit nef mrnas die Expression dieser Moleküle verändert. Dazu wurden PBMC von Normalspendern und HIV-1-infizierten Patienten mit verschiedenen nef mrnas elektroporiert und nach 24 Stunden mit spezifischen Antikörpern für CD4, CXCR4 und HLA-A*02 gefärbt. Die Analyse der Fluoreszenzintensitäten erfolgte am 56
65 4 Ergebnisse Durchflusszytometer. Wie in Abbildung 4.22 zu sehen ist, konnte eine effiziente Reduktion für alle drei Moleküle nach Elektroporation mit nef mrna beobachtet werden. Während knapp 50% aller mock-elektroporierten PBMC CD4 positiv sind, sank die Anzahl der CD4 positiven Zellen nach nef mrna-elektroporation auf 36%. Dabei blieb nur eine kleine Population von Zellen zurück, die das CD4-Molekül in hohem Ausmaß exprimierten, die meisten Zellen zeigten nur geringe Mengen an CD4 (Abbildung 4.22 A). Für das HLA- A*02 Molekül konnte ein Rückgang der Expression bis zu 50% verzeichnet werden (Abbildung 4.22 B). Die CXCR4-Menge wurde durch nef mrna am wenigsten beeinflusst, dennoch wurde ein Unterschied von bis zu 25% CXCR4-positiver Zellen im Vergleich zu mock-elektroporierten Zellen nachgewiesen (Abbildung 4.22). Bei der Analyse von verschiedenen nef mrnas konnte festgestellt werden, dass sich die unterschiedlichen Nef-Proteine auch in ihrer Fähigkeit differenzieren, wie effektiv sie die genannten Moleküle regulieren (Daten nicht gezeigt). Folglich sind die generierten nef mrnas in der Lage die Expression von CD4, CXCR4 und HLA-I entscheidend zu modulieren und somit auch funktionell aktiv. Abbildung 4.22 Expression der Oberflächenmoleküle CD4, HLA-A*02 und CXCR4 wurde nach Elektroporation mit nef mrna reduziert. PBMC von Normalspendern wurden mit nef mrna bzw. mockelektroporiert, nach 24 Stunden mit spezifischen Antikörpern gefärbt und durchflusszytometrisch analysiert. A: Expression des Oberflächenmarkers CD4 in PBMC. In den mock-elektroporierten PBMC (links) waren knapp 50% CD4 +. Nach Elektroporation mit nef mrna sank die Anzahl CD4 + -Zellen auf 36% (rechts). B und C: Die Expression der Moleküle HLA-A*02 (B) und CXCR4 (C) auf der Oberfläche von PBMC verringerte sich nach nef mrna Elektroporation. 57
66 4 Ergebnisse Folgen der simultanen Elektroporation von gag und nef mrna Die simultane Elektroporation zweier HIV-1-spezifischer mrnas in B-Zelllinien hat keine Auswirkungen Im Hinblick auf eine Anwendung der mrna als therapeutische Vakzine ist wichtig zu untersuchen, ob die gleichzeitige Verabreichung von mehreren HIV-1-spezifischen mrnas die Wirkung der verschiedenen, einzelnen mrnas beeinflusst. Zum einen könnte die gleichzeitige Impfung mit nef und gag mrna die Expression eines der beiden Konstrukte verändern oder zum anderen könnte eines der Proteine zu einer schlechteren Induktion der γ-ifn Ausschüttung führen. Daher wurden EBV-transformierte B-Zelllinien einzeln mit gag bzw. nef mrna elektroporiert. In einem weiteren Ansatz wurde die B- Zelllinie simultan mit beiden mrna-konstrukten elektroporiert. Die Expression der beiden Proteine wurde vier Stunden nach Elektroporation in allen Ansätzen mittels Durchflusszytometrie gemessen und verglichen. Dabei ließ sich ermitteln, dass die gleichzeitige Elektroporation von beiden mrnas in einem gemeinsamen Ansatz zu einer geringen Reduktion der Expression führte, die jedoch maximal bei 25% lag. In den meisten Fällen war keine Veränderung oder nur eine schwache Minderung von 10% nach gemeinsamer Elektroporation zu beobachten (Abbildung 4.23). Somit gibt es im Hinblick auf die Effektivität der Transfektion nur geringe Einbußen, wenn zwei unterschiedliche HIV-1-spezifische mrnas gleichzeitig in einem Ansatz verabreicht werden. Die elektroporierten B-Zelllinien wurden zudem in einen γ-ifn ELISPOT Assay eingesetzt. Da B-Zellen zu den Antigen-präsentierenden Zellen gehören, wurden diese mit Gag- bzw. Nef-spezifischen CTL Linien inkubiert. Für die Generierung dieser CTL Linien wurden PBMC von HIV-1-infizierten Patienten, welche zudem Träger des HLA-Allels B*13 waren, mit dem Nef-Peptid Nef RI9 und mit dem Gag-Peptid p15 RI9 stimuliert, beides HLA- B*13-restringierte Epitope, und für zehn Tage in R10IL2 kultiviert. Für die Elektroporation wurde daher auch eine HLA-B*13 positive B-Zelllinie verwendet, damit die entsprechenden Epitope von der B-Zelle den CTL Linien präsentiert werden können. Die Ausschüttung von γ-ifn bleibt meist unverändert, unabhängig davon, ob die HIV-1- spezifische mrna einzeln oder zusammen mit einer weiteren HIV-1-spezifischen mrna verwendet wurde (Abbildung 4.24). Wurde eine geringe Reduktion der Gag- oder Nef- Expression in den Zellen detektiert, die mit beiden mrnas elektroporiert wurden, so wurde teilweise eine geringfügig schlechtere Erkennung der elektroporierten B-Zellen durch die CTL Linie beobachtet. Allerdings wird dies wahrscheinlich die Folge der 58
67 4 Ergebnisse geringeren Proteinexpression sein. Der Versuch wurde dreimal durchgeführt und in allen Versuchen konnte das Resultat bestätigt werden. Abbildung 4.23 B-Zelllinien, welche mit gag und nef mrna simultan elektroporiert wurden, zeigten im Gegensatz zur Einzelelektroporation der jeweiligen mrna nur eine geringe Reduktion der Proteinexpression. B-Zelllinien wurden 4 Stunden nach Elektroporation mit den HIV-1-spezifischen Antikörpern (p24 183/H12 bzw. Nef EH1) intrazellulär gefärbt und am Durchflusszytometer analysiert. Die Population der Gag- bzw. Nef-positiven Zellen befindet sich auf der y-achse. 46% der B-Zellen, die nur mit gag mrna elektroporiert wurden, exprimierten das Gag-Protein (Mitte links). Im Vergleich dazu waren es 34% Gag-positive Zellen in den B-Zellen, die sowohl mit gag als auch mit nef mrna elektroporiert wurden (unten links). Für Nef waren 53% der nef mrna-elektroporierten B-Zellen positiv (Mitte rechts), dagegen sind es in den B-Zellen, welche mit gag und nef mrna elektroporiert wurden, 38% (unten rechts). Als Kontrolle dienten mock-elektroporierte B-Zelllinien, die ebenfalls mit den HIV-1-spezifischen Antikörpern gefärbt wurden (oben). 59
68 4 Ergebnisse Abbildung 4.24 Die Erkennung von mrna-elektroporierten B-Zelllinien durch HIV-1-spezifische CTL Linien im γ-ifn ELISPOT Assay blieb konstant, unabhängig davon, ob die B-Zelllinie mit der mrna einzeln oder zusammen mit einer weiteren mrna elektroporiert wurde. 1x10 5 Zellen einer B-Zelllinie wurden vier Stunden nach Elektroporation zusammen mit 5x10 4 Zellen einer CTL Linie im γ-ifn ELISPOT Assay inkubiert. A: Eine Nef-spezifische CTL Linie erkannte die B-Zelllinie, welche mit nef mrna elektroporiert wurde, im gleichen Ausmaß wie die B-Zelllinie, die sowohl mit nef als auch mit gag mrna elektroporiert wurde. Ebenso wurden die Peptid-inkubierte B-Zelllinie sowie das Peptid erkannt. B: Eine Gagspezifische CTL Linie (Epitop p15 RI9) erkannte die B-Zelllinie, welche mit gag mrna elektroporiert wurde, gleichermaßen wie die B-Zelllinie, die mit gag und nef mrna zusammen elektroporiert wurde. Das Peptid alleine und die Peptid-inkubierte B-Zelllinie wurden von der CTL Linie auch erkannt. Alle weiteren Ansätze dienten als Negativ-Kontrolle und zeigten keine Erkennung durch die CTL Linien. E-BZ: elektroporierte B-Zelllinie. 60
69 4 Ergebnisse Die simultane Elektroporation zweier HIV-1-spezifischer mrnas in PBMC führt zu einem Verlust der Immunantworten Nachdem keine wesentlichen Auswirkungen der gemeinsamen Elektroporation von gag und nef mrna in B-Zelllinien festgestellt wurde, analysierte ich die Folgen der simultanen Verabreichung zweier mrnas in PBMC von HIV-1-infizierten Patienten. Dazu wurden PBMC ebenfalls mit gag und nef mrna einzeln und in einem weiteren Ansatz zusammen elektroporiert. Vier Stunden nach der Elektroporation wurden die Zellen im γ-ifn ELISPOT analysiert. In Abbildung 4.25 ist ein exemplarisches Ergebnis dargestellt. PBMC des Patienten 0284 zeigten hierbei eine deutliche T-Zellantwort auf sein autologes Gag- Protein mit 434 SFU/2x10 5 PBMC. Eine mäßige Nef-spezifische T-Zellantwort konnte ebenso detektiert werden (45 SFU/2x10 5 PBMC). Bei einer gemeinsamen Elektroporation von gag und nef mrna könnte man aufgrund der Addition der Gag- und Nef-spezifischen T-Zellantwort eine Erhöhung der Immunantwort erwarten. Allerdings sank die HIV-1- spezifische Immunantwort auf 29 SFU/2x10 5 PBMC (Abbildung 4.25) herab. Die Proteinexpression von Gag und Nef im gemeinsamen Elektroporationsansatz (79% Gagexprimierende Zellen; 80% Nef-exprimierende Zellen) war identisch zur Expression im Einzelansatz, so dass die geringere T-Zellantwort nicht die Folge einer schwächeren Proteinexpression ist. Nef-Proteine, welche von in vitro synthetisierten nef mrnas kodiert werden, sind in der Lage die Konzentration der HLA-Moleküle A und B auf der Oberfläche zu reduzieren (siehe 4.2.4). Daher kann man vermuten, dass hier ebenfalls dieser Effekt auftritt. Durch Reduktion der HLA-A- und HLA-B-Moleküle werden HLA-A- und HLA-Brestringierte Epitope nicht mehr präsentiert, was zur Abschwächung der allgemeinen Immunantwort führt. Dieser Effekt konnte unter Verwendung der autologen nef mrna des Patienten ebenfalls beobachtet werden. 61
70 4 Ergebnisse Abbildung 4.25 Die HIV-1-spezifische T-Zellantwort verringerte sich drastisch bei simultaner Elektroporation von gag und nef mrna in PBMC. PBMC eines HIV-1-infizierten Patienten (0284) wurden zum einen einzeln mit SF2 nef mrna oder der autologen gag mrna elektroporiert, zum anderen wurden die beiden mrnas vereinigt elektroporiert. Vier Stunden nach Elektroporation wurden die Zellen im ELISPOT eingesetzt. Der Patient zeigte eine mäßige T-Zellantwort auf SF2 Nef und eine sehr starke Reaktion auf das autologe Gag. PBMC, welche mit beiden mrnas elektroporiert wurden, demonstrierten eine drastische Reduktion der Immunantwort. Als Kontrollen dienten mock-elektroporierte und GFP mrna-elektroporierte PBMC, die keine γ-ifn-sekretion zeigten. Aufgrund der Beobachtung, dass durch die simultane Elektroporation von gag und nef mrna die HIV-1-spezifische Immunantwort in PBMC abnimmt und Nef die Expression der HLA-A- und HLA-B-Moleküle reduzieren kann (127), sollte genauer untersucht werden, ob T-Zellantworten, welche durch HLA-A- und HLA-B-Epitope entstehen, durch die Elektroporation mit nef mrna verringert werden. Dafür wurden PBMC eines HIV-1- infizierten Patienten (0566) mit gag bzw. nef mrna elektroporiert und diese elektroporierten PBMC im Anschluss in einen γ-ifn ELISPOT mit diversen HIV-1- Peptiden inkubiert. Dabei wurden Peptide gewählt, die Epitope zu den jeweiligen HLA-Aund HLA-B-Allelen des Patienten enthalten. Als Kontrolle dienten mock-elektroporierte PBMC. Die Inkubation der mock-elektroporierten PBMC mit den Peptiden führte zu einer deutlichen Sekretion von γ-ifn (Abbildung 4.26). PBMC, welche mit gag mrna elektroporiert wurden, zeigten ähnliche, wenn auch schwächere T-Zellantworten auf die Peptidstimulationen. Dagegen resultierte die Stimulation mit Peptiden in den nef mrnaelektroporierten PBMC in einer deutlich verringerten T-Zellantwort (Abbildung 4.26). Da die verwendeten Peptide allen Epitopen entsprechen, welche von HLA-A- bzw. HLA-B- 62
71 4 Ergebnisse Molekülen präsentiert werden, kann die verminderte Expression dieser Moleküle durch Nef Ursache der geringeren T-Zellantwort sein. Zusammenfassend offenbaren diese Ergebnisse, dass durch das mrna-kodierten Nef- Protein in PBMC die Expression von HLA-A- und HLA-B-Moleküle reduziert wird, so dass sich die Immunantworten verringern. Abbildung 4.26 PBMC eines HIV-1-infizierten Patienten (0566), welche mit nef mrna elektroporiert wurden, zeigten eine geringere T-Zellantwort nach Peptidstimulation als mock-elektroporierte PBMC oder gag mrna-elektroporierte PBMC. PBMC wurden mit gag mrna bzw. mit nef mrna elektroporiert und vier Stunden nach Elektroporation wurden 2x10 5 PBMC mit HIV-1-Peptiden inkubiert. Dabei führte die Stimulation mit Peptiden in den mock- und gag mrna-elektroporierten PBMC zu einer klaren Immunantwort, wohingegen die nef mrna-elektroporierten PBMC nur eine schwache T-Zellantwort auf die Peptidstimulation aufwiesen. Die Anzahl der SFU, welche durch die jeweilige Elektroporation verursacht wird, ist in dieser Darstellung bereits subtrahiert Mit mrna-elektroporierten PBMC kann die CTL-Epitop- Hierarchie studiert werden Da trotz einer vorhandenen CTL Antwort gegen eine Vielzahl von Epitopen viele Patienten AIDS entwickeln, gibt es anscheinend wichtige Unterschiede in der Qualität der Epitope. Manche CTL-Epitope werden viel besser von HIV-1-spezifischen CTLs erkannt, da sie anscheinend besser prozessiert und präsentiert werden. Daher stellte sich die Frage, ob es eine gewisse CTL-Epitop-Hierarchie gibt und welche Epitope besonders gut sind. Dies sollte mit der entwickelten Methode der mrna-elektroporation getestet werden. Dazu wurden PBMC eines HIV-1-infizierten Patienten mit gag oder nef mrna elektroporiert und 63
72 4 Ergebnisse anschließend zusammen mit autologen Gag- bzw. Nef-spezifischen CTL Linien in einen γ-ifn ELISPOT verwendet. Es wurden 1x10 5 Zellen der CTL Linie zusammen mit 2x10 5 elektroporierten PBMC pro Well inkubiert. Dabei konnte festgestellt werden, dass die Erkennung der Gag-Epitope, welche von der gag mrna prozessiert und präsentiert werden, durch die verschiedenen Gag-spezifischen CTL Linien sehr variiert. Das HLA- A*02-restringierte SL9v8 Epitop (27), welches in der autologen gag mrna enthalten ist, wurde im Vergleich zur Peptiderkennung zu 47% von der CTL Linie erkannt (Abbildung 4.27 A). Im Gegensatz dazu wurde das HLA-A*03-restringierte p17 KK9 Epitop (128), welches in der autologen gag mrna enthalten ist, zu 75% von der spezifischen CTL Linie erkannt (Abbildung 4.27 B). Somit kann man konstatieren, dass das p17 KK9 Epitop in Gag besser prozessiert und präsentiert wird als das SL9 Epitop. Bezüglich des Nef- Proteins konnten keine großen Unterschiede in der Erkennung der verschiedenen, untersuchten Epitope festgestellt werden. Wie in Abbildung 4.27 ersichtlich wird, war die Erkennung von Nef-Epitopen, welche durch nef mrna-elektroporierte PBMC präsentiert werden, sehr gering und lag zwischen 10% und 20%. Dennoch konnte eine leicht bessere Präsentation des B*18-Epitops RY10 (129) mit 17% im Vergleich zu dem B*07-Epitop Nef7v1 (130) mit 10% eruiert werden (Vergleiche Abbildung 4.27 A und B). Es konnten zudem nicht viele Nef CTL-Epitope miteinander verglichen werden, da die Generierung von Nef-spezifischen CTL Linien, die nicht mit anderen Epitopen kreuzreagierten, sehr schwierig war. In einem anderen Versuchsansatz (siehe ) wurde deutlich, dass das HLA-B*13- restringierte Nef RI9 Epitop ein gut prozessiertes Nef-Epitop ist. PBMC von dem HIV-1- infizierten Patienten 0609 wurden mit heterologer nef mrna elektroporiert und anschließend im γ-ifn ELISPOT Assay analysiert. Im Vergleich zur Peptiderkennung wird das Nef RI9 Epitop, welches in der nef mrna enthalten ist, zu 63% erkannt, wenn PBMC im Assay eingesetzt wurden. Bei PBMC ist der Effekt geringer (Abbildung 4.16). Frühere Versuche der Arbeitsgruppe zeigten, dass sich die Nefspezifische CTL Antwort des Patienten dominant gegen das Peptid Nef RI9 richtet und andere Peptide nicht erkannt wurden. Damit kann die Immunantwort der PBMC gegen die heterologe nef mrna allein dem Epitop RI9 zugeschrieben werden (Abbildung 4.16). Die gute Prozessierung des Nef-Epitops RI9 könnte auch die gute Kontrolle der Virusreplikation von HLA-B*13-positiven Individuen erklären. Die Daten zeigen, dass die entwickelte Methode gut verwendet werden kann, um die Hierarchie der CTL-Epitope zu studieren. Einige CTL-Epitope werden deutlich besser von den mrna-elektroporierten PBMC präsentiert und wären daher geeignete Kandidaten bei einer Vakzinierung. 64
73 4 Ergebnisse Abbildung 4.27 Das p17 KK9 Epitop in Gag wird von gag mrna-elektroporierten Zellen deutlich besser präsentiert als das SL9 Epitop in Gag, das RY10 Epitop in Nef wird von nef mrnaelektroporierten Zellen leicht besser präsentiert als das Nef7v1 Epitop. PBMC eines HIV-1-infizierten Patienten wurden mit dem p17 KK9 Peptid, einem bekanntem A*03-restringierten Peptid, stimuliert oder mit dem Peptid SL9v8, der autologen Variante des A*02-Epitops SL9, um CTL Linien zu generieren. PBMC des Patienten wurden mit gag mrna elektroporiert und vier Stunden nach Elektroporation mit autologen Gagspezifischen CTL Linien in einem γ-ifn ELISPOT Assay inkubiert. A: Die relative γ-ifn Sekretion der SL9v8- spezifischen CTL Linie. Im Vergleich zur Peptiderkennung lag die Erkennung des Epitops, präsentiert von den gag mrna-elektroporierten PBMC, bei 47%. B: Die relative γ-ifn Sekretion der p17 KK9-spezifischen CTL Linie. Dabei betrug die Epitoperkennung, präsentiert durch die gag mrna-elektroporierten PBMC, 75% im Vergleich zur Peptiderkennung der CTL Linie. Für Nef wurden ebenfalls PBMC eines HIV-1-infizierten Patienten mit dem HLA-B*18-restringierten Peptid RY10 und dem HLA-B*07-restringierten Peptid Nef7v1 stimuliert. Die daraus resultierenden CTL Linien wurde dann mit autologer nef mrna-elektroporierten PBMC des Patienten im ELISPOT inkubiert. C: Die relative Sekretion von γ-ifn der Nef7v1-spezifischen CTL Linie. Im Vergleich zur Peptiderkennung war die Erkennung des Epitops, präsentiert von den nef mrna-elektroporierten PBMC, sehr gering und lag bei 10%. D: Die relative γ-ifn Sekretion der RY10-spezifischen CTL Linie zeigte im Vergleich zur Peptiderkennung eine Erkennung des Epitops von 17%, wenn es von den nef mrna-elektroporierten PBMC präsentiert wurde. 65
74 4 Ergebnisse 4.3 Kreuzreaktion HIV-1-spezifischer CTLs mit humanen Proteinen Das Nef-Epitop TM9 ähnelt einem Epitop im humanen Protein Fat Breite CTL-Antwort gegen die Epitope Nef TM9 und Fat3 TM9 T/P Das CTL-Epitop Nef TM9 TPQVPLRPM ist über die HLA-I-Allele B*07 und B*35 restringiert (131, 132). Durch BLAST entdeckten wir eine Sequenzähnlichkeit des Epitops zu dem humanen Protein Fat3 (NCBI-Zugangsnummer: NM_ ). Die betreffende Sequenz des Fat3 unterscheidet sich dabei nur in zwei Aminosäuren und lautet PPQVPVRPM (TM9 T/P). Aufgrund dieser Sequenzähnlichkeit sollte getestet werden, ob HIV-1-infizierte Patienten CTLs gegen das HIV-1-Epitop TM9 sowie gegen das Fat3- Epitop TM9 T/P besitzen und ob diese kreuzreaktiv zueinander sind. Daher wurden PBMC von HIV-1-Patienten, die für HLA-B*07 oder HLA-B*35 positiv sind, mit den Peptiden TM9 und TM9 T/P in vitro stimuliert, um spezifische CTL Linien zu generieren. Das Auswachsen von Antigen-spezifischen CTL Linien wurde im γ-ifn ELISPOT analysiert. Insgesamt wurden 100 Patienten in 214 Tests untersucht. Das Ergebnis der ELISPOT-Screening Assays ist in Tabelle 4.3 zusammengefasst. Eine TM9-spezifische CTL Antwort konnte in 53 Patienten (53%) detektiert werden, eine TM9 T/P-spezifische Antwort in 43 Patienten (43%). Eine starke T-Zellantwort mit mehr als 250 SFU/1x10 5 Zellen konnte in 26 der 53 TM9-spezifischen CTL Linien beobachtet werden (26%) und 39 TM9-spezifische CTL Linien (39%) zeigten eine deutliche Kreuzreaktivität zu dem TM9 T/P Peptid. Von den 43 TM9 T/P-spezifischen CTL Linien wiesen 19 eine sehr starke T-Zellantwort (19%) auf und 31 der 43 TM9 T/P-spezifischen CTL Linien erkannten das Epitop Nef TM9. Für die Mehrzahl der Patienten konnte eine Immunreaktion gegen beide Peptide quantifiziert werden, wohingegen 11 Patienten eine spezifische Immunantwort nur gegen eins der Peptide zeigte. Zusätzlich habe ich die Daten auch hinsichtlich ihres HLA- B-Allels untersucht und gegliedert (Tabelle 4.4 A und B). Dabei konnte kein statistisch signifikanter Unterschied zwischen HLA-B*07- oder HLA-B*35-positiven Individuen bezüglich der Erkennung der Peptide TM9 und TM9 T/P festgestellt werden (Chi-Quadrat- Vierfeldertest). In Abbildung 4.28 sind exemplarische TM9- und TM9 T/P-spezifische CTL Linien dargestellt. Neben Kreuzreaktionen wurden einige CTL Linien auch in Titrationsanalysen eingesetzt. Dabei wurden keine kongruenten Ergebnisse erzielt. Einige CTL Linien erkannten das Peptid noch bei einer Konzentration von 10 ng/ml, andere CTL 66
75 4 Ergebnisse Linien wiederum konnten das Peptid bereits bei einer Konzentration von 10 µg/ml nicht mehr erkennen (Daten nicht gezeigt). Abbildung 4.28 Exemplarische γ-ifn ELISPOT Assays mit TM9- bzw. TM9 T/P-spezifischen CTL Linien. PBMC von HIV-1-infizierten Patienten wurden mit den Peptiden Nef TM9 (TPQVPLRPM) und TM9 T/P (PPQVPVPM), einem sequenzähnlichen Epitop im humanen Protein Fat3, stimuliert und im γ-ifn ELISPOT analysiert. TM9-spezifische CTL Linien sind in schwarz dargestellt, TM9 T/P-spezifische CTL Linien in Weiß. A: TM9-spezifische CTL Linie mit einer starken Kreuzreaktivität zu TM9 T/P. B: TM9-spezifische CTL Linie ohne Kreuzreaktivität zu TM9 T/P. C: TM9 T/P-spezifische CTL Linie, die ebenso das Peptid Nef TM9 erkennt. D: TM9 T/P-spezifische CTL Linie, die das Nef TM9 Peptid nicht erkennt. Zusammenfassend konnte festgestellt werden, dass das HIV-1 Nef-Epitop TM9 und das sequenzähnliche Epitop TM9 T/P im Fat3-Protein relativ häufig von HLA-B*07 und HLA- B*35-positiven HIV-1-Patienten erkannt wurden. In einigen Fällen wurde sogar eine stärkere CTL-Antwort gegen das Fat3-Epitop als gegen das HIV-1 Nef Epitop beobachtet. 67
76 4 Ergebnisse Tabelle 4.3 Übersicht der γ-ifn ELISPOT Screeningergebnisse. Insgesamt wurden 100 HIV-1-infizierte Patienten, die positiv für das HLA-B*07 oder/und HLA-B*35-Allel sind, auf eine Erkennung der beiden Peptide Nef TM9 und Fat3 TM9 T/P getestet. Eine starke Erkennung definierte sich durch eine Anzahl von mehr als 250 SFU/1x10 5 Zellen. Anzahl Patienten Häufigkeit [%] Erkennung von TM % Starke Erkennung von TM % Kreuzreaktion zu TM9 T/P 39 39% Erkennung von TM9 T/P 43 43% Starke Erkennung von TM9 T/P 19 19% Kreuzreaktion zu TM % Tabelle 4.4 Übersicht der γ-ifn ELISPOT Screeningergebnisse eingeteilt nach dem entsprechenden HLA-B-Allel. Insgesamt waren von 100 Patienten 57 HLA-B*07-positiv und 48 HLA-B*35-positiv. Davon trugen 5 Patienten beide Allele, HLA-B*07 und HLA-B*35. Es wurde kein statistisch signifikanter Unterschied in der Erkennung der beiden Peptide zwischen den zwei HLA-B-Allelen festgestellt. Eine starke Erkennung definierte sich durch eine Anzahl von mehr als 250 SFU/1x10 5 Zellen. A: Ergebnisse für das Peptid Nef TM9. B: Ergebnisse für das Peptid Fat3 TM9 T/P. A B Erkennung von TM9 Starke Erkennung von TM9 Kreuzreaktion zu TM9 T/P Erkennung von TM9 T/P Starke Erkennung von TM9 T/P Kreuzreaktion zu TM9 Anzahl Patienten HLA-B*07 Häufigkeit [%] HLA-B*07 Anzahl Patienten HLA-B*35 Häufigkeit [%] HLA-B* ,14% 23 47,92% 20 35,09% 7 14,58% 25 43,86% 16 33,33% Anzahl Patienten HLA-B*07 Häufigkeit [%] HLA-B*07 Anzahl Patienten HLA-B*35 Häufigkeit [%] HLA-B* ,61% 20 41,67% 10 17,54% 9 18,75% 22 38,60% 15 31,.25% pfat3 mrna Elektroporation führte trotz effizienter Proteinexpression zu keiner Erkennung durch Nef-spezifische CTLs Ob TM9- bzw. TM9 T/P-spezifische CTL Linien das Epitop TM9 T/P nicht nur als Peptid sondern auch als prozessiertes Peptid aus dem Protein erkennen, sollte in einer weiteren Analyse untersucht werden. Dazu stand ein bereits im Labor kloniertes Plasmid zur Verfügung, welches 609 Basen des Fat3-Gens als offenen Leserahmen mit einem N-terminalen Flag-tag enthält. Aufgrund der Größe des Fat3-Gens wurde nur ein kleiner Bereich des Gens kloniert, welcher auch das homologe Epitop beinhaltet. Das Plasmid beruht auf dem Vektor pgem4z64a und konnte somit zur mrna-synthese verwendet 68
77 4 Ergebnisse werden. Die in vitro transkripierte mrna pfat3 (partielles fat3) wurde anschließend zur Elektroporation von EBV-transformierten lymphoiden B-Zelllinien eingesetzt. Zudem wurden die Zelllinien auch mit GFP mrna, HIV-1 nef mrna (autologes Nef von 0545) sowie mock elektroporiert. Daraufhin wurde die Proteinexpression durch intrazelluläre Färbung mit einem Nef- bzw. Flag-tag-spezifischen Antikörper und anschließender Analyse am Durchflusszytometer überprüft. Die Elektroporation der B-Zelllinien war in allen vier durchgeführten Versuchen sehr effizient und zeigte folgende Mediane der Proteinexpression: 91% GFP-positive Zellen (Range 78%-92%), 59% der Zellen exprimierten das partielle Fat3-Protein (Range: 22%-78%) und 75% der Zellen exprimierten das HIV-1 Nef-Protein (Range: 30-88%) (Abbildung 4.29). Abbildung 4.29 Sowohl das Nef-Protein als auch das partielle Fat3-Protein wurden effizient nach mrna-elektroporation in B-Zelllinien exprimiert. B-Zelllinien wurden mock, mit HIV-1 nef mrna oder pfat3 mrna elektroporiert. Vier Stunden nach Elektroporation wurden die Zellen permeabilisiert, intrazellulär mit einem Nef- bzw. einem Flag-tag-spezifischen Antikörper gefärbt und im Durchflusszytometer analysiert. Dargestellt ist eine exemplarische Färbung aus vier Versuchen. In der oberen Reihe ist die Färbung mit dem Flag-tag-spezifischen Antikörper ersichtlich, in der unteren Reihe die Färbung mit dem Nef-spezifischen Antikörper. Der linke Dot Plot zeigt jeweils die mock-elektroporierten B-Zellen. Es konnte eine Expression des pfat3 mrna-kodierten Proteins von 78% detektiert werden. Knapp 88% der nef mrna-elektroporierten B-Zellen exprimierten das HIV-1 Nef-Protein. Die elektroporierten B-Zelllinien wurden mit den TM9- bzw. TM9 T/P-spezifischen CTL Linien im γ-ifn ELISPOT Assay ko-inkubiert, um die spezifische Erkennung der von den 69
78 4 Ergebnisse mrna-kodierten Proteinen durch die CTL Linien zu analysieren. Abbildung 4.30 repräsentiert ein exemplarisches Ergebnis eines ELISPOT Assays mit CTL Linien eines HLA-B*07-positiven Patienten (A) und mit CTL Linien eines HLA-B*35-positiven Patienten (B). Das Experiment wurde je dreimal durchgeführt, auch mit CTL Linien von anderen Patienten. In allen Versuchen konnte man eine deutliche Erkennung des Nef TM9- Epitops, welches durch die nef mrna-elektroporierten B-Zellen prozessiert und präsentiert wurde, durch die TM9- und TM9 T/P-spezifische CTL Linie beobachten. Die Ko-inkubation der TM9- und TM9 T/P-spezifischen CTL Linie mit B-Zellen, welche mit pfat3 mrna elektroporiert wurden, induzierten nur eine geringe Freisetzung von γ-ifn. Diese war vergleichbar mit der Erkennung von mock-elektroporierten und GFP mrnaelektroporierten B-Zellen durch die CTL Linien und stellten somit Hintergrundwerte dar. Eine starke Sekretion von γ-ifn wurde stets durch die Inkubation der CTL Linien mit Peptiden und mit Peptid-inkubierten B-Zelllinien erreicht (Abbildung 4.30). Folglich wird das Epitop Nef TM9 sowohl als exogenes Peptid als auch als Peptid, welches endogen von den nef mrna-elektroporierten B-Zellen prozessiert und präsentiert wird, von CTL Linien erkannt. Dagegen wird das Fat3 Epitop TM9 T/P gut als exogenes Peptid von TM9 T/P-spezifischen und TM9-kreuzreaktiven CTL Linien erkannt, jedoch wurden B-Zellen, die mit pfat3 mrna elektroporiert wurden und das Protein exprimierten, von diesen CTL Linien nicht erkannt. 70
79 4 Ergebnisse Abbildung 4.30 TM9- und TM9 T/P-spezifische CTL Linien erkennen nef mrna-elektroporierte B- Zelllinien, aber nicht pfat3 mrna-elektroporierte B-Zelllinien. B-Zellen (B-LCL) wurden mock, mit GFP mrna, HIV-1 nef mrna oder mit pfat3 mrna elektroporiert. Vier Stunden nach Elektroporation wurden die elektroporierten B-LCL mit TM9- und TM9 T/P-spezifischen CTL Linien in einem γ-ifn ELISPOT Assay koinkubiert. Es wurden jeweils 5x10 4 Zellen der CTL Linie und 1x10 5 Zellen der elektroporierten B-LCL eingesetzt. Auf der y-achse sind die SFU dargestellt. A: Exemplarisches Ergebnis von CTL Linien, die HLA- B*07-positiv sind. B: Exemplarisches Ergebnis von CTL Linien, die HLA-B*35-positiv sind. Es wurden jeweils drei Versuche durchgeführt und in keinem konnte eine Erkennung der pfat3 mrna-elektroporierten B-LCL beobachtet werden In silico Analysen des fat3 mrna-konstrukts sagen eine Prozessierung des Epitops TM9 T/P durch das Proteasom mit einer 50%igen Wahrscheinlichkeit voraus Obwohl eine klare Erkennung des Peptides TM9 T/P durch TM9 T/P-spezifische und TM9-kreuzreaktive CTL Linien detektiert werden konnte, beobachtete ich trotz effizienter Proteinexpression keine Erkennung des pfat3-proteins, welches durch die mrna kodiert wird. Eine fehlende oder veränderte Prozessierung des Konstrukts zu dem entsprechenden Epitop durch das Proteasom könnte eine mögliche Erklärung sein. Aus diesem Grund untersuchte ich die 609 Basen lange Sequenz des fat3-gens, welche in unserem Plasmid enthalten ist, in silico auf Proteasom-Spaltung. Dazu wurde das 71
80 4 Ergebnisse Programm MAPPP verwendet (siehe Tabelle 6.21). Dieses Programm beruht auf FRAGPREDICT (133, 134) und prognostiziert eine Prozessierung des Epitops TM9 T/P durch das Proteasom (Abbildung 4.31). Allerdings ist die vorhergesagte Wahrscheinlichkeit mittelmäßig. Das Nef-Epitop TM9 wird mit einer 99%igen Wahrscheinlichkeit prozessiert, das TM9 T/P Epitop hingegen nur mit einer Erwartung von 50-59%. Es ist daher zu vermuten, dass das Epitop TM9 T/P aus dem pfat3-konstrukt nicht durch das Proteasom gespalten wird und es daher auch trotz einer nachweisbaren Proteinexpression nach pfat3 mrna-elektroporation in B-Zelllinien zu keiner Erkennung durch CTL Linien kommt. Abbildung 4.31 In silico Analyse auf Proteasomschnittstellen der Proteinsequenzen von HIV-1 Nef und Fat3 offenbarte, dass die Wahrscheinlichkeit einer Prozessierung für das Epitop TM9 bei 99% liegt, für das Epitop TM9 T/P dagegen nur bei maximal 59%. Die Sequenzen der Proteine, die von der mrna kodiert werden, wurden mit Hilfe des Programms MAPPP analysiert. Dargestellt ist nur der betreffende Ausschnitt aus der Proteinsequenz. Orange hervorgehobene Sequenzen markieren die besagten Epitope. Die Zahlen in Klammern geben die Wahrscheinlichkeit an, mit welcher das entsprechende Peptid prozessiert wird. Je höher der Wert bei 1 liegt, desto wahrscheinlicher wird das Epitop prozessiert Das Nef-Epitop Nef7v1 ist einem Epitop im humanen Enzym D-Aminosäuren-Oxidase (DAO) ähnlich Ein weiteres Nef CTL-Epitop, welches eine hohe Sequenzähnlichkeit zu einem Epitop eines humanen Proteins besitzt, ist das Epitop Nef7v1 mit der Sequenz FPVRPQVPL. Es ist über die HLA-I-Allele B*07, B*35 und B*51 restringiert (135, 136) und unterscheidet sich in nur drei Aminosäuren von einem Epitop des humanen Flavoproteins D- Aminosäuren-Oxidase (DAO) (NCBI-Zugangsnummer: NM_001917) mit der Sequenz RPVRPQIRL (Nef7v4). Es sollte nun getestet werden, ob HIV-1-infizierte Patienten 72
81 4 Ergebnisse spezifische CTLs aufweisen, die das humane Epitop aus DAO erkennen. Daher wurden PBMC von HIV-1-Patienten mit den entsprechenden HLA-B-Allelen mit den Peptiden Nef7v1 und Nef7v4 in vitro stimuliert und die Generierung von Antigen-spezifischen CTL Linien wurde nach zehntägiger Kultivierung im γ-ifn ELISPOT Assay überprüft. In 149 Tests wurden 104 Patienten untersucht. Von diesen 104 Patienten wiesen 37 Patienten Nef7v1-spezifische CTLs auf (36%), wobei 11 CTL Linien eine sehr starke Reaktion mit über 250 SFU/1x10 5 Zellen zeigten (11%) und 14 CTL Linien das humane Epitop Nef7v4 kreuzreaktiv erkennen konnten (13%). Die Stimulation von PBMC mit dem Peptid Nef7v4 führte in 26 Patienten zu einer Erkennung (25%). Dabei zeigten sieben CTL Linien (7%), die spezifisch sind für das DAO-Epitop, eine starke Reaktion mit über 250 SFU/1x10 5 Zellen und 19 wiesen eine Kreuzreaktion zu dem HIV-1 Nef-Epitop Nef7v1 auf (18%). Diese Ergebnisse sind in Tabelle 4.5 zusammengefasst. Tabelle 4.5 Übersicht der γ-ifn ELISPOT Screeningergebnisse. Insgesamt wurden 104 HIV-1-infizierte Patienten, die positiv für das HLA-Allel-B*07, -B*35 und/oder B*51 sind, auf eine Erkennung der beiden Peptide Nef7v1 und DAO Nef7v4 getestet. Eine starke Erkennung definierte sich durch eine Anzahl von mehr als 250 SFU/1x10 5 Zellen. Anzahl Patienten Häufigkeit [%] Erkennung von Nef7v ,58% Starke Erkennung von Nef7v ,58% Kreuzreaktion zu Nef7v4 (DAO) 14 13,46% Erkennung von Nef7v4 (DAO) 26 25,00% Starke Erkennung von Nef7v4 7 6,73% Kreuzreaktion zu Nef7v ,27% Des Weiteren wurden die Daten nach dem jeweiligen HLA-B-Allel gegliedert. Unter den 104 Patienten waren 45 Patienten für HLA-B*07 positiv, 39 für HLA-B*35 und 25 für HLA- B*51. Jedoch konnte zwischen den verschiedenen HLA-B-Allelen kein signifikanter Unterschied in der Erkennung der Epitope detektiert werden (Chi-Quadrat-Vierfeldertest). Die Ergebnisse sind in Tabelle 4.6 dargestellt. Insgesamt konnten in HIV-1-infizierten Patienten spezifische CTLs gegen das DAO- Epitop, welches dem HIV-1-Epitop Nef7v1 stark ähnelt, nachgewiesen werden. Allerdings war das Ausmaß sehr gering, denn in nur 25% der Fälle ließ sich eine CTL-Antwort gegen das humane Epitop identifizieren und nur 7% zeigten dabei eine deutliche Reaktion. Aufgrund der geringen Frequenz starker CTL-Reaktionen gegen das humane Nef7v4 Epitop wurden keine weiteren Analysen durchgeführt. 73
82 4 Ergebnisse Tabelle 4.6 Übersicht der γ-ifn ELISPOT Screeningergebnisse für die Peptide Nef7v1 und DAO Nef7v4 eingeteilt nach dem entsprechenden HLA-B-Allel. Insgesamt waren von 104 Patienten 45 HLA-B*07- positiv, 39 HLA-B*35-positiv und 25 HLA-B*51-positiv. Es wurde kein statistisch signifikanter Unterschied in der Erkennung der beiden Peptide zwischen den HLA-B-Allelen festgestellt. Eine starke Erkennung definierte sich durch eine Anzahl von mehr als 250 SFU/1x10 5 Zellen. A: Ergebnisse für das Peptid Nef7v1. B: Ergebnisse für das Peptid Nef7v4. A B Anzahl Patienten HLA-B*07 Anzahl Patienten HLA-B*35 Anzahl Patienten HLA-B*51 Erkennung von Nef7v Starke Erkennung von Nef7v Kreuzreaktion zu Nef7v Anzahl Patienten HLA-B*07 Anzahl Patienten HLA-B*35 Anzahl Patienten HLA-B*51 Erkennung von Nef7v4 (DAO) Starke Erkennung von Nef7v Kreuzreaktion zu Nef7v
83 4 Ergebnisse 4.4 Analyse und Screening von neuen und bekannten CTL-Epitopen Bei einer therapeutischen Vakzine, die CTLs induzieren soll, spielen die verwendeten CTL-Epitope eine entscheidende Rolle. Eine effiziente Vakzine sollte CTL-Epitope enthalten, die starke Immunantworten bei möglichst vielen Patienten hervorrufen. Daher war es ein weiteres Ziel dieser Arbeit neue CTL-Epitope zu definieren, wobei der Fokus auf der Analyse von CTL-Epitopen in Patienten mit guter Kontrolle der Virusreplikation lag. Zum anderen sollte die Immunantwort, welche durch bekannte CTL-Epitope ausgelöst wird, in einer möglichst großen Kohorte an HIV-1-Infizierten untersucht werden. Hierbei rückten insbesondere CTL-Epitope ins Interesse, welche über das HLA-C-Molekül präsentiert werden, da diese durch das Nef-Protein nicht negativ beeinflusst werden Definition von drei neuen Gag und zwei neuen Nef CTL- Epitopen Zwei HIV-1-infizierte Patienten, die noch therapie-naiv sind und bisher eine gute Kontrolle der Virusreplikation aufzeigten, wurden in diese Untersuchung eingeschlossen. Nach Isolation aus Citratblut wurden die PBMC direkt in einen γ-ifn ELISPOT Assay mit Peptidsets, die einen Großteil der HIV-1-Proteine abdecken, inkubiert. Bei den Peptiden handelte es sich stets um Peptide, deren Aminosäurensequenz der Consensus B Sequenz ( von HIV-1 entsprechen. Die Peptide wurden vom NIH (National Institute of Health) bezogen. Anhand des HLA-Typs der Patienten und der Epitop-Datenbank (ELF=epitope location finder, HIV sequence database) konnten die Immunantworten, welche durch die verwendeten Peptide ausgelöst wurden, bestimmten CTL-Epitopen zugeordnet werden. Patient 0284 Für Patient 0284 wurden die Immunantworten gegen HIV-1 Gag, Nef, Tat, Rev, Vif, Vpr, Vpu und gegen die Hälfte von Pol mit Hilfe der Peptidsets analysiert. Die Untersuchung der T-Zellantworten gegen die akzessorischen Moleküle Vif, Vpr, Vpu, Tat und Rev ergab keine Reaktion. Somit wies dieser Patient keine CTL-Antwort gegen diese Proteine auf oder die Reaktion war zu gering, um sie ohne Vorstimulation der Zellen im γ-ifn ELISPOT Assay detektieren zu können. Auch bei der Analyse mit Pol-Peptiden konnte nur eine geringe T-Zellantwort gegen ein einziges, bekanntes Peptid ermittelt werden (Pol TYQIYQEPFKNLKTG, HLA-A*11, 21 SFU/1x10 5 PBMC). Im Gegensatz dazu konnten gegen Peptide der Proteine Gag und Nef deutliche T-Zellantworten erfasst werden. So besitzt Patient 0284 CTLs gegen sieben verschiedene Gag-Epitope und ein 75
84 4 Ergebnisse dominantes Nef-Epitop (Tabelle 4.7). Für das Peptid Gag war zum damaligen Zeitpunkt noch kein CTL-Epitop für den HLA-Typ des Patienten in der Datenbank (ELF) beschrieben. Tabelle 4.7 Übersicht über die vorhandenen CTL-Antworten des Patienten 0284 nach Inkubation der PBMC mit Gag- und Nef-Peptidsets. Die CTL-Antwort wurde im γ-ifn ELISPOT gemessen. Angegeben sind Name und Sequenz der verwendeten Peptide, ihre ausgelöste Immunantwort im ELISPOT (SFU) sowie darin enthaltene Epitope und das dazugehörige HLA-Allel. Die Epitope wurden mit dem Programm ELF (HIV sequence database) vorhergesagt. Peptidname Peptidsequenz SFU/ 1x10 5 PBMC Epitop HLA-Allel Gag07-20 SLYNTVATLYCVHQR 9 SLYNTVATL A*02 Gag07-36 QMVHQAISPRTLNAW 26 ISPRTLNAW B*57 QAISPRTL Cw*07 Gag07-41 EKAFSPEVIPMFSAL 75 KAFSPEVIPMF B*57 Gag07-77 RAEQASQEVKNWMTE 39 QASQEVKNW B*57 Gag07-87 EMMTACQGVGGPGHK 51 EMMTACQGV A*02 ACQGVGGPGHK A*11 Gag FLGKIWPSHKGRPGN 23 FLGKIWPSHK A*02 Gag LQSRPEPTAPPEESF 36?? Nef07-26 IYSQKRQDILDLWVY 49 RQDILDLWVY Cw*07 HLA-I 0284: A*02, A*11, B*39, B*57, Cw*06, Cw*07 Patient 0925 Für Patient 0925 wurden die Immunantworten gegen die HIV-1-Proteine Gag und Nef analysiert. Die Inkubation der PBMC mit den Nef-Peptiden offenbarte T-Zellreaktionen gegen insgesamt vier Epitope, wovon drei kein bekanntes CTL-Epitop zum HLA-Typ des Patienten enthielten, jedoch potentielle Epitope von dem Programm ELF vorgeschlagen wurden (Tabelle 4.8). Für Gag wurde eine Immunreaktion gegen drei Epitope beobachtet, wobei wiederum zwei Peptide noch unbekannte CTL-Epitope innehaben. 76
85 4 Ergebnisse Tabelle 4.8 Übersicht über die vorhandenen CTL-Antworten des Patienten 0925 nach Inkubation der PBMC mit Gag- und Nef-Peptidsets. Die CTL-Antwort wurde im γ-ifn ELISPOT gemessen. Angegeben sind Name und Sequenz der verwendeten Peptide, ihre ausgelöste Immunantwort im ELISPOT (SFU) sowie darin enthaltene Epitope und das dazugehörige HLA-Allel. Potentielle, aber noch nicht beschriebene CTL- Epitope sind kursiv dargestellt. Die Epitope wurden mit dem Programm ELF HIV (sequence database) vorhergesagt. Peptidname Peptidsequenz SFU/ 2x10 5 PBMC Epitop HLA-Allel Gag07-02 ASVLSGGELDRWEKI 11 VLSGGELDR GELDRWEKI A*66 B*40 Gag07-11 LERFAVNPGLLETSE 73?? Gag07-75 VDRFYKTLRAEQASQ 15?? Nef07-08 GVGAVSRDLEKHGAI 20 GAVSRDLEK A*66 Nef07-10 LEKHGAITSSNTAAN 26 LEKHGAITS B*40 Nef07-20 RPMTYKAAVDLSHFL 30 AAVDLSHFL Cw*16 Nef07-23 HFLKEKGGLEGLIYS 24 KEKGGLEGL B*44 HLA-I 0925: A*01, A*66, B*41, B*44, Cw*16, Cw* Analyse der neu definierten CTL-Epitope Das Peptid Gag enthält ein HLA-A*02-restringiertes CTL-Epitop Durch Inkubation der PBMC von Patient 0284 konnte eine T-Zellantwort für das Peptid Gag beobachtet werden (4.4.1). Da hierfür noch kein CTL-Epitop beschrieben war, welches zu den HLA-Allelen des Patienten passte, sollte geklärt werden, wie die exakte Sequenz des Epitops lautet und über welches HLA-Allel die Präsentation des Epitops erfolgt. Daher wurden in einer gemeinsamen Arbeit mit Frau A. Hückelhoven mehrmals spezifische CTL Linien von 0284 mit diesem Peptid generiert. Diese wurden in Restriktionsanalysen eingesetzt, um das präsentierende HLA-Molekül zu identifizieren. Dabei erwies sich in wiederholenden Versuchen, auch mit CTL Linien von anderen Spendern, das HLA-Allel A*02 als das restringierende Molekül (Abbildung 4.32). Kreuzreaktionen zu den überlappenden Peptiden Gag und Gag traten in keinen getesteten Linien auf, so dass das Epitop auf die Sequenz RPEPTAPPE eingrenzt werden konnte. Jedoch passt diese Sequenz nicht zum bekannten Bindungsmuster des HLA-A*02-Moleküls. Da außerdem die Inkubation der CTL Linie sowohl mit einem Anti- CD4 als auch mit einem Anti-CD8 Antikörper die Reaktion inhibierte, handelte es sich offensichtlich um eine Linie, die spezifisch für ein CD4-Epitop und ein CD8-Epitop ist. 77
86 4 Ergebnisse Abbildung 4.32 Restriktionsanalyse mit einer Gag spezifischen CTL Linie offenbarte, dass sich in dem Peptid ein bisher unbekanntes HLA-A*02-restringiertes CTL-Epitop befindet. Durch Inkubation mit dem Peptid Gag und Kultivierung für 10 Tage in R10IL2-Medium wurde aus PBMC von 0284 eine CTL Linie generiert. B-LCL, die möglichst nur ein HLA-Allel mit der CTL Linie teilen, wurden sowohl mit als auch ohne Peptid für eine Stunde inkubiert, 3x gewaschen und anschließend wurden 5x10 4 B-LCL mit 5x10 4 Zellen der CTL Linie im γ-ifn ELISPOT Assay ko-inkubiert. Nur die Peptid-inkubierte, HLA-A*02-positive B-LCL wurde von der CTL Linie erkannt und zeigte eine deutliche γ-ifn Sekretion. Angegeben sind nur die HLA-Allele, die B-LCL und CTL Linie gemeinsam haben. Schwarze Balken stellen die Peptid-inkubierten B-LCL dar, weiße Balken die Kontrolle ohne Peptid. HLA-I CTL Linie: A*02, A*11, B*39, B*57, Cw*06, Cw* Das Peptid Gag07-02 enthält ein HLA-B*41-restringiertes CTL-Epitop Da das Peptid Gag07-02 eine T-Zellantwort bei Patient 0925 auslöste und darin kein bekanntes CTL-Epitop vorhanden war, sondern nur potentielle Epitope zu den HLA- Allelen A*66 bzw. B*40 aus der Datenbank vorgeschlagen wurden (Tabelle 4.8), wurde eine spezifische CTL Linie zu diesem Peptid ausgehend von PBMC des Patienten 0925 generiert. Diese wurde anschließend in eine Restriktionsanalyse und in Kreuzreaktionen eingesetzt, um sowohl die Sequenz des Epitops eingrenzen zu können sowie auch das restringierende HLA-Allel zu bestimmen. Wie in Abbildung 4.33 A dargestellt, wird das Peptid nur von einer HLA-B*41-positiven B-Zelllinie präsentiert und demzufolge von der spezifischen CTL Linie erkannt. Zudem wies die Inhibierung der Reaktion durch den Anti- 78
87 4 Ergebnisse CD8 Antikörper ebenfalls auf ein HLA Klasse I-Epitop hin. Kreuzreaktionen mit den Peptiden Gag07-01 und Gag07-03 führten zu keiner Erkennung von Gag07-01, jedoch zu einer schwachen Erkennung von Gag07-03 (Abbildung 4.33 B). Damit lässt sich das Epitop auf die Sequenz SGGELDRWEKI einschränken. Da für das Molekül HLA-B*41 neun Aminosäuren lange Epitope optimal sind und sich an den Ankerpositionen ein Glutamat (Position zwei) bzw. ein Leucin (Position neun) befinden sollen (137), lautet die Sequenz des Epitops vermutlich GELDRWEKI. Darüber hinaus würde dies dem potentiellen HLA-B*40 Epitop (siehe Tabelle 4.8) entsprechen. Allerdings müsste man mit kürzeren Peptiden noch beweisen, ob HLA-B*41 wirklich das gleiche Epitop präsentiert wie HLA-B*40. Abbildung 4.33 Restriktionsanalyse und Kreuzreaktionen einer Gag07-02-spezifischen CTL Linie identifizierten ein HLA-B*41-restringiertes CTL-Epitop, welches vermutlich die Sequenz GELDRWEKI besitzt. PBMC von 0925 wurden durch Peptidinkubation mit Gag07-02 zur CTL Linie generiert. A: Nur die B- LCL mit dem HLA-Allel B*41 konnte das Epitop präsentieren und wurde daher von der CTL Linie erkannt. 1x10 5 Zellen der CTL Linie wurden zusammen mit 5x10 4 Zellen von gematchten B-Zelllinien, die möglichst nur ein HLA-Allel mit der CTL Linie teilen, in einem γ-ifn ELISPOT Assay inkubiert. Zuvor wurden die B-Zelllinien entweder mit dem Peptid Gag07-02 (schwarze Balken) oder ohne Peptid (weiße Balken) für eine Stunde inkubiert und daraufhin 3x gewaschen. Angegeben sind nur die HLA-Allele, welche der B-LCL und der CTL Linie gemeinsam sind. B: Die Gag07-02-spezifische CTL Linie erkannte sowohl die Peptide Gag07-02 als auch Gag07-03 und wurde durch einen Anti-CD8 Antikörper geblockt. 1x10 4 Zellen der CTL Linie wurden allein, mit den überlappenden Peptiden Gag07-01, -02 oder -03 oder einen Anti-CD8-Antikörper in einem γ-ifn ELISPOT Assay inkubiert. HLA-I CTL Linie: A*01, A*66, B*41, B*44, Cw*16, Cw*17 79
88 4 Ergebnisse Die restringierenden HLA-Allele für die Peptide Gag07-11 und Gag07-75 konnten nicht bestimmt werden Die Inkubation der PBMC von 0925 mit den Peptiden Gag07-11 und Gag07-75 löste eine T-Zellreaktion im ELISPOT aus. Jedoch waren für diese Peptide bisher noch keine CTL- Epitope beschrieben. Daher sollten sowohl die exakten Sequenzen der beiden Epitope als auch die HLA-Allele, welche das entsprechende Epitop präsentieren, bestimmt werden. Für das Peptid Gag07-11 wurden Kreuzreaktionen und Restriktionsanalysen mit der spezifischen CTL Linie durchgeführt. Eine Inkubation mit den beiden überlappenden Peptiden Gag07-10 sowie Gag07-12 führte zu keiner Freisetzung von γ-ifn (Abbildung 4.34). Folglich sollte unter der Annahme eines maximal zehn Aminosäuren langen Epitops das Epitop im Sequenzbereich RFAVNPGLLET liegen. Die Reaktion der CTL Linie konnte mit einem Anti-CD8 Antikörper gehemmt werden, so dass es sich um ein HLA Klasse I- Epitop handeln muss (Abbildung 4.34). In mehreren Restriktionsanalysen konnte kein restringierendes HLA-Allel identifiziert werden, da die HLA-Allele Cw*16 und Cw*17 ein Alanin an Position zwei des Epitops bevorzugen, könnte es sich um ein HLA-Crestringiertes CTL-Epitop handeln. Weitere Analysen könnten zur Bestätigung dieser Vermutung beitragen. Eine T-Zellantwort auf das Peptid Gag07-75 konnte bereits beim nächsten Untersuchungstermin des Patienten nicht mehr festgestellt werden, so dass keine Studien mehr folgten. Abbildung 4.34 Kreuzreaktionen und CD8-Block der Gag07-11-spezifischen CTL Linie. 1x10 5 Zellen der spezifisch generierten CTL Linie von Patient 0925 wurden ohne Peptid, mit den dargestellten Peptiden und mit einem Anti-CD8-Antikörper im γ-ifn ELISPOT Assay inkubiert. Es wurde keine Kreuzreaktion zu den überlappenden Peptiden beobachtet, was die Sequenz des Epitops auf RFAVNPGLLET eingrenzt. Die Reaktion konnte mit dem Anti-CD8 Antikörper geblockt werden. 80
89 4 Ergebnisse Die Peptide Nef07-08 und Nef07-10 enthalten HLA-B*41-Epitope Zwar wurden für die beiden Nef-Peptide Nef07-08 und Nef07-10 potentielle Epitope aus der Datenbank vorgeschlagen, dennoch ist unklar, welches CTL-Epitop in dem jeweiligen Peptid liegt und die T-Zellantwort bei Patient 0925 auslöste (Tabelle 4.8). Daher wurden spezifische CTL Linien von 0925 mit diesen Peptiden generiert und diese nach positiver Testung im γ-ifn ELISPOT in Restriktionsanalysen und Kreuzreaktionen eingesetzt. Beide CTL Linien erkannten dabei nur Peptid-inkubierte B-Zelllinien, die das HLA-Allel B*41 tragen (Abbildung 4.35). Da die Immunreaktionen bei beiden CTL Linien zumindest teilweise mit einem Anti-CD8 Antikörper geblockt werden konnte (Abbildung 4.36), kann man jeweils auf ein CTL-Epitop schließen, das über das Molekül HLA-B*41 restringiert ist. Abbildung 4.35 Restriktionsanalysen mit Nef und Nef07-10-spezifischen CTL Linien wiesen auf HLA-B*41-restringierende CTL-Epitope in den jeweiligen Peptiden hin. Nach Generierung der spezifischen CTL Linien aus PBMC von Patient 0925 wurden B-LCL ausgewählt, welche möglichst nur ein HLA-Allel mit der CTL Linie gemeinsam haben. Diese wurden einmal mit und einmal ohne Peptid inkubiert, gewaschen und 5x10 4 B-LCL mit 1x10 5 Zellen der CTL Linie in einem γ-ifn ELISPOT Assay inkubiert. Angegeben sind nur die gemeinsamen HLA-Allele der B-Zelllinie und der CTL Linie. A: Die Restriktionsanalyse mit der Nef07-08-spezifischen CTL Linie zeigte eine klare Reaktion bei Ko-Inkubation mit der HLA-B*41-positiven B-LCL, die zuvor mit dem Peptid Nef07-08 inkubiert wurde (blau). B: Die Restriktionsanalyse mit der Nef07-10-spezifischen CTL Linie demonstrierte ebenso eine klare Erkennung der HLA-B*41-positiven B-LCL, die zuvor mit dem Peptid Nef07-10 inkubiert wurde (magenta). HLA-I CTL Linie: A*01, A*66, B*41, B*44, Cw*16, Cw*17 81
90 4 Ergebnisse Kreuzreaktionen der Nef07-08-spezifischen CTL Linie mit den Peptiden Nef07-07 und Nef07-09 zeigten keine Erkennung der beiden überlappenden Peptide (Abbildung 4.36 A), so dass das Epitop im Bereich der Sequenz AVSRDLEKHG liegen müsste. Die Ankeraminosäuren würden bei einem HLA-B*41-restringierten Epitop Glutamat an Position zwei und Leucin an Position neun sein (137), welche jedoch in der Sequenz nicht vorkommen. Das Programm SYFPEITHI sagt die Sequenz SRDLEKHGA für das HLA- B*41-Epitop vorher. In diesem Fall hätte man aber eine Kreuzreaktion der Nef spezifischen CTL Linie mit dem Peptid Nef07-09 erhalten müssen. Daher ist noch unklar, wie die exakte Sequenz des Epitops ist. Unsere Ergebnisse stimmen nicht mit dem vorgeschlagenen, potentiellen Epitop überein (ELF, HIV sequence database), da wir anstelle einer HLA-A*66-Restriktion eine HLA-B*41-Restriktion feststellten. Kreuzreaktionen der Nef07-10-spezifischen CTL Linie demonstrierten eine Erkennung des überlappenden Peptides Nef07-09, aber nicht des Peptides Nef07-11 (Abbildung 4.36 B). Dadurch muss das Epitop in der Sequenz LEKHGAITSSN liegen, welche sowohl in Nef07-09 als auch Nef07-10 vorhanden ist. Da HLA-B*41 an Position zwei ein Glutamat als Ankeraminosäure bevorzugt und meist Liganden mit einer Länge von neun Aminosäuren bindet, könnte das HLA-B*41 CTL-Epitop in Nef07-10 dem potentiellen HLA-B*40 LEKHGAITS entsprechen (siehe Tabelle 4.8). Dies soll durch die Verwendung von kürzeren Peptiden noch bewiesen werden. Abbildung 4.36 Kreuzreaktionen und CD8-Block mit den CTL Linien, die spezifisch für Nef07-08 bzw. Nef07-10 sind. 1x10 5 Zellen einer spezifisch generierten CTL Linie von Patient 0925 wurden ohne Peptid, mit den dargestellten Peptiden und mit einem Anti-CD8-Antikörper im γ-ifn ELISPOT Assay inkubiert. A: Die Nef07-08-spezifische CTL Linie (blau) zeigte keine Kreuzreaktion zu den überlappenden Peptiden, was die Sequenz des Epitops auf AVSRDLEKHG eingrenzt. Die T-Zellantwort konnte mit dem Anti-CD8 Antikörper geblockt werden. B: Die Nef07-10-spezifische CTL Linie (magenta) erkannte das überlappende Peptid Nef07-09, so dass das CTL-Epitop auf die Sequenz LEKHGAITSSN eingegrenzt werden konnte. Die Inkubation mit dem Anti-CD8-Antikörper inhibierte die Sekretion von γ-ifn um die Hälfte. 82
91 4 Ergebnisse Das bekannte HLA-C-restringierte Nef-Epitop Nef11T3 ist ein häufig erkanntes Epitop Das CTL-Epitop Nef11T3 RQDILDLWIY wird über HLA-Cw*07 restringiert (138). Da HLA- C-Moleküle durch Nef nicht negativ beeinflusst werden können, sind HLA-Crestringierende Epitope interessante Kandidaten bei einer Vakzinierung zur Induktion von CTLs. Aufgrund der Häufigkeit des HLA-Allels Cw*07 sollte untersucht werden, wie oft und in welchem Ausmaß eine CTL-Antwort gegen dieses Epitop auftritt. Dazu wurden CTL Linien von HIV-1-infizierten Patienten generiert, welche HLA-Cw*07-positiv sind. Nach zehntägiger Kultivierung wurden die Zellen im γ-ifn ELISPOT Assay getestet. Insgesamt wurden 85 Patienten in 92 Assays untersucht. Von diesen 85 Patienten wiesen 36 eine CTL-Antwort gegen dieses Epitop auf (42%), 19 davon sogar eine ausgeprägte Immunantwort mit mehr als 250 SFU (22%). Anhand der autologen Nef Sequenzen, die uns zur Verfügung standen, konnte eine Variante des Epitops festgestellt werden. Bei dieser, Nef11T3 I/V RQDILDLWVY, findet man an Position neun ein Valin (V) anstelle eines Isoleucins (I). Da diese in den autologen Virussequenzen der Patienten häufig zu finden war (siehe auch Abbildung 4.10 B), wurden von 35 Patienten auch CTL Linien mit dem varianten Peptid generiert und analysiert. Bei 15 Patienten (43%) konnte hierbei eine CTL-Antwort gegen das Peptid Nef11T3 I/V detektiert werden, neun davon (26%) wiesen eine starke Immunreaktion mit über 250 SFU auf. Des Weiteren waren alle Nef11T3- spezifischen CTL Linien kreuzreaktiv zu Nef11T3 I/V und alle Nef11T3 I/V-spezifischen CTL Linien erkannten das Wildtyp Peptid. Bei Titrationsversuchen konnte eine höhere Affinität der Variante detektiert werden, da eine Erkennung des Peptids zum Teil bis zu einer Konzentration von 10 ng/ml stattfand (Abbildung 4.37 B). Im Falle des Wildtyp Peptids führten nur Konzentrationen bis 100 ng/ml zu einer Sekretion von γ-ifn (Abbildung 4.37 A). Zusammenfassend kann festgehalten werden, dass das HLA-Cw*07-restringierende Peptid Nef11T3 sowie die Variante Nef11T3 I/V sehr häufig von HIV-1-infizierten Patienten erkannt wurden. 83
92 4 Ergebnisse Abbildung 4.37 Titrationsversuche mit Nef11T3- und Nef11T3 I/V-spezifischen CTL Linien zeigten eine höhere Affinität von Nef11T3 I/V. Von den generierten CTL Linien wurden 1x10 5 Zellen im γ-ifn ELISPOT Assay mit verschiedenen Konzentrationen von Peptiden inkubiert. A: Titration des Nef11T3-Peptids (schwarz) und Kreuztitration des Peptids Nef11T3 I/V (blau) mit einer Nef11T3-spezifischen CTL Linie. Beide Peptide wurden von der CTL Linie nur bis zu einer Peptidkonzentration von 100 ng/ml erkannt. B: Titration des Peptides Nef11T3 I/V (blau) und Kreuztitration des Peptides Nef11T3 (schwarz) mit einer Nef11T3 I/Vspezifischen CTL Linie. Das Peptid Nef11T3 I/V wurde auch bei einer Konzentration von 10 ng/ml noch deutlich erkannt. 84
93 4 Ergebnisse 4.5 Etablierung alternativer Transfektionsmethoden Zur Transfektion von mrna in vitro stellt die Elektroporation die vermutlich schnellste, kostengünstigste und effizienteste Methode dar. Für eine in vivo Applikation fehlen aktuell noch die technischen Voraussetzungen. Daher sollten alternative Transfektionsmethoden etabliert werden, die im Hinblick auf eine in vivo Anwendung sicher und gut verträglich sind. Für alle in dieser Arbeit untersuchten Reagenzien konnte bereits ein erfolgreicher Transfer von DNA und zum Teil von sirna in Zellen gezeigt werden, ein Transfer von mrna jedoch noch nicht Chitosan transfiziert DNA, aber keine mrna in HEK293T Bei Chitosan handelt es sich um ein natürlich vorkommendes Polysaccharid aus dem Panzer von Krebsen (139), welches DNA und sirna in verschiedene Zellen transportieren kann (140, 141). Aufgrund seiner nicht-toxischen, biologisch abbaubaren und biokompatiblen Eigenschaften stellt es eine interessante Möglichkeit zur Transfektion in vivo dar. Daher wurden Transfektionen von GFP mrna, welche mit Chitosan komplexiert ist, in HEK293T Zellen, PBMC, Makrophagen und Dendritische Zellen (DCs) durchgeführt. Zur Kontrolle wurden die Zellen auch mit der Plasmid-DNA pegfp-c1 transfiziert. Es konnte ein erfolgreicher Transfer der mit Chitosan komplexierten Plasmid- DNA in HEK293T Zellen bereits nach 22 Stunden detektiert werden, welche nach 120 Stunden besonders deutlich war (Abbildung 4.38). Zellen, welche mit komplexierter GFP mrna transfiziert wurden, zeigten auch nach 120 Stunden keine Fluoreszenz. In PBMC, Makrophagen und DCs konnte weder nach DNA-Transfektion noch nach mrna- Transfektion eine GFP-Fluoreszenz gemessen werden. Folglich war mit Chitosan nur die Transfektion von Plasmid-DNA in HEK293T Zellen erfolgreich, alle anderen Versuche schlugen fehl. 85
94 4 Ergebnisse Abbildung 4.38 Chitosan komplexierte Plasmid-DNA, wodurch eine erfolgreiche Transfektion der DNA in HEK293T Zellen stattfinden konnte. Sowohl GFP mrna als auch Plasmid-DNA von pegfp-c1 wurde mit einer Chitosanlösung vermischt und anschließend zu HEK293T Zellen gegeben. Durch Detektion der GFP-Fluoreszenz an einem Fluoreszenzmikroskop konnte der Erfolg der Transfektion ermittelt werden. Die grüne Fluoreszenz ist in der Fluoreszenzaufnahme in weiß dargestellt (rechts) und zeigt wenige GFPexprimierende Zellen. Vergleicht man dazu die Durchlichtaufnahme (links), welche die Dichte der Zellen zeigt, wird deutlich, dass nur wenige Zellen GFP exprimieren. Diese Aufnahmen wurden 120 Stunden nach Transfektion dokumentiert HIV-1 Tat führt zu keinem Transfer von Nukleinsäuren in Zellen Verschiedene Studien mit dem HIV-1 Tat-Protein oder einem einzelnen Peptid, welches die Protein-Transduktionsdomäne von Tat enthält, zeigten bereits eine erfolgreiche und sichere in vivo-transfektion von Proteinen, sirna und DNA ( ). In dieser Arbeit wurden HEK293T und Makrophagen zur Transfektion mit Tat-Protein und Tat-Peptid verwendet. Sowohl GFP mrna als auch GFP-DNA in Form des Plasmides pegfp-c1 kamen zum Einsatz. Allerdings konnte in keinem Versuch ein erfolgreicher Transfer der Nukleinsäure mit Hilfe des Tat-Proteins bzw. des Tat-Peptides beobachtet werden. Auch nach Veränderung der Ratio zwischen Tat und DNA bzw. mrna konnte keine GFP- Fluoreszenz detektiert werden Atelokollagen transfiziert weder DNA noch mrna in HEK293 und HEK293T Atelokollagen ist ein Pepsin-verdautes Typ I Kollagen und kann aufgrund seiner positiven Ladung mit negativ geladenen Nukleinsäuren wie DNA oder sirna Komplexe bilden und diese in Zellen übertragen. Atelokollagen ist als Biomaterial sehr sicher einsetzbar und weder toxisch noch immunogen (145). Eine Transfektion von sirna und DNA über diese 86
95 4 Ergebnisse Form der Komplexbildung wurde bereits mehrfach gezeigt (146, 147). Es wurde sowohl GFP mrna als auch als Kontrolle GFP-DNA (pegfp-c1) mit Atelokollagen vermengt und anschließend die Transfektion der Komplexe in HEK293 und HEK293T durchgeführt. Allerdings war keine der durchgeführten Transfektionen erfolgreich und auch diverse Abänderungen im Protokoll der Transfektion führten zu keiner Detektion der GFP- Fluoreszenz FuGENE kann neben DNA auch mrna erfolgreich in HEK293T Zellen transfizieren Bei allen Transfektionsversuchen (4.5.1 bis 4.5.3) wurde als Kontrolle die Plasmid-DNA pegfp-c1 mit FuGENE in die verwendeten Zellen HEK293T, HEK293, PBMC, DCs und Makrophagen transfiziert. Zudem wurde auch versucht GFP mrna mit FuGENE zu transfizieren. Dabei konnte neben der erfolgreichen Transfektion von HEK293T und HEK293 Zellen mit Plasmid-DNA auch ein Transfer der GFP mrna in HEK293T beobachtet werden. Zwar waren nur wenige Zellen schwach positiv für GFP, aber die Fluoreszenz war dennoch klar zu erkennen (Abbildung 4.39). In den weiter genannten Zelltypen wurde keine GFP Fluoreszenz detektiert. Der Transfer von mrna wurde für die Verwendung von FuGENE zum damaligen Zeitpunkt noch nicht beschrieben. Abbildung 4.39 GFP mrna konnte mit Hilfe von FuGENE in HEK293T Zellen transfiziert werden. HEK293T Zellen wurden sowohl mit GFP-DNA als auch mit GFP mrna transfiziert, indem zuvor die Nukleinsäuren mit FuGENE komplexiert wurden. Die erfolgreiche Transfektion wurde anhand der GFP- Fluoreszenz an einem Fluoreszenzmikroskop ermittelt. Links ist die Durchlichtmikroskop-Aufnahme zu sehen, im rechten Bild dazu ist der gleich Ausschnitt in der Fluoreszenzaufnahme dargestellt, wobei die grüne Fluoreszenz in weiß dargestellt wird. Wenige Zellen zeigten eine schwache Expression von GFP. Die Aufnahmen wurden 48 Stunden nach Transfektion erstellt. 87
96 4 Ergebnisse 4.6 Analyse des Sequenzverlaufs und der Immunantworten eines HIV-1-infizierten Patientenpaares, das mit demselben Virus infiziert worden war Unterschiedliche Erkennung der homologen und heterologen viralen mrna-konstrukten Bei den HIV-1-infizierten Patienten 0225 und 0284 handelt es sich um ein homosexuelles Paar mit demselben Ausgangsvirus, wobei 0284 seinen Partner 0225 kurz nach seiner eigenen Infektion infizierte. Jedoch zeigen die beiden Patienten einen divergenten Krankheitsverlauf. So konnte das Immunsystem von 0284 das Virus sehr schnell gut kontrollieren, so dass die Viruslasten stetig sanken und inzwischen bei 80 Kopien/ml liegen. Patient 0225 hingegen konnte die Replikation des Virus von selbst nicht eindämmen, wodurch schon frühzeitig wegen einer persistierenden hohen Virämie eine ART eingeleitet wurde. Von beiden Patienten standen uns nach Klonierung die autologen gag und nef Gene in Form von mrna zur Verfügung. Ob das eng verwandte Virus des Partners vergleichbare CTL-Antworten induziert wie das autologe Virus, sollte mit Hilfe der mrna-elektroporation überprüft werden. PBMC des Patienten 0225 wurden zum einen mit den drei verschiedenen gag mrnas (autolog, 0284 und SF2) sowie zum anderen mit den drei verschiedenen nef mrnas (autolog, 0284 und SF2) elektroporiert und anschließend in einen γ-ifn ELISPOT Assay eingesetzt. Ebenso wurde bei Patient 0284 verfahren. Unbehandelte PBMC, mock-elektroporierte und GFP mrnaelektroporierte PBMC dienten stets als Kontrolle und führten zu keiner γ-ifn Sekretion. In Abbildung 4.40 ist das Ergebnis dieser Analyse dargestellt. Beide Patienten wiesen, wie bereits unter erwähnt, eine stärkere CTL-Antwort gegen das autologe Gag-Protein im Vergleich zu SF2 auf (Abbildung 4.40 obere Reihe). Allerdings differenzierten sich die Immunantworten im Hinblick auf das Virus des Partners. So zeigte Patient 0284 eine sehr starke CTL-Antwort gegen das Gag-Protein von 0225 (181 SFU/2x10 5 PBMC) ähnlich wie gegen das autologe Virus (252 SFU/2x10 5 PBMC). Zellen des Patienten 0225 erkannten dagegen das Gag-Protein seines Partners am schlechtesten (7 SFU/2x10 5 PBMC). Unterschiede in der Proteinexpression sind für die verschieden stark ausgelösten Immunantworten in den beiden Patienten nicht verantwortlich, da die Expression der verschiedenen Gag-Proteine um maximal 8% variierte (Daten nicht gezeigt). 88
97 4 Ergebnisse Für die CTL-Antworten gegen das Nef-Protein (Abbildung 4.40 untere Reihe) konnten ebenso keine einheitlichen Ergebnisse erzielt werden. In PBMC des Patienten 0225 wurde trotz der höchsten Expression des autologen Nef-Proteins (58%) eine sehr schwache T-Zellreaktion detektiert (8 SFU/2x10 5 PBMC). Die beste Immunantwort wurde gegen das SF2 Nef erzielt (37 SFU/2x10 5 PBMC) (Abbildung 4.40). Das SF2 Nef-Protein konnte in 25% der Zellen detektiert werden, während nur 8% der elektroporierten Zellen für 0284 Nef positiv waren. Daher ist unklar, ob eine vergleichbare starke CTL-Antwort durch das 0284 Nef erreicht werden hätte können, wenn die Expression des 0284 Nef- Proteins stärker gewesen wäre. In PBMC des Patienten 0284 hingegen löste das autologe Nef-Protein die stärkste Immunantwort aus (33 SFU/2x10 5 PBMC), die Immunreaktionen gegen SF2 Nef und 0225 Nef waren trotz höherer oder gleicher Expression deutlich geringer (Abbildung 4.40). Abbildung 4.40 Virale Gene, welche unterschiedlich sind, jedoch dasselbe Ausgangsvirus haben, lösten nicht immer stärkere Immunantworten aus als Referenzviren. Die Patienten 0225 und 0284 haben dasselbe Ausgangsvirus. PBMC von Patient 0225 und 0284 wurden jeweils mit SF2 nef, 0225 nef und 0284 nef mrna sowie mit SF2 gag, 0225 gag und 0284 gag mrna elektroporiert. Nach vier Stunden wurden die Zellen in einem γ-ifn ELISPOT Assay inkubiert, um die T-Zellantworten gegen die verschiedenen mrnakodierten Proteine zu messen. Dargestellt sind die SFU/2x10 5 PBMC. Die obere Reihe zeigt die T-Zellantworten gegen die Gag-Proteine, die untere die T-Zellantworten gegen die Nef-Proteine (linke Spalte: 0225, rechte Spalte: 0284). 89
98 4 Ergebnisse Somit kann festgehalten werden, dass die T-Zellantworten auf eng verwandte Viren, die dasselbe Ausgangsvirus haben, in Patienten sehr unterschiedlich sein können und nicht unbedingt zu einer besseren Immunreaktion führen als Referenzviren Die viralen autologen Sequenzen erklären die detektierten Immunantworten Weshalb die verschiedenen mrna-kodierten HIV-1-Proteine zu den kontroversen Immunantworten in den beiden Patienten führten, sollte durch eine vergleichende Analyse der entsprechenden viralen Sequenzen geklärt werden. Patient 0284 zeigte die stärkste Immunreaktion gegen das autologe Gag-Protein, gefolgt von der Immunreaktion gegen das Gag-Proteins seines Partner (0225) und die schwächste Reaktion wurde gegen SF2 Gag detektiert (Abbildung 4.40). In Gag befinden sich viele CTL-Epitope, welche durch HLA-Moleküle des Patienten 0284 präsentiert werden können. Die Mehrheit weist keine bis wenige Mutationen auf (Abbildung 4.41 A). Das HLA-A*02-restringierte Epitop FLGKIWPSHK (AS ) (123) zeigt in der Sequenz des SF2-Proteins einen H-zu-Y Austausch an Position neun. Ebenso findet sich in der SF2 Gag Sequenz ein E-zu-D Austausch an Position fünf des HLA-B*57- restringierten Epitops QASQEVKNW (AS ) (120). Die Sequenz des HLA-A*02- restringierten Epitops SL9 SLYNTVATL (AS 77-85) (27) unterscheidet sich sowohl in der SF2 Gag Sequenz als auch in der 0225 Gag Sequenz. Zusätzlich zu dem SL9-Epitop weisen auch die Epitope EMMTACQGV (HLA-A*02, AS ) (148), ACQGVGGPGHK (HLA-A*11, AS ) (149) und TLYCVHQR (HLA-A*11, AS 84-91) (150) in der Sequenz von 0225 Aminosäuren-Austausche auf. Insbesondere die R-zu-N Substitution an Position acht des Epitops TLYCVHQR in der Sequenz von 0225 führte wahrscheinlich zum Verlust des Epitops und trug damit zur geringeren T-Zellantwort auf 0225 Gag bei. Somit unterscheiden sich die Sequenzen von SF2 und 0225 zur Sequenz von 0284 in sechs CTL-Epitopen, welche vermutlich durch CTLs von 0284 schlechter erkannt werden wie die autologen Varianten. Auch in der autologen Sequenz von 0284 findet sich im Epitop TSTLQEQIGW (HLA-B*57, AS ) (120) eine CTL Fluchtmutante und zudem eine Prozessierungsmutante vor dem CTL-Epitop ISPRTLNAW (HLA-B*57, AS ) (151). Da die Immunantwort gegen das autologe Gag-Protein dennoch am stärksten war, kann man vermuten, dass der Patient weitere, für seine Sequenz spezifische Epitope erkannte. Patient 0225 zeigte die stärkste Immunreaktion gegen sein autologes Gag-Protein. Die Gag-Proteine von SF2 und seinem Partner 0284 induzierten deutlich geringere CTL- 90
99 4 Ergebnisse Antworten, wobei 0284 Gag die schwächste T-Zellreaktion in PBMC von 0225 auslöste. Im Alignment der Gag Sequenzen ist ersichtlich, dass bei Patient 0225 aufgrund seiner HLA-Allele weniger publizierte CTL-Epitope in Gag vorliegen als bei Patient 0284 (Abbildung 4.41 A). Außerdem weisen die Epitope SL9 SLYNTVATL (HLA-A*02, AS 77-85) (27) und KL14 KIRLRPGGKKKYKL (HLA-A*03, AS 18-31) (116) Mutationen in SF2 Gag und 0284 Gag auf und sind so vermutlich für die schlechtere Erkennung der entsprechenden Proteine verantwortlich. Das autologe Nef-Protein bewirkte eine stärkere T-Zellreaktion in Zellen des Patienten 0284 als das SF2 Nef bzw. das Nef-Protein seines Partners 0225 (Abbildung 4.40). Bei einem Vergleich der Aminosäurensequenzen der verschiedenen Nef-Proteine konnten in drei Epitopen Unterschiede ermittelt werden. Die Epitope AITSSNTAA (HLA-A*02, AS 52-60) (152), HTQGYFPDWQ (HLA-B*57, AS ) (153) und AFHHMAREL (HLA-A*02, AS ) (113) haben in der autologen Sequenz von 0284 eine veränderte Aminosäuren-Abfolge als in den beiden heterologen Proteinen. Wenn es sich bei diesen Veränderungen um Fluchtmutationen gehandelt haben sollte, beweist die stärkere Erkennung der autologen Variante, dass das Immunsystem in der Lage ist, neue CTL- Spezifitäten zu generieren, welche spezifisch die mutierten Varianten erkennen können. Für Patient 0225 konnte eine sehr geringe Immunreaktion gegen sein autologes Nef-Protein detektiert werden, wohingegen SF2 Nef und 0284 Nef stärkere CTL- Antworten induzierten (Abbildung 4.40). Eine vergleichende Analyse der Aminosäurensequenzen offenbart im HLA-A*03-restringierten CTL-Epitop QVPLRPMTYK (AS 83-92) (118) einen T-zu-N Austausch an Position acht in der autologen Sequenz (Abbildung 4.42 A). Es handelt sich dabei um eine CTL Fluchtmutante, welche zur verminderten Erkennung der autologen Nef-Variante führte. Auch die beiden Epitope AVRERMRRT (AS 15-23) (154) und AFHHMAREL (AS ) (113), welche über die HLA-Allele B*07 bzw. A*02 präsentiert werden, beinhalten Mutationen in der autologen Sequenz (Abbildung 4.42 A). Da SF2 Nef und 0284 Nef weder in den genannten Epitopen noch in anderen CTL-Epitopen des Patienten 0225 Aminosäuren-Austausche aufweisen (Abbildung 4.42 A), lösten die beiden heterologen Proteine stärkere T-Zellreaktionen aus als das autologe Protein. Folglich deckte der Vergleich der Proteinsequenzen Unterschiede in CTL-Epitopen auf, welche die divergente Erkennung der HIV-1-Proteine der Patienten 0225 und 0284 erklären könnten. 91


100 4 Ergebnisse Abbildung 4.41 Alignment der Aminosäurensequenzen des Gag- und des Nef-Proteins von SF2, 0225 und 0284 offenbart in den betreffenden CTL-Epitopen der beiden Patienten CTL Fluchtmutationen. Die Alignments und die Consensus-Sequenz wurden mit der Vector NTI Software erstellt. Grau unterlegte Bereiche stellen Aminosäuren dar, die zwischen den drei Sequenzen unterschiedlich sind. CTL-Epitope zum HLA-Typ von Patient 0225 sind grün umrandet, CTL-Epitope zum HLA-Typ von Patient 0284 sind blau umrandet. Rot umrandet wurden CTL-Epitope, welche von beiden Patienten präsentiert werden können und HLA-A*02 oder HLA-Cw*07 restringiert sind. A: Alignment der Aminosäurensequenzen der verwendeten gag mrnas. B: Alignment der Aminosäurensequenzen der verwendeten nef mrnas HLA-I: A*02, A*03, B*07, B*18, Cw*05, Cw*07; 0284 HLA-I: A*02, A*11, B*39, B*57, Cw*06, Cw*07 92
101 4 Ergebnisse In Patient 0225 mutierte das Virus im selben Zeitraum deutlich mehr als in Patient 0284 Da sich trotz desselben Ausgangsvirus erhebliche Unterschiede in den viralen Gag und Nef Sequenzen der beiden Patienten 0225 und 0284 feststellen ließen (Abbildung 4.41 A und B), stellte sich die Frage, welche Mutationen bereits im Ausgangsvirus vorhanden waren und welche sich erst in einen der beiden Probanden entwickelt haben. Daher wurde aus Plasmaproben der Patienten, welche zum Zeitpunkt der Erstdiagnose (ED) im August 2006 aserviert wurden, virale RNA isoliert und diese nach spezifischer cdna- Synthese und PCR sequenziert. Es konnte dabei die vollständige Gag Sequenz von 0284 und die vollständige Nef Sequenz von 0225 erhalten werden. Für Gag 0225 fehlen die Aminosäuren 44 bis 148 und für Nef 0284 die ersten 76 Aminosäuren. In diesen Bereichen wurden mehrere Sequenzen gleichzeitig detektiert, auch die Verwendung von verschiedenen Primern bei der Sequenzierreaktion zeigte keinen Erfolg. In den Gag Sequenzen von 0225 werden viele Ungleichheiten zwischen den beiden Zeitpunkten deutlich (Abbildung 4.42 A). So können bereits zu Beginn der Sequenz Mutationen identifiziert werden, welche durch Selektionsdruck ausgelöst durch CTLs entstanden sind. Die Aminosäure 30 ist ein Beispiel. An vielen anderen Positionen in Gag kann gut verfolgt werden, welche Viren sich in diesem Patienten manifestierten. So kamen zum Zeitpunkt der Erstdiagnose Viren vor, welche als Aminosäure 240 entweder ein Glycin (G) oder ein Arginin (R) trugen. Da sich in der späteren Sequenz nur noch Viren mit Glycin finden, haben sich diese Viren durchgesetzt. Viren mit Isoleucin (I) an Position 282 konnten sich gegen Viren mit Threonin (T) behaupten und die Aminosäure Asparagin (N) an Position 305 konnte sich gegen Threonin (T) manifestieren (Abbildung 4.42 A). Vergleicht man nun die Gag Sequenz von 0284, welche vom Zeitpunkt der Erstdiagnose stammt, mit der Sequenz, die der mrna entspricht, lassen sich wenige Differenzen ermitteln. Nur 14 Aminosäuren haben sich zwischen den beiden Zeitpunkten verändert (Abbildung 4.42 B). Beispielsweise finden sich bei Erstdiagnose sowohl Viren, die ein Threonin (T) an Aminosäureposition 242 tragen und solche mit einem Asparagin (N), wie es in der mrna-sequenz enthalten ist. Da die Aminosäure in einem CTL-Epitop liegt, fand vermutlich ein Selektionsdruck durch CTLs statt, der dann in einer Fluchtmutante resultierte. Die Aminosäure 338 lautete zum Zeitpunkt der Erstdiagnose noch Glycin (G) und mutierte zu Arginin (R), ebenso die Aminosäure 377, welche von Valin (V) zu Methionin (M) mutierte. Ursprünglich fanden sich zunächst Viren, die an Aminosäureposition 396 ein Arginin (R) besaßen. Letztendlich hat sich die Variante mit Lysin (K) durchgesetzt (Abbildung 4.42 B). 93
102 4 Ergebnisse Wie Gag zeigte auch der Sequenzvergleich für Nef von Patient 0225 sehr viele Differenzen zwischen den beiden Zeitpunkten (Abbildung 4.43 A). Neben vielfach vorkommenden Mutationen, die zu veränderten Aminosäuren führten, entwickelte das Virus auch eine Insertion von zehn Aminosäuren (Abbildung 4.43 A). Dass es sich bei dem T-zu-N Austausch im Epitop QK10 (AS 83-93) um eine CTL Fluchtmutante handelt, wird durch das Fehlen dieser Variante zu Beginn der Infektion bestätigt. Die autologe Sequenz des Epitops AFHHMAREL (AS ) war zum Ausgangszeitpunkt bereits vorhanden. Im CTL-Epitop AIRERMRRA (AS 15-23) jedoch zeigen sich deutliche Veränderungen. Das Nef-Protein des Virus hat sich in Patient 0284 zwischen den beiden Zeitpunkten kaum verändert. In dem Sequenz-Abschnitt, der uns zu Verfügung stand, konnten nur sechs mutierte Aminosäuren beobachtet werden (Abbildung 4.43 B). Das HLA-A*02- restringierte CTL-Epitop AFHHMAREL (AS ) wurde dabei sehr stark mutiert, insgesamt drei Aminosäuren-Austausche fanden statt, die letztendlich zum Verlust des Epitops führten. Durch diese Daten wird klar demonstriert, dass sich dasselbe Ausgangsvirus in zwei HIV-1-Infizierten sehr unterschiedlich entwickeln kann. Das HI-Virus hat sich in den Genen gag und nef in Patient 0225 stark verändert, in Patient 0284 im selben Zeitraum dagegen kaum. Da die beiden Patienten einen divergenten Krankheitsverlauf aufzeigen, wird erneut deutlich, dass nicht nur das Virus selbst, sondern auch Faktoren wie das eigene Immunsystem entscheidend zur Progression der Erkrankung beitragen. 94
103 4 Ergebnisse Abbildung 4.42 Alignment der Aminosäurensequenzen des Gag-Proteins von 0225 und 0284 zum Zeitpunkt der Erstdiagnose (ED) und nach ca. 2 Jahren, welche den mrna-sequenzen entsprechen. Die Alignments wurden mit der Vector NTI Software gebildet. Grau unterlegte Bereiche stellen Aminosäuren dar, die zwischen den Gag Sequenzen unterschiedlich sind. Aminosäuren, welche mit X in der Erstdiagnose auftauchen, sind in den beiden Möglichkeiten für X aufgeschlüsselt. A: Alignment der Gag Sequenzen des Patienten Es konnte keine vollständige Sequenz erhalten werden, so dass der Bereich zwischen den Aminosäuren fehlt und mit X als Platzhalter dargestellt ist. Hier ist kein Vergleich der Sequenzen zu den verschiedenen Zeitpunkten möglich. B: Alignment der Gag Sequenzen des Patienten
104 4 Ergebnisse Abbildung 4.43 Alignment der Aminosäurensequenzen des Nef-Proteins von 0225 und 0284 zum Zeitpunkt der Erstdiagnose (ED) und nach ca. 2 Jahren, welche den mrna-sequenzen entsprechen. Die Alignments wurden mit der Vector NTI Software gebildet. Grau unterlegte Bereiche stellen Aminosäuren dar, die zwischen den Nef Sequenzen unterschiedlich sind. Aminosäuren, welche mit X in der Erstdiagnose auftauchen, sind in den beiden Möglichkeiten für X aufgeschlüsselt. A: Alignment der Nef Sequenzen des Patienten B: Alignment der Nef Sequenzen des Patienten Es konnte keine vollständige Sequenz erhalten werden, so dass die ersten 76 Aminosäuren fehlen und mit X als Platzhalter dargestellt sind. Hier ist kein Vergleich der Sequenzen zu den verschiedenen Zeitpunkten möglich. 96
105 4 Ergebnisse 4.7 Interaktion zwischen HIV-1-spezifischen mrnas und zellulären mirnas Einzelne HIV-1-Gene beeinflussen das zelluläre mirna- Profil in CD4 + T-Zellen Durch Deep Sequencing konnten zelluläre mirnas identifiziert werden, deren Expressionsstärke von einzelnen HIV-1-Genen verändert wird Bisherige Studien zur Interaktion von zellulären mirnas und HIV-1 haben die Veränderung des mirna-profils meist in Zelllinien nach Transfektion mit einem HIV-1 NL4-3 Virusisolat oder in PBMC von HIV-1-infizierten Patienten untersucht (69, 155). In dieser Arbeit sollte nun eruiert werden, ob und in welcher Weise einzelne, virale Gene einen Einfluss auf das zelluläre mirna-muster haben. Da es sich bei CD4 + T-Zellen um die Zielzellen des HI-Virus handelt, sollten diese zur Analyse verwendet werden. Zu diesem Anlass wurden PBMC eines gesunden Spenders isoliert und daraus die CD4 + T- Zellen selektioniert. Die gewonnenen CD4 + T-Zellen wurden mit GFP mrna, HIV-1 nef sowie HIV-1 gag mrna elektroporiert. Es wurden dabei zwei nef mrnas aus verschiedenen Patienten mit unterschiedlichem Krankheitsverlauf verwendet. Patient 0225 konnte das Virus nicht kontrollieren und wurde daher mit ART behandelt, wohingegen Patient 0284 eine gute Kontrolle der Infektion zeigte und seit sechs Jahren keine Therapie benötigt. Durch die Verwendung dieser beiden nef mrnas sollte ermittelt werden, ob HIV-1-Gene von verschiedenen Probanden mirnas unterschiedlich beeinflussen können und somit eine Rolle in der Krankheitsprogression spielen. 24 Stunden nach Elektroporation wurde ein Aliquot der Zellen zur Detektion der Proteinexpression verwendet, während die restlichen Zellen geerntet wurden und daraus die Isolierung der gesamten, zellulären RNA erfolgte. Es konnte in allen Elektroporationen eine deutliche Proteinexpression detektiert und genügend RNA isoliert werden (Tabelle 4.9). Zudem wurde die Qualität der RNA mit Hilfe des Agilent Bioanalyzer überprüft und jeweils ein RIN-Wert von über 8 erzielt, was auf eine sehr gute RNA-Qualität hinweist. 97
106 4 Ergebnisse Tabelle 4.9 Übersicht der generierten Bibliotheken für das Deep Sequencing. Insgesamt wurden vier Bibliotheken generiert, welche GFP bzw. HIV-1-Proteine exprimierten. Die isolierte RNA war ausreichend konzentriert und von guter Qualität. Probe Expression des Konzentration der Gesamtmenge an Proteins [%] isolierten RNA [ng/µl] RNA [µg] RIN GFP ,5 9,6 9, Nef ,5 10,7 8, Nef ,2 9, Gag ,9 Das Deep Sequencing konnte für alle Proben erfolgreich durchgeführt werden. In drei von vier Proben wurden mehr als 15 Mio. Sequenzen detektiert und die Mehrzahl war Nukleotide lang (Abbildung 4.44), so dass es sich dabei um mirnas handelt. Diese wurden in jeder Probe nach ihrer Häufigkeit sortiert und anschließend wurden die Daten mit der Datenbank des miranalyzers abgeglichen, um die erhaltenen Sequenzen bekannten mirnas zuzuordnen. Die Anzahl der erhaltenen Sequenzen für jede mirna wurde prozentual auf die insgesamt detektierten mirna-sequenzen umgerechnet. Daraufhin wurden in jeder Probe nur mirnas weiter analysiert, bei welchen die Anzahl der gelesenen Sequenzen über 0,05% der insgesamt erhaltenen mirna-sequenzen liegt. Die Elektroporation mit GFP mrna diente als Kontrolle, so dass die Expression der verschiedenen mirnas in den HIV-1 mrna-elektroporierten CD4 + T-Zellen mit der Expression in den GFP mrna-elektroporierten Zellen verglichen wurde. Somit konnte eine Zu- oder Abnahme der Expression von einigen mirnas nach Elektroporation mit HIV-1 mrna im Vergleich zu GFP gemessen werden, wobei die Unterschiede sehr gering waren. Es wurden 14 mirnas detektiert, welche ihre Expression nach Elektroporation mit HIV-1 mrna im Vergleich zu GFP mrna modifizieren. Allerdings induzierten alle drei verschiedenen HIV-1 mrnas Veränderungen in die gleiche Richtung (Tabelle 4.10 A). Hierbei könnte es sich um mirnas handeln, die eine allgemeine Rolle in der viralen Immunantwort spielen und das zelluläre mirna-profil in ähnlicher Weise manipulieren. Ein verändertes Expressionsprofil der HIV-1 mrna-elektroporierten CD4 + T-Zellen im Vergleich zu GFP mrna-elektroporierten T-Zellen wurde bei 20 weiteren mirnas nachgewiesen, jedoch gingen die Expressionsunterschiede der mirnas zwischen den diversen HIV-1-Genen in unterschiedliche Richtungen (Tabelle 4.10 B). In den meisten Fällen resultierte die Elektroporation mit 0284 nef mrna in einer verringerten Expression der jeweiligen mirna im Vergleich zur Elektroporation mit GFP mrna, wohingegen 0225 nef oder 0225 gag mrna zu einer erhöhten Expression führten oder keinen Effekt zeigten. Dies würde darauf hinweisen, dass HIV-1-Gene aus Patienten mit unterschiedlichem Krankheitsverlauf das mirna-profil divergent beeinflussen können. Die Ergebnisse sind in Tabelle 4.10 A und B zusammengefasst. 98
107 4 Ergebnisse Abbildung 4.44 Darstellung der erhaltenen Sequenzgrößen (x-achse) gegen ihre Häufigkeit in der Gesamtzahl (y-achse) beim Deep Sequencing der vier Bibliotheken. Die Mehrzahl der sequenzierten Fragmente ist zwischen 18 und 25 Nukleotide lang, was den mirnas entspricht. Des Weiteren sind 5% der Sequenzen ca. fünf Nukleotide lang, wobei es sich um degradierte Fragmente handelt. Zudem wurden auch Fragmente mit einer Länge von 28 bis 30 Nukleotiden sequenziert, die den pirnas (piwi interacting RNA) entsprechen. GWO-1: GFP mrna, GWO-2: 0225 nef mrna, GWO-3: 0225 gag mrna, GWO-4: 0284 nef mrna Damit enthüllte die Analyse von CD4 + T-Zellen, welche mit HIV-1 mrna elektroporiert wurden, dass einzelne HIV-1-Gene das zelluläre mirna-profil modifizieren können. Das mirna-muster der Zelle wurde jedoch nur geringfügig verändert. 99
108 4 Ergebnisse Tabelle 4.10 Veränderungen der Expression von zellulären mirnas nach Elektroporation mit HIV-1- spezifischen mrnas. Ein Wert größer 1 bedeutet, dass diese mirna in der jeweiligen Probe im Vergleich zu den GFP mrna-elektroporierten Zellen überexprimiert ist. Ein Wert kleiner 1 weist auf eine Abnahme der Expression hin. Dargestellt sind nur mirnas mit Werten größer 1,1 oder kleiner 0,9. Die Bezeichnung -3p und -5p gibt den betreffenden Strang der mirna an. A: Die Modifikation der Expression dieser 14 mirnas geht bei allen Proben in die gleiche Richtung. B: Die Modifikation der Expression dieser mirnas ist zwischen den Proben unterschiedlich. Die Probe, welche im Vergleich zu den beiden anderen Proben eine andere Modifikation aufweist, ist grau hervorgehoben. In den meisten Fällen handelt es sich dabei um die Zellen, welche mit 0284 nef mrna elektroporiert wurden. A B mirna 0225 Gag 0225 Nef 0284 Nef let-7a 1,27 1,28 1,38 let-7b 1,41 1,39 1,42 let-7d 1,15 1,27 1,27 let-7f 1,38 1,39 1,33 let-7i 1,13 1,19 1,15 mir-10a-5p 1,16 1,19 1,07 mir-30e-3p 1,24 1,25 1,15 mir-103a-3p 1,14 1,15 1,08 mir-125a-5p 0,86 0,80 1,06 mir-146a-5p 0,79 0,79 0,74 mir-146b-5p 0,81 0,77 0,74 mir-148a-3p 1,14 1,15 1,05 mir-425-5p 0,76 0,77 0,78 mir ,93 0,79 0,72 mirna 0225 Gag 0225 Nef 0284 Nef mir-19b-3p 1,13 1,06 0,82 mir-21-3p 1,03 1,07 0,81 mir-21-5p 1,08 1,09 0,85 mir-24-3p 0,97 0,84 0,89 mir-26a-5p 1,18 1,17 1,02 mir-26b-5p 1,44 1,22 0,98 mir-27a-3p 1,23 1,26 0,87 mir-29a-3p 1,05 1,09 0,87 mir-29c-3p 1,09 1,09 0,80 mir-30c-5p 0,89 0,94 1,12 mir-92a-3p 0,98 1,01 1,20 mir-92b-3p 0,96 1,07 1,41 mir-99b-5p 1,07 0,95 1,20 mir-101-3p 1,06 1,11 0,84 mir-141-3p 1,01 1,01 0,76 mir-142-3p 1,08 1,09 0,62 mir-142-5p 1,12 1,12 0,81 mir-150-5p 0,97 0,99 1,36 mir-342-3p 1,21 1,11 1,04 mir-342-5p 0,83 0,94 1,07 mir-361-3p 0,97 0,86 0,77 mir-423-3p 0,79 0,90 0,98 mir-423-5p 0,93 1,10 1,37 mir p 1,12 0,92 0,59 100
109 4 Ergebnisse Durch HIV-1-Proteine modifizierte mirnas konnten teilweise in einzelnen Proben durch qrt-pcr validiert werden Zunächst wurden zu den 34 mirnas, die eine veränderte Expressionsstärke durch einzelne HIV-1-Proteine zeigten, bekannte Funktionen und Krankheitsbilder recherchiert. Dabei wurden mirnas identifiziert, für welche bereits eine Rolle in der HIV-1-Replikation beschrieben wurde (z.b. mir-30, -125, -150) (155, 156). Ebenso wurden mirnas gefunden, die eine Rolle bei anderen viralen Infektionen haben (mir-27a, -141) (157, 158), die eine wichtige immunologische Funktion zu haben scheinen (z.b. let-7-familie, mir-26a, -142, -146a, -148a) ( ) und für die noch keine Funktion bekannt ist (z.b. mir-1307). Von diesen wurden nun interessante Kandidaten ausgewählt, deren Expression in quantitativer Real time-pcr (qrt-pcr) validiert werden sollte. Die Daten sollten sowohl in Aliquots der RNA, die für das Deep Sequencing gewonnen wurde, als auch in RNA-Proben eines zweiten Spenders erhoben werden. Die RNA des zweiten, gesunden Spenders wurde nach demselben Verfahren gewonnen wie die RNA der Deep Sequencing Proben. Die Expression der HIV-1-Proteine 24 Stunden nach Elektroporation war außer für 0225 Nef bei Spender 2 höher als bei Spender 1 (GFP: 92%, 0225 Gag: 82%, 0225 Nef: 35%, 0284 Nef: 21%; Vergleiche dazu Tabelle 4.9). Aus den gewonnenen RNA-Proben wurde zunächst spezifische cdna zu jeder mirna synthetisiert, welche anschließend als Matrize in der qrt-pcr diente. Die Analyse fand in Triplikaten statt. Die kleinen, nukleären RNAs RNU44 und RNU6B dienten als endogene Kontrolle für die quantitativen mirna-expressions-levels. Für alle ausgewählten mirnas konnte die qrt-pcr erfolgreich durchgeführt werden. Die Triplikate zeigten kaum Abweichungen voneinander. Der Vergleich der mirna- Expression bezog sich auf die RNA-Proben der GFP mrna-elektroporierten CD4 + T-Zellen, so dass diese auf den Wert 1 gesetzt wurden. Die Ergebnisse sind in Abbildung 4.45 dargestellt. Es wurde deutlich, dass sich die Resultate der Deep Sequencing- Analyse nur bei einigen, wenigen mirnas bestätigen ließen. So konnten die Expressionsveränderungen von let-7b, mir-148a, mir-150 und mir-342 nicht verifiziert werden. Die Beobachtungen der Deep Sequencing-Analyse von mir-26b, p und konnten nur in einem Spender bestätigt werden (Abbildung 4.45 C, G und L). Die Ergebnisse der restlichen mirnas konnten zumindest in einzelnen Proben in beiden Spendern validiert werden. So wurde eine Expressionszunahme von let-7a im Deep Sequencing nach Elektroporation der drei HIV-1-spezifischen mrnas detektiert. In der qrt-pcr konnte dieser Anstieg für die Probe 0225 Gag in beiden Spendern und für die beiden Nef-Proben in jeweils nur einem Spender bestätigt werden (Abbildung 4.45 A). Ähnlich verhielt es sich bei let-7f, mir-27a, mir-30c und mir-146a (Abbildung 4.45 B, D, 101
.")
110 4 Ergebnisse E und H). In drei von diesen vier mirnas konnte stets das Resultat in der Probe 0284 Nef validiert werden. Für eine mirnas konnte das Ergebnis der Deep Sequencing-Analyse vollständig bestätigt werden. Das Nef-Protein von 0284 führte in beiden Spendern zu einer Reduktion der mir-142-3p (Abbildung 4.45 F). Insgesamt waren die Differenzen von geringem Ausmaß, was mit den Beobachtungen der Deep Sequencing-Analyse übereinstimmte. Somit kann festgehalten werden, dass nicht alle gemessenen mirna- Expressionsunterschiede im Deep Sequencing durch qrt-pcr validiert werden konnten. Dennoch zeigten einige mirnas durch HIV-1-spezifische mrnas eine Veränderung ihrer Expression, was zudem in zwei unterschiedlichen Spendern bestätigt werden konnte. Diese stellen interessante Kandidaten für weitere Analysen dar. 102
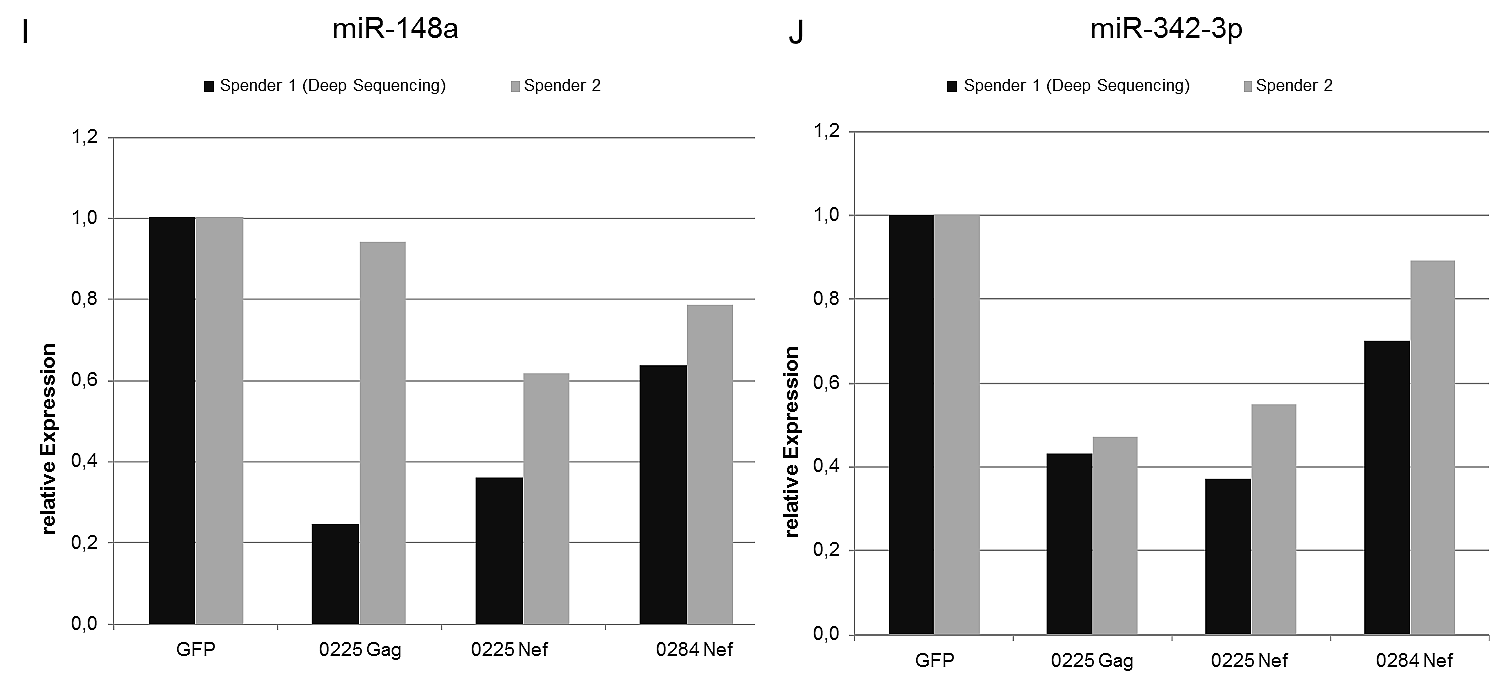
111 103 4 Ergebnisse

112 4 Ergebnisse Abbildung 4.45 Die Expression verschiedener zellulärer mirnas wird durch HIV-1-Gene beeinflusst, jedoch wurden nur einzelne Proben durch qrt-pcr validiert. CD4 + T-Zellen zweier gesunder Spender wurden mit GFP mrna, 0225 gag, 0225 nef und 0284 nef mrna elektroporiert. Nach 24 Stunden wurden die Zellen geerntet und RNA isoliert. Nach spezifischer cdna-synthese für die jeweiligen mirnas wurde eine qrt-pcr durchgeführt. Die Auswertung erfolgte nach der ΔΔC T-Methode, RNU6B und RNU44 dienten als endogene Kontrolle. Schwarze Säulen stellen das Ergebnis der Analyse mit RNA-Proben des Spenders 1 dar, welche auch zur Deep Sequencing-Analyse verwendet wurden. In grau sind die Ergebnisse mit RNA-Proben des Spender 2 dargestellt. Resultate des Deep Sequencing, die durch qrt-pcr bestätigt werden konnten, sind orange umrandet, wobei eine durchgezogene Linie anzeigt, dass es in beiden Spendern reproduziert werden konnte. Eine gestrichelte Linie weist darauf hin, dass es nur in einem der beiden Spender validiert wurde CTL Fluchtmutationen in mirna-binderegionen beeinflussen die Regulation von mirnas Da das HI-Virus aufgrund seiner hohen Sequenzdiversität sehr variabel ist, kann es der Inhibierung mittels mirnas durch Mutationen in der Zielsequenz der mirna entkommen (65). In erster Linie tragen Fluchtmutationen in CTL-Epitopen zur Variabilität bei. So liegt die Bindesequenz der humanen mir-29a, welche bekannter Weise die Nef-Expression reprimiert und die Virusreplikation hemmt, im offenen Leserahmen des nef Gens (71). In dieser Zielsequenz befindet sich auch das bekannte HLA-A*24-restringierte CTL-Epitop RY10 und das HLA-B*18-restringierte CTL-Epitop YY9 (129, 153). Ob Fluchtmutationen in diesem CTL-Epitop einen Einfluss auf die Bindungsaffinität der mirna haben, sollte nun untersucht werden. Dazu wurde der offene Leserahmen des nef Gens von SF2 und von vier autologen Patientenisolaten als 3`UTR hinter die cdna-sequenz des Luziferasegens in den Vektor psicheck3 kloniert. Anschließend wurden die verschiedenen Plasmide zusammen mit einem mir-29a-expressionsplasmid in HEK293 transfiziert. Nach 48 Stunden wurde ein dualer Luziferase-Reporter-Assay mit den Zellen durchgeführt, bei welchem die Aktivität der Firefly-Luziferase gemessen wurde. Abbildung 4.46 zeigt die 104
AUFGABENSAMMLUNG Lösungen. Variabilität von Antikörpern 1
 Variabilität von Antikörpern 1 Rezeptoren bzw. Antikörper eines noch undifferenzierten B-Lymphocyten: a) Schreiben Sie die Anzahl der variablen Exons je Chromosom auf. b) Berechnen Sie die mögliche Anzahl
Variabilität von Antikörpern 1 Rezeptoren bzw. Antikörper eines noch undifferenzierten B-Lymphocyten: a) Schreiben Sie die Anzahl der variablen Exons je Chromosom auf. b) Berechnen Sie die mögliche Anzahl
Lineargleichungssysteme: Additions-/ Subtraktionsverfahren
 Lineargleichungssysteme: Additions-/ Subtraktionsverfahren W. Kippels 22. Februar 2014 Inhaltsverzeichnis 1 Einleitung 2 2 Lineargleichungssysteme zweiten Grades 2 3 Lineargleichungssysteme höheren als
Lineargleichungssysteme: Additions-/ Subtraktionsverfahren W. Kippels 22. Februar 2014 Inhaltsverzeichnis 1 Einleitung 2 2 Lineargleichungssysteme zweiten Grades 2 3 Lineargleichungssysteme höheren als
Bekommen durch Ansteckung. H Human Beim Menschen. Acquired I D. Schwäche des Immunsystems. Schwäche des Immunsystems.
 Test 3.1.1 HIV / Aids: Was ist das? H Human Beim Menschen A Acquired Bekommen durch Ansteckung I Immunodeficiency Schwäche des Immunsystems I D Immune Deficiency Schwäche des Immunsystems V Virus Virus
Test 3.1.1 HIV / Aids: Was ist das? H Human Beim Menschen A Acquired Bekommen durch Ansteckung I Immunodeficiency Schwäche des Immunsystems I D Immune Deficiency Schwäche des Immunsystems V Virus Virus
Klonierung von S2P Rolle der M19-Zellen. POL-Seminar der Biochemie II 13.02.2007 Sebastian Gabriel
 Klonierung von S2P Rolle der M19-Zellen POL-Seminar der Biochemie II 13.02.2007 Sebastian Gabriel Inhalt 1. Was ist eine humane genomische DNA-Bank? 2. Unterschied zwischen cdna-bank und genomischer DNA-Bank?
Klonierung von S2P Rolle der M19-Zellen POL-Seminar der Biochemie II 13.02.2007 Sebastian Gabriel Inhalt 1. Was ist eine humane genomische DNA-Bank? 2. Unterschied zwischen cdna-bank und genomischer DNA-Bank?
HIV-Infektion und AIDS. Seminar aus funktioneller Pathologie
 HIV-Infektion und AIDS Seminar aus funktioneller Pathologie Retroviren des Menschen Lentiviren von Primaten Das Virion des HI-Virus schematisch und elektronenmikroskopisch Virale Gene Bindungssequenzen
HIV-Infektion und AIDS Seminar aus funktioneller Pathologie Retroviren des Menschen Lentiviren von Primaten Das Virion des HI-Virus schematisch und elektronenmikroskopisch Virale Gene Bindungssequenzen
VORANGEGANGENE MODELLE
 VORANGEGANGENE MODELLE UNSER THEMA HEUTE Ziel dieses Papers: - Nähere Charakterisierung der AuxREs - Analyse eines zu den AuxREs gehörenden Transkriptionsfaktors WAS BEREITS BEKANNT WAR: - Auxin moduliert
VORANGEGANGENE MODELLE UNSER THEMA HEUTE Ziel dieses Papers: - Nähere Charakterisierung der AuxREs - Analyse eines zu den AuxREs gehörenden Transkriptionsfaktors WAS BEREITS BEKANNT WAR: - Auxin moduliert
Übung 11 Genregulation bei Prokaryoten
 Übung 11 Genregulation bei Prokaryoten Konzepte: Differentielle Genexpression Positive Genregulation Negative Genregulation cis-/trans-regulation 1. Auf welchen Ebenen kann Genregulation stattfinden? Definition
Übung 11 Genregulation bei Prokaryoten Konzepte: Differentielle Genexpression Positive Genregulation Negative Genregulation cis-/trans-regulation 1. Auf welchen Ebenen kann Genregulation stattfinden? Definition
Cytomegalie & Co. Häufige Virusinfektionen in der Schwangerschaft. Deutsches Grünes Kreuz e.v.
 Cytomegalie & Co Häufige Virusinfektionen in der Schwangerschaft Schwangerschaft Eine ganz besondere Zeit der Vorfreude Verantwortung Sorge Die werdenden Eltern möchten alles richtig machen für das Wohl
Cytomegalie & Co Häufige Virusinfektionen in der Schwangerschaft Schwangerschaft Eine ganz besondere Zeit der Vorfreude Verantwortung Sorge Die werdenden Eltern möchten alles richtig machen für das Wohl
Biologie I/B: Klassische und molekulare Genetik, molekulare Grundlagen der Entwicklung Theoretische Übungen SS 2014
 Biologie I/B: Klassische und molekulare Genetik, molekulare Grundlagen der Entwicklung Theoretische Übungen SS 2014 Fragen für die Übungsstunde 8 (14.07-18.07.) 1) Von der DNA-Sequenz zum Protein Sie können
Biologie I/B: Klassische und molekulare Genetik, molekulare Grundlagen der Entwicklung Theoretische Übungen SS 2014 Fragen für die Übungsstunde 8 (14.07-18.07.) 1) Von der DNA-Sequenz zum Protein Sie können
Übung 8. 1. Zellkommunikation. Vorlesung Bio-Engineering Sommersemester 2008. Kapitel 4. 4
 Bitte schreiben Sie Ihre Antworten direkt auf das Übungsblatt. Falls Sie mehr Platz brauchen verweisen Sie auf Zusatzblätter. Vergessen Sie Ihren Namen nicht! Abgabe der Übung bis spätestens 05. 05. 08
Bitte schreiben Sie Ihre Antworten direkt auf das Übungsblatt. Falls Sie mehr Platz brauchen verweisen Sie auf Zusatzblätter. Vergessen Sie Ihren Namen nicht! Abgabe der Übung bis spätestens 05. 05. 08
Ein Einfaches AIDS Modell
 Ein Einfaches AIDS Modell Martin Bauer: 990395 Guntram Rümmele: 99008 Das SIR - Modell Die Modellierung von epidemischen Modellen hat schon lange Tradition. Man hat schon immer versucht Erklärungen für
Ein Einfaches AIDS Modell Martin Bauer: 990395 Guntram Rümmele: 99008 Das SIR - Modell Die Modellierung von epidemischen Modellen hat schon lange Tradition. Man hat schon immer versucht Erklärungen für
Minimale Resterkrankung, Hochdurchsatz-Sequenzierung und Prognose beim multiplen Myelom
 Minimale Resterkrankung, Hochdurchsatz-Sequenzierung und Prognose beim multiplen Myelom Von Navneet Ramesh und Maike Haehle, übersetzt von Sabine Schock Vom 8. Apr 2014 7:41 Uhr; aktualisiert am 8. Apr
Minimale Resterkrankung, Hochdurchsatz-Sequenzierung und Prognose beim multiplen Myelom Von Navneet Ramesh und Maike Haehle, übersetzt von Sabine Schock Vom 8. Apr 2014 7:41 Uhr; aktualisiert am 8. Apr
Influenza des Schweines
 Influenza des Schweines Historie: - erstmals beobachtet 1918 in USA, China, Ungarn - zeitgleich mit der Pandemie beim Menschen (> 20 Mill. Tote) - Virus-Subtyp H1N1 - Übertragung sehr wahrscheinlich vom
Influenza des Schweines Historie: - erstmals beobachtet 1918 in USA, China, Ungarn - zeitgleich mit der Pandemie beim Menschen (> 20 Mill. Tote) - Virus-Subtyp H1N1 - Übertragung sehr wahrscheinlich vom
Würfelt man dabei je genau 10 - mal eine 1, 2, 3, 4, 5 und 6, so beträgt die Anzahl. der verschiedenen Reihenfolgen, in denen man dies tun kann, 60!.
 040304 Übung 9a Analysis, Abschnitt 4, Folie 8 Die Wahrscheinlichkeit, dass bei n - maliger Durchführung eines Zufallexperiments ein Ereignis A ( mit Wahrscheinlichkeit p p ( A ) ) für eine beliebige Anzahl
040304 Übung 9a Analysis, Abschnitt 4, Folie 8 Die Wahrscheinlichkeit, dass bei n - maliger Durchführung eines Zufallexperiments ein Ereignis A ( mit Wahrscheinlichkeit p p ( A ) ) für eine beliebige Anzahl
RSV. RSV kennen. Das Virus, das Eltern kennen sollten. Informationen. Kinder schützen
 RSV kennen Kinder schützen RSV Das Virus, das Eltern kennen sollten. Informationen zu einem wenig bekannten Virus sowie praktische Ratschläge, wie Sie das Risiko einer RSV-Infektion verringern können.
RSV kennen Kinder schützen RSV Das Virus, das Eltern kennen sollten. Informationen zu einem wenig bekannten Virus sowie praktische Ratschläge, wie Sie das Risiko einer RSV-Infektion verringern können.
Text: Octapharma GmbH Illustrationen: Günter Hengsberg Layout: nonmodo, Köln
 Unser Immunsystem Text: Octapharma GmbH Illustrationen: Günter Hengsberg Layout: nonmodo, Köln Bakterien und Viren können uns krank machen. Wir bekommen dann Husten, Schnupfen oder Durchfall. Unser Körper
Unser Immunsystem Text: Octapharma GmbH Illustrationen: Günter Hengsberg Layout: nonmodo, Köln Bakterien und Viren können uns krank machen. Wir bekommen dann Husten, Schnupfen oder Durchfall. Unser Körper
Versuch 8. Plasmid - Isolierung
 Versuch 8 Plasmid - Isolierung Protokollant: E-mail: Studiengang: Gruppen-Nr: Semester: Betreuer: Max Mustermann max@quantentunnel.de X X X C. Weindel & M. Schwarz Wird benotet?: Einleitung Ein Plasmid
Versuch 8 Plasmid - Isolierung Protokollant: E-mail: Studiengang: Gruppen-Nr: Semester: Betreuer: Max Mustermann max@quantentunnel.de X X X C. Weindel & M. Schwarz Wird benotet?: Einleitung Ein Plasmid
Gefährlich hohe Blutzuckerwerte
 Gefährlich hohe Blutzuckerwerte In besonderen Situationen kann der Blutzuckerspiegel gefährlich ansteigen Wenn in Ausnahmefällen der Blutzuckerspiegel sehr hoch ansteigt, dann kann das für Sie gefährlich
Gefährlich hohe Blutzuckerwerte In besonderen Situationen kann der Blutzuckerspiegel gefährlich ansteigen Wenn in Ausnahmefällen der Blutzuckerspiegel sehr hoch ansteigt, dann kann das für Sie gefährlich
6. DNA -Bakteriengenetik
 6. DNA -Bakteriengenetik Konzepte: Francis Crick DNA Struktur DNA Replikation Gentransfer in Bakterien Bakteriophagen 2. Welcher der folgenden Sätze entspricht der Chargaff-Regel? A) Die Menge von Purinen
6. DNA -Bakteriengenetik Konzepte: Francis Crick DNA Struktur DNA Replikation Gentransfer in Bakterien Bakteriophagen 2. Welcher der folgenden Sätze entspricht der Chargaff-Regel? A) Die Menge von Purinen
Grundideen der Gentechnik
 Grundideen der Gentechnik Die Gentechnik kombiniert Biotechnik und Züchtung. Wie in der Züchtung wird die Erbinformation eines Lebewesen verändert. Dabei nutzte man in den Anfängen der Gentechnik vor allem
Grundideen der Gentechnik Die Gentechnik kombiniert Biotechnik und Züchtung. Wie in der Züchtung wird die Erbinformation eines Lebewesen verändert. Dabei nutzte man in den Anfängen der Gentechnik vor allem
Patienteninformation: Gentestung bei familiärem Brust- und Eierstockkrebs (Basis-Information):
:") Frauenklinik Gynäkologie und Gynäkologische Onkologie Patienteninformation: Gentestung bei familiärem Brust- und Eierstockkrebs (Basis-Information): Universitätsspital Basel Frauenklinik PD Dr. med. Nicole
Frauenklinik Gynäkologie und Gynäkologische Onkologie Patienteninformation: Gentestung bei familiärem Brust- und Eierstockkrebs (Basis-Information): Universitätsspital Basel Frauenklinik PD Dr. med. Nicole
Epstein-Barr-Virus-Infektion: Möglichkeiten und Grenzen der serologischen Diagnostik von Reaktivierungen und chronischen Verläufen
 Epstein-Barr-Virus-Infektion: Möglichkeiten und Grenzen der serologischen Diagnostik von Reaktivierungen und chronischen Verläufen Dr. Claudia Wolff Viramed Biotech AG 1 Akute EBV-Primärinfektion Erster
Epstein-Barr-Virus-Infektion: Möglichkeiten und Grenzen der serologischen Diagnostik von Reaktivierungen und chronischen Verläufen Dr. Claudia Wolff Viramed Biotech AG 1 Akute EBV-Primärinfektion Erster
Kulturelle Evolution 12
 3.3 Kulturelle Evolution Kulturelle Evolution Kulturelle Evolution 12 Seit die Menschen Erfindungen machen wie z.b. das Rad oder den Pflug, haben sie sich im Körperbau kaum mehr verändert. Dafür war einfach
3.3 Kulturelle Evolution Kulturelle Evolution Kulturelle Evolution 12 Seit die Menschen Erfindungen machen wie z.b. das Rad oder den Pflug, haben sie sich im Körperbau kaum mehr verändert. Dafür war einfach
Musterlösung - Übung 5 Vorlesung Bio-Engineering Sommersemester 2008
 Aufgabe 1: Prinzipieller Ablauf der Proteinbiosynthese a) Erklären Sie folgende Begriffe möglichst in Ihren eigenen Worten (1 kurzer Satz): Gen Nukleotid RNA-Polymerase Promotor Codon Anti-Codon Stop-Codon
Aufgabe 1: Prinzipieller Ablauf der Proteinbiosynthese a) Erklären Sie folgende Begriffe möglichst in Ihren eigenen Worten (1 kurzer Satz): Gen Nukleotid RNA-Polymerase Promotor Codon Anti-Codon Stop-Codon
Pädagogik. Melanie Schewtschenko. Eingewöhnung und Übergang in die Kinderkrippe. Warum ist die Beteiligung der Eltern so wichtig?
 Pädagogik Melanie Schewtschenko Eingewöhnung und Übergang in die Kinderkrippe Warum ist die Beteiligung der Eltern so wichtig? Studienarbeit Inhaltsverzeichnis 1. Einleitung.2 2. Warum ist Eingewöhnung
Pädagogik Melanie Schewtschenko Eingewöhnung und Übergang in die Kinderkrippe Warum ist die Beteiligung der Eltern so wichtig? Studienarbeit Inhaltsverzeichnis 1. Einleitung.2 2. Warum ist Eingewöhnung
Herpes simplex Infektionen bei HIV - infizierten Patienten. Johannes R. Bogner Uni München
 Herpes simplex Infektionen bei HIV - infizierten Patienten Johannes R. Bogner Uni München 1. Die klinische Präsentation von HSV 1 und HSV 2 Läsionen unterscheidet sich oft von Läsionen bei immunkompetenten
Herpes simplex Infektionen bei HIV - infizierten Patienten Johannes R. Bogner Uni München 1. Die klinische Präsentation von HSV 1 und HSV 2 Läsionen unterscheidet sich oft von Läsionen bei immunkompetenten
Herstellung und Selektion rekombinanter Antikörper
 Herstellung und Selektion rekombinanter Antikörper Für jeden Topf ein Deckel Sintox 16.04.2015 16.04.15 Sintox 1 Inhalte Begriffsklärungen Rekombinant Lymphozyten Antikörper und Antigene Somatische Hypermutation
Herstellung und Selektion rekombinanter Antikörper Für jeden Topf ein Deckel Sintox 16.04.2015 16.04.15 Sintox 1 Inhalte Begriffsklärungen Rekombinant Lymphozyten Antikörper und Antigene Somatische Hypermutation
L10N-Manager 3. Netzwerktreffen der Hochschulübersetzer/i nnen Mannheim 10. Mai 2016
 L10N-Manager 3. Netzwerktreffen der Hochschulübersetzer/i nnen Mannheim 10. Mai 2016 Referentin: Dr. Kelly Neudorfer Universität Hohenheim Was wir jetzt besprechen werden ist eine Frage, mit denen viele
L10N-Manager 3. Netzwerktreffen der Hochschulübersetzer/i nnen Mannheim 10. Mai 2016 Referentin: Dr. Kelly Neudorfer Universität Hohenheim Was wir jetzt besprechen werden ist eine Frage, mit denen viele
Osteoporose. Ein echtes Volksleiden. Schon jetzt zählen die Osteoporose und die damit verbundene erhöhte Brüchigkeit der Knochen
 Osteoporose Osteoporose 9 Osteoporose Ein echtes Volksleiden Schon jetzt zählen die Osteoporose und die damit verbundene erhöhte Brüchigkeit der Knochen in den entwickelten Ländern zu den häufigsten Erkrankungen
Osteoporose Osteoporose 9 Osteoporose Ein echtes Volksleiden Schon jetzt zählen die Osteoporose und die damit verbundene erhöhte Brüchigkeit der Knochen in den entwickelten Ländern zu den häufigsten Erkrankungen
Klausur zur Vorlesung Biochemie III im WS 2000/01
 Klausur zur Vorlesung Biochemie III im WS 2000/01 am 15.02.2001 von 15.30 17.00 Uhr (insgesamt 100 Punkte, mindestens 40 erforderlich) Bitte Name, Matrikelnummer und Studienfach unbedingt angeben (3 1.
Klausur zur Vorlesung Biochemie III im WS 2000/01 am 15.02.2001 von 15.30 17.00 Uhr (insgesamt 100 Punkte, mindestens 40 erforderlich) Bitte Name, Matrikelnummer und Studienfach unbedingt angeben (3 1.
1. Einführung 2. 2. Erstellung einer Teillieferung 2. 3. Erstellung einer Teilrechnung 6
 Inhalt 1. Einführung 2 2. Erstellung einer Teillieferung 2 3. Erstellung einer Teilrechnung 6 4. Erstellung einer Sammellieferung/ Mehrere Aufträge zu einem Lieferschein zusammenfassen 11 5. Besonderheiten
Inhalt 1. Einführung 2 2. Erstellung einer Teillieferung 2 3. Erstellung einer Teilrechnung 6 4. Erstellung einer Sammellieferung/ Mehrere Aufträge zu einem Lieferschein zusammenfassen 11 5. Besonderheiten
Erfolg im Verkauf durch Persönlichkeit! Potenzialanalyse, Training & Entwicklung für Vertriebsmitarbeiter!
 Wer in Kontakt ist verkauft! Wie reden Sie mit mir? Erfolg im Verkauf durch Persönlichkeit! Potenzialanalyse, Training & Entwicklung für Vertriebsmitarbeiter! www.sizeprozess.at Fritz Zehetner Persönlichkeit
Wer in Kontakt ist verkauft! Wie reden Sie mit mir? Erfolg im Verkauf durch Persönlichkeit! Potenzialanalyse, Training & Entwicklung für Vertriebsmitarbeiter! www.sizeprozess.at Fritz Zehetner Persönlichkeit
AGROPLUS Buchhaltung. Daten-Server und Sicherheitskopie. Version vom 21.10.2013b
 AGROPLUS Buchhaltung Daten-Server und Sicherheitskopie Version vom 21.10.2013b 3a) Der Daten-Server Modus und der Tresor Der Daten-Server ist eine Betriebsart welche dem Nutzer eine grosse Flexibilität
AGROPLUS Buchhaltung Daten-Server und Sicherheitskopie Version vom 21.10.2013b 3a) Der Daten-Server Modus und der Tresor Der Daten-Server ist eine Betriebsart welche dem Nutzer eine grosse Flexibilität
Berechnung der Erhöhung der Durchschnittsprämien
 Wolfram Fischer Berechnung der Erhöhung der Durchschnittsprämien Oktober 2004 1 Zusammenfassung Zur Berechnung der Durchschnittsprämien wird das gesamte gemeldete Prämienvolumen Zusammenfassung durch die
Wolfram Fischer Berechnung der Erhöhung der Durchschnittsprämien Oktober 2004 1 Zusammenfassung Zur Berechnung der Durchschnittsprämien wird das gesamte gemeldete Prämienvolumen Zusammenfassung durch die
Zeichen bei Zahlen entschlüsseln
 Zeichen bei Zahlen entschlüsseln In diesem Kapitel... Verwendung des Zahlenstrahls Absolut richtige Bestimmung von absoluten Werten Operationen bei Zahlen mit Vorzeichen: Addieren, Subtrahieren, Multiplizieren
Zeichen bei Zahlen entschlüsseln In diesem Kapitel... Verwendung des Zahlenstrahls Absolut richtige Bestimmung von absoluten Werten Operationen bei Zahlen mit Vorzeichen: Addieren, Subtrahieren, Multiplizieren
Schnell reagieren schneller wieder fit.
 Wehr Dich gegen die Erkältungs- Viren! Schnell reagieren schneller wieder fit. Clevere Taktik: Viren ausschalten, ehe die Erkältung richtig zuschlägt Ganz egal, ob es gerade erst im Hals kratzt oder die
Wehr Dich gegen die Erkältungs- Viren! Schnell reagieren schneller wieder fit. Clevere Taktik: Viren ausschalten, ehe die Erkältung richtig zuschlägt Ganz egal, ob es gerade erst im Hals kratzt oder die
Die HIT ist keine Allergie! Da die von ihr ausgelösten. Krankheitsbild. Was ist eine Histamin- Intoleranz?
 Was ist eine Histamin- Intoleranz? Die Histamin-Intoleranz ist eine Pseudoallergie. Die HIT ist keine Allergie! Da die von ihr ausgelösten Gesundheitsstörungen jedoch von allergiebedingten Beschwerden
Was ist eine Histamin- Intoleranz? Die Histamin-Intoleranz ist eine Pseudoallergie. Die HIT ist keine Allergie! Da die von ihr ausgelösten Gesundheitsstörungen jedoch von allergiebedingten Beschwerden
Der HIV-Antikörper-Schnelltest aus Sicht des Labormediziners. Dr. Thomas Berg, Berlin www.bergdoctor.de
 Der HIV-Antikörper-Schnelltest aus Sicht des Labormediziners Was untersucht der HIV-Antikörper- Schnelltest? (am Beispiel Vitest HIV) Der HIV-Antikörper-Schnelltest ist ein SUCHTEST, der untersucht, ob
Der HIV-Antikörper-Schnelltest aus Sicht des Labormediziners Was untersucht der HIV-Antikörper- Schnelltest? (am Beispiel Vitest HIV) Der HIV-Antikörper-Schnelltest ist ein SUCHTEST, der untersucht, ob
Primzahlen und RSA-Verschlüsselung
 Primzahlen und RSA-Verschlüsselung Michael Fütterer und Jonathan Zachhuber 1 Einiges zu Primzahlen Ein paar Definitionen: Wir bezeichnen mit Z die Menge der positiven und negativen ganzen Zahlen, also
Primzahlen und RSA-Verschlüsselung Michael Fütterer und Jonathan Zachhuber 1 Einiges zu Primzahlen Ein paar Definitionen: Wir bezeichnen mit Z die Menge der positiven und negativen ganzen Zahlen, also
Repetitionsaufgaben Wurzelgleichungen
 Repetitionsaufgaben Wurzelgleichungen Inhaltsverzeichnis A) Vorbemerkungen B) Lernziele C) Theorie mit Aufgaben D) Aufgaben mit Musterlösungen 4 A) Vorbemerkungen Bitte beachten Sie: Bei Wurzelgleichungen
Repetitionsaufgaben Wurzelgleichungen Inhaltsverzeichnis A) Vorbemerkungen B) Lernziele C) Theorie mit Aufgaben D) Aufgaben mit Musterlösungen 4 A) Vorbemerkungen Bitte beachten Sie: Bei Wurzelgleichungen
2.8 Grenzflächeneffekte
 - 86-2.8 Grenzflächeneffekte 2.8.1 Oberflächenspannung An Grenzflächen treten besondere Effekte auf, welche im Volumen nicht beobachtbar sind. Die molekulare Grundlage dafür sind Kohäsionskräfte, d.h.
- 86-2.8 Grenzflächeneffekte 2.8.1 Oberflächenspannung An Grenzflächen treten besondere Effekte auf, welche im Volumen nicht beobachtbar sind. Die molekulare Grundlage dafür sind Kohäsionskräfte, d.h.
Transgene Organismen
 Transgene Organismen Themenübersicht 1) Einführung 2) Komplementäre DNA (cdna) 3) Vektoren 4) Einschleusung von Genen in Eukaryontenzellen 5) Ausmaß der Genexpression 6) Genausschaltung (Gen-Knockout)
Transgene Organismen Themenübersicht 1) Einführung 2) Komplementäre DNA (cdna) 3) Vektoren 4) Einschleusung von Genen in Eukaryontenzellen 5) Ausmaß der Genexpression 6) Genausschaltung (Gen-Knockout)
Ein neues System für die Allokation von Spenderlungen. LAS Information für Patienten in Deutschland
 Ein neues System für die Allokation von Spenderlungen LAS Information für Patienten in Deutschland Ein neues System für die Allokation von Spenderlungen Aufgrund des immensen Mangels an Spenderorganen
Ein neues System für die Allokation von Spenderlungen LAS Information für Patienten in Deutschland Ein neues System für die Allokation von Spenderlungen Aufgrund des immensen Mangels an Spenderorganen
10.000 10.000. an Masern erkrankt. an Mumps erkrankt. mit Röteln infiziert
 Nutzen: Bei Viruskontakt ist mein geimpftes Kind auch künftig besser gegen Masern, Mumps und Röteln geschützt. Je mehr Menschen geimpft sind, desto unwahrscheinlicher werden Kontakte mit dem Virus. ohne
Nutzen: Bei Viruskontakt ist mein geimpftes Kind auch künftig besser gegen Masern, Mumps und Röteln geschützt. Je mehr Menschen geimpft sind, desto unwahrscheinlicher werden Kontakte mit dem Virus. ohne
13. Lineare DGL höherer Ordnung. Eine DGL heißt von n-ter Ordnung, wenn Ableitungen y, y, y,... bis zur n-ten Ableitung y (n) darin vorkommen.
 darin vorkommen.") 13. Lineare DGL höherer Ordnung. Eine DGL heißt von n-ter Ordnung, wenn Ableitungen y, y, y,... bis zur n-ten Ableitung y (n) darin vorkommen. Sie heißt linear, wenn sie die Form y (n) + a n 1 y (n 1)
13. Lineare DGL höherer Ordnung. Eine DGL heißt von n-ter Ordnung, wenn Ableitungen y, y, y,... bis zur n-ten Ableitung y (n) darin vorkommen. Sie heißt linear, wenn sie die Form y (n) + a n 1 y (n 1)
Professionelle Seminare im Bereich MS-Office
 Der Name BEREICH.VERSCHIEBEN() ist etwas unglücklich gewählt. Man kann mit der Funktion Bereiche zwar verschieben, man kann Bereiche aber auch verkleinern oder vergrößern. Besser wäre es, die Funktion
Der Name BEREICH.VERSCHIEBEN() ist etwas unglücklich gewählt. Man kann mit der Funktion Bereiche zwar verschieben, man kann Bereiche aber auch verkleinern oder vergrößern. Besser wäre es, die Funktion
Insiderwissen 2013. Hintergrund
 Insiderwissen 213 XING EVENTS mit der Eventmanagement-Software für Online Eventregistrierung &Ticketing amiando, hat es sich erneut zur Aufgabe gemacht zu analysieren, wie Eventveranstalter ihre Veranstaltungen
Insiderwissen 213 XING EVENTS mit der Eventmanagement-Software für Online Eventregistrierung &Ticketing amiando, hat es sich erneut zur Aufgabe gemacht zu analysieren, wie Eventveranstalter ihre Veranstaltungen
Lineare Funktionen. 1 Proportionale Funktionen 3 1.1 Definition... 3 1.2 Eigenschaften... 3. 2 Steigungsdreieck 3
 Lineare Funktionen Inhaltsverzeichnis 1 Proportionale Funktionen 3 1.1 Definition............................... 3 1.2 Eigenschaften............................. 3 2 Steigungsdreieck 3 3 Lineare Funktionen
Lineare Funktionen Inhaltsverzeichnis 1 Proportionale Funktionen 3 1.1 Definition............................... 3 1.2 Eigenschaften............................. 3 2 Steigungsdreieck 3 3 Lineare Funktionen
Tutorial about how to use USBView.exe and Connection Optimization for VNWA.
 Tutorial about how to use USBView.exe and Connection Optimization for VNWA. Tutorial über den Gebrauch von USBView.exe und die Anschluss-Optimierung für den VNWA. Es wurde beobachtet, dass bestimmte VNWA
Tutorial about how to use USBView.exe and Connection Optimization for VNWA. Tutorial über den Gebrauch von USBView.exe und die Anschluss-Optimierung für den VNWA. Es wurde beobachtet, dass bestimmte VNWA
«Eine Person ist funktional gesund, wenn sie möglichst kompetent mit einem möglichst gesunden Körper an möglichst normalisierten Lebensbereichen
 18 «Eine Person ist funktional gesund, wenn sie möglichst kompetent mit einem möglichst gesunden Körper an möglichst normalisierten Lebensbereichen teilnimmt und teilhat.» 3Das Konzept der Funktionalen
18 «Eine Person ist funktional gesund, wenn sie möglichst kompetent mit einem möglichst gesunden Körper an möglichst normalisierten Lebensbereichen teilnimmt und teilhat.» 3Das Konzept der Funktionalen
in vivo -- Das Magazin der Deutschen Krebshilfe vom 09.11.2010
 Seite 1/5 in vivo -- Das Magazin der Deutschen Krebshilfe vom 09.11.2010 Expertengespräch zum Thema Retinoblastom Und zu diesem Thema begrüße ich jetzt Professor Norbert Bornfeld, Direktor des Zentrums
Seite 1/5 in vivo -- Das Magazin der Deutschen Krebshilfe vom 09.11.2010 Expertengespräch zum Thema Retinoblastom Und zu diesem Thema begrüße ich jetzt Professor Norbert Bornfeld, Direktor des Zentrums
Bewertung des Blattes
 Bewertung des Blattes Es besteht immer die Schwierigkeit, sein Blatt richtig einzuschätzen. Im folgenden werden einige Anhaltspunkte gegeben. Man unterscheidet: Figurenpunkte Verteilungspunkte Längenpunkte
Bewertung des Blattes Es besteht immer die Schwierigkeit, sein Blatt richtig einzuschätzen. Im folgenden werden einige Anhaltspunkte gegeben. Man unterscheidet: Figurenpunkte Verteilungspunkte Längenpunkte
Konzentration auf das. Wesentliche.
 Konzentration auf das Wesentliche. Machen Sie Ihre Kanzleiarbeit effizienter. 2 Sehr geehrte Leserin, sehr geehrter Leser, die Grundlagen Ihres Erfolges als Rechtsanwalt sind Ihre Expertise und Ihre Mandantenorientierung.
Konzentration auf das Wesentliche. Machen Sie Ihre Kanzleiarbeit effizienter. 2 Sehr geehrte Leserin, sehr geehrter Leser, die Grundlagen Ihres Erfolges als Rechtsanwalt sind Ihre Expertise und Ihre Mandantenorientierung.
Projektmanagement in der Spieleentwicklung
 Projektmanagement in der Spieleentwicklung Inhalt 1. Warum brauche ich ein Projekt-Management? 2. Die Charaktere des Projektmanagement - Mastermind - Producer - Projektleiter 3. Schnittstellen definieren
Projektmanagement in der Spieleentwicklung Inhalt 1. Warum brauche ich ein Projekt-Management? 2. Die Charaktere des Projektmanagement - Mastermind - Producer - Projektleiter 3. Schnittstellen definieren
Q1 B1 KW 49. Genregulation
 Q1 B1 KW 49 Genregulation Transkription Posttranskription elle Modifikation Genregulation bei Eukaryoten Transkriptionsfaktoren (an TATA- Box) oder Silencer (verringert Transkription) und Enhancer (erhöht
Q1 B1 KW 49 Genregulation Transkription Posttranskription elle Modifikation Genregulation bei Eukaryoten Transkriptionsfaktoren (an TATA- Box) oder Silencer (verringert Transkription) und Enhancer (erhöht
Screening Das Programm. zur Früherkennung von Brustkrebs
 Mammographie Screening Das Programm zur Früherkennung von Brustkrebs das Mammographie Screening Programm Wenn Sie zwischen 50 und 69 Jahre alt sind, haben Sie alle zwei Jahre Anspruch auf eine Mammographie-Untersuchung
Mammographie Screening Das Programm zur Früherkennung von Brustkrebs das Mammographie Screening Programm Wenn Sie zwischen 50 und 69 Jahre alt sind, haben Sie alle zwei Jahre Anspruch auf eine Mammographie-Untersuchung
aktive / passive Immunisierung
 FS 2010 aktive / passive Immunisierung DTP-Impfstoff Michelle, Andrea 3Ma Überblick Wo befinden wir uns eigentlich? 10 000 Mia. Zellen Nerven-, Muskel-, Bindegewebsoder Blutzellen weißes BK (Leukozyt)
FS 2010 aktive / passive Immunisierung DTP-Impfstoff Michelle, Andrea 3Ma Überblick Wo befinden wir uns eigentlich? 10 000 Mia. Zellen Nerven-, Muskel-, Bindegewebsoder Blutzellen weißes BK (Leukozyt)
Repräsentative Umfrage zur Beratungsqualität im deutschen Einzelhandel (Auszug)
") Porsche Consulting Exzellent handeln Repräsentative Umfrage zur Beratungsqualität im deutschen Einzelhandel (Auszug) Oktober 2013 Inhalt Randdaten der Studie Untersuchungsziel der Studie Ergebnisse der
Porsche Consulting Exzellent handeln Repräsentative Umfrage zur Beratungsqualität im deutschen Einzelhandel (Auszug) Oktober 2013 Inhalt Randdaten der Studie Untersuchungsziel der Studie Ergebnisse der
geben. Die Wahrscheinlichkeit von 100% ist hier demnach nur der Gehen wir einmal davon aus, dass die von uns angenommenen
 geben. Die Wahrscheinlichkeit von 100% ist hier demnach nur der Vollständigkeit halber aufgeführt. Gehen wir einmal davon aus, dass die von uns angenommenen 70% im Beispiel exakt berechnet sind. Was würde
geben. Die Wahrscheinlichkeit von 100% ist hier demnach nur der Vollständigkeit halber aufgeführt. Gehen wir einmal davon aus, dass die von uns angenommenen 70% im Beispiel exakt berechnet sind. Was würde
APP-GFP/Fluoreszenzmikroskop. Aufnahmen neuronaler Zellen, mit freund. Genehmigung von Prof. Stefan Kins, TU Kaiserslautern
 Über die Herkunft von Aβ42 und Amyloid-Plaques Heute ist sicher belegt, dass sich die amyloiden Plaques aus einer Vielzahl an Abbaufragmenten des Amyloid-Vorläufer-Proteins (amyloid-precursor-protein,
Über die Herkunft von Aβ42 und Amyloid-Plaques Heute ist sicher belegt, dass sich die amyloiden Plaques aus einer Vielzahl an Abbaufragmenten des Amyloid-Vorläufer-Proteins (amyloid-precursor-protein,
Psychologie im Arbeitsschutz
 Fachvortrag zur Arbeitsschutztagung 2014 zum Thema: Psychologie im Arbeitsschutz von Dipl. Ing. Mirco Pretzel 23. Januar 2014 Quelle: Dt. Kaltwalzmuseum Hagen-Hohenlimburg 1. Einleitung Was hat mit moderner
Fachvortrag zur Arbeitsschutztagung 2014 zum Thema: Psychologie im Arbeitsschutz von Dipl. Ing. Mirco Pretzel 23. Januar 2014 Quelle: Dt. Kaltwalzmuseum Hagen-Hohenlimburg 1. Einleitung Was hat mit moderner
Mehr Geld verdienen! Lesen Sie... Peter von Karst. Ihre Leseprobe. der schlüssel zum leben. So gehen Sie konkret vor!
 Peter von Karst Mehr Geld verdienen! So gehen Sie konkret vor! Ihre Leseprobe Lesen Sie...... wie Sie mit wenigen, aber effektiven Schritten Ihre gesteckten Ziele erreichen.... wie Sie die richtigen Entscheidungen
Peter von Karst Mehr Geld verdienen! So gehen Sie konkret vor! Ihre Leseprobe Lesen Sie...... wie Sie mit wenigen, aber effektiven Schritten Ihre gesteckten Ziele erreichen.... wie Sie die richtigen Entscheidungen
Chemotherapie -ein Bilderbuch für Kinder
 Chemotherapie -ein Bilderbuch für Kinder Unser Körper besteht aus verschiedenen Zellen, die ganz unterschiedlich aussehen. Jede Art erfüllt eine besondere Aufgabe. Da gibt es zum Beispiel Gehirnzellen,
Chemotherapie -ein Bilderbuch für Kinder Unser Körper besteht aus verschiedenen Zellen, die ganz unterschiedlich aussehen. Jede Art erfüllt eine besondere Aufgabe. Da gibt es zum Beispiel Gehirnzellen,
Abbildungsverzeichnis
 I INHALTSVERZEICHNIS Abkürzungen VI Abbildungsverzeichnis VIII I. Einleitung 1. Neurone und Axonwachstum 1 2. Oligodendrozyten und Myelin 3 3. Das Proteolipid Protein (PLP) 6 4. Mutationen im PLP-Gen und
I INHALTSVERZEICHNIS Abkürzungen VI Abbildungsverzeichnis VIII I. Einleitung 1. Neurone und Axonwachstum 1 2. Oligodendrozyten und Myelin 3 3. Das Proteolipid Protein (PLP) 6 4. Mutationen im PLP-Gen und
Psychosen. By Kevin und Oliver
 Psychosen By Kevin und Oliver 9. Umgang Inhaltsverzeichnis 1. Definition Psychosen 2. Häufigkeit 3. Ursachen 4. Formen / Symptome 5. Phasen einer Psychose 6. Diagnosen 7. Medikamente 8. Therapie Definition
Psychosen By Kevin und Oliver 9. Umgang Inhaltsverzeichnis 1. Definition Psychosen 2. Häufigkeit 3. Ursachen 4. Formen / Symptome 5. Phasen einer Psychose 6. Diagnosen 7. Medikamente 8. Therapie Definition
Diese Broschüre fasst die wichtigsten Informationen zusammen, damit Sie einen Entscheid treffen können.
 Aufklärung über die Weiterverwendung/Nutzung von biologischem Material und/oder gesundheitsbezogen Daten für die biomedizinische Forschung. (Version V-2.0 vom 16.07.2014, Biobanken) Sehr geehrte Patientin,
Aufklärung über die Weiterverwendung/Nutzung von biologischem Material und/oder gesundheitsbezogen Daten für die biomedizinische Forschung. (Version V-2.0 vom 16.07.2014, Biobanken) Sehr geehrte Patientin,
Gleichungen Lösen. Ein graphischer Blick auf Gleichungen
 Gleichungen Lösen Was bedeutet es, eine Gleichung zu lösen? Was ist überhaupt eine Gleichung? Eine Gleichung ist, grundsätzlich eine Aussage über zwei mathematische Terme, dass sie gleich sind. Ein Term
Gleichungen Lösen Was bedeutet es, eine Gleichung zu lösen? Was ist überhaupt eine Gleichung? Eine Gleichung ist, grundsätzlich eine Aussage über zwei mathematische Terme, dass sie gleich sind. Ein Term
Anleitung über den Umgang mit Schildern
 Anleitung über den Umgang mit Schildern -Vorwort -Wo bekommt man Schilder? -Wo und wie speichert man die Schilder? -Wie füge ich die Schilder in meinen Track ein? -Welche Bauteile kann man noch für Schilder
Anleitung über den Umgang mit Schildern -Vorwort -Wo bekommt man Schilder? -Wo und wie speichert man die Schilder? -Wie füge ich die Schilder in meinen Track ein? -Welche Bauteile kann man noch für Schilder
Meinungen der Bürgerinnen und Bürger in Hamburg und Berlin zu einer Bewerbung um die Austragung der Olympischen Spiele
 Meinungen der Bürgerinnen und Bürger in Hamburg und Berlin zu einer Bewerbung um die Austragung der Olympischen Spiele 4. März 2015 q5337/31319 Le forsa Politik- und Sozialforschung GmbH Büro Berlin Schreiberhauer
Meinungen der Bürgerinnen und Bürger in Hamburg und Berlin zu einer Bewerbung um die Austragung der Olympischen Spiele 4. März 2015 q5337/31319 Le forsa Politik- und Sozialforschung GmbH Büro Berlin Schreiberhauer
1) Warum sollten Sie die Bindehautentzündung grundsätzlich vom Arzt behandeln lassen? 4. 2) Was ist eine Bindehautentzündung? 5
 Warum sollten Sie die Bindehautentzündung grundsätzlich vom Arzt behandeln lassen? 4. 2) Was ist eine Bindehautentzündung? 5") 1) Warum sollten Sie die Bindehautentzündung grundsätzlich vom Arzt behandeln lassen? 4 2) Was ist eine Bindehautentzündung? 5 3) Durch welche Symptome kann sich eine Bindehautentzündung bemerkbar machen?
1) Warum sollten Sie die Bindehautentzündung grundsätzlich vom Arzt behandeln lassen? 4 2) Was ist eine Bindehautentzündung? 5 3) Durch welche Symptome kann sich eine Bindehautentzündung bemerkbar machen?
Fernausbildung Fachberater/in für holistische Gesundheit. Modul 6
 Fernausbildung Fachberater/in für holistische Gesundheit Modul 6 Vorwort Liebe(r) Teilnehmer/in Herzlich willkommen in Modul 6 Mit dem 6. Modul beenden Sie nun Ihre Ausbildung, denn einmal muss schliesslich
Fernausbildung Fachberater/in für holistische Gesundheit Modul 6 Vorwort Liebe(r) Teilnehmer/in Herzlich willkommen in Modul 6 Mit dem 6. Modul beenden Sie nun Ihre Ausbildung, denn einmal muss schliesslich
Patientenverfügung. Was versteht man genau unter einer Patientenverfügung? Meine persönliche Patientenverfügung
 Patientenverfügung Erarbeitet vom Klinischen Ethikkomitee im Klinikum Herford. Im Komitee sind sowohl Ärzte als auch Mitarbeitende der Pflegedienste, Theologen und ein Jurist vertreten. Der Text entspricht
Patientenverfügung Erarbeitet vom Klinischen Ethikkomitee im Klinikum Herford. Im Komitee sind sowohl Ärzte als auch Mitarbeitende der Pflegedienste, Theologen und ein Jurist vertreten. Der Text entspricht
Tag des Datenschutzes
 Tag des Datenschutzes Datenschutz und Software: Vertrauen ist gut, Kontrolle ist besser Dr. Michael Stehmann Zur Person Rechtsanwalt Dr. Michael Stehmann Studium der Rechtswissenschaft an der Universität
Tag des Datenschutzes Datenschutz und Software: Vertrauen ist gut, Kontrolle ist besser Dr. Michael Stehmann Zur Person Rechtsanwalt Dr. Michael Stehmann Studium der Rechtswissenschaft an der Universität
Der Gesundheitskompass für die junge Frau
 Haarausfall? Blutentnahme zu Hormonlabor, Eisenwerte, Schilddrüsenwerte, Biotin und Zink 86,87 EURO Wiederholte Scheideninfektionen? Mit dem Vaginalstatus können Infektionen durch spezielle Kulturen auf
Haarausfall? Blutentnahme zu Hormonlabor, Eisenwerte, Schilddrüsenwerte, Biotin und Zink 86,87 EURO Wiederholte Scheideninfektionen? Mit dem Vaginalstatus können Infektionen durch spezielle Kulturen auf
Gutes Leben was ist das?
 Lukas Bayer Jahrgangsstufe 12 Im Hirschgarten 1 67435 Neustadt Kurfürst-Ruprecht-Gymnasium Landwehrstraße22 67433 Neustadt a. d. Weinstraße Gutes Leben was ist das? Gutes Leben für alle was genau ist das
Lukas Bayer Jahrgangsstufe 12 Im Hirschgarten 1 67435 Neustadt Kurfürst-Ruprecht-Gymnasium Landwehrstraße22 67433 Neustadt a. d. Weinstraße Gutes Leben was ist das? Gutes Leben für alle was genau ist das
Geld Verdienen im Internet leicht gemacht
 Geld Verdienen im Internet leicht gemacht Hallo, Sie haben sich dieses E-book wahrscheinlich herunter geladen, weil Sie gerne lernen würden wie sie im Internet Geld verdienen können, oder? Denn genau das
Geld Verdienen im Internet leicht gemacht Hallo, Sie haben sich dieses E-book wahrscheinlich herunter geladen, weil Sie gerne lernen würden wie sie im Internet Geld verdienen können, oder? Denn genau das
PCD Europe, Krefeld, Jan 2007. Auswertung von Haemoccult
 Auswertung von Haemoccult Ist das positiv? Nein! Ja! Im deutschen Krebsfrüherkennungsprogramm haben nur etwa 1 % der Frauen und 1,5 % der Männer ein positives Haemoccult -Ergebnis, da dieser Test eine
Auswertung von Haemoccult Ist das positiv? Nein! Ja! Im deutschen Krebsfrüherkennungsprogramm haben nur etwa 1 % der Frauen und 1,5 % der Männer ein positives Haemoccult -Ergebnis, da dieser Test eine
Tipp III: Leiten Sie eine immer direkt anwendbare Formel her zur Berechnung der sogenannten "bedingten Wahrscheinlichkeit".
 Mathematik- Unterrichts- Einheiten- Datei e. V. Klasse 9 12 04/2015 Diabetes-Test Infos: www.mued.de Blutspenden werden auf Diabetes untersucht, das mit 8 % in der Bevölkerung verbreitet ist. Dabei werden
Mathematik- Unterrichts- Einheiten- Datei e. V. Klasse 9 12 04/2015 Diabetes-Test Infos: www.mued.de Blutspenden werden auf Diabetes untersucht, das mit 8 % in der Bevölkerung verbreitet ist. Dabei werden
Ist Fernsehen schädlich für die eigene Meinung oder fördert es unabhängig zu denken?
 UErörterung zu dem Thema Ist Fernsehen schädlich für die eigene Meinung oder fördert es unabhängig zu denken? 2000 by christoph hoffmann Seite I Gliederung 1. In zu großen Mengen ist alles schädlich. 2.
UErörterung zu dem Thema Ist Fernsehen schädlich für die eigene Meinung oder fördert es unabhängig zu denken? 2000 by christoph hoffmann Seite I Gliederung 1. In zu großen Mengen ist alles schädlich. 2.
DISSERTATION. Genanalyse bei Patienten mit Akuter Intermittierender Porphyrie. Zur Erlangung des akademischen Grades Doctor Medicinae (Dr. med.
 Aus der Medizinischen Klinik mit Schwerpunkt Onkologie und Hämatologie Charité Campus Mitte Charité Universitätsmedizin Berlin und dem Hämatologisch-Onkologischen Zentrum München DISSERTATION Genanalyse
Aus der Medizinischen Klinik mit Schwerpunkt Onkologie und Hämatologie Charité Campus Mitte Charité Universitätsmedizin Berlin und dem Hämatologisch-Onkologischen Zentrum München DISSERTATION Genanalyse
Unterrichtsmaterialien in digitaler und in gedruckter Form. Auszug aus: Antibiotikaresistenz bei Pseudomonaden
 Unterrichtsmaterialien in digitaler und in gedruckter Form Auszug aus: Antibiotikaresistenz bei Pseudomonaden Das komplette Material finden Sie hier: School-Scout.de S 1 M 6 M 7 Erarbeitung II, Präsentation
Unterrichtsmaterialien in digitaler und in gedruckter Form Auszug aus: Antibiotikaresistenz bei Pseudomonaden Das komplette Material finden Sie hier: School-Scout.de S 1 M 6 M 7 Erarbeitung II, Präsentation
Vor- und Nachteile der Kastration
 Vor- und Nachteile der Kastration Was versteht man unter Kastration? Unter Kastration versteht man die chirugische Entfernung der Keimdrüsen. Bei der Hündin handelt es sich dabei um die Eierstöcke, beim
Vor- und Nachteile der Kastration Was versteht man unter Kastration? Unter Kastration versteht man die chirugische Entfernung der Keimdrüsen. Bei der Hündin handelt es sich dabei um die Eierstöcke, beim
1 MIO ÖSTERREICHISCHE SKIFAHRER SCHÜTZEN SICH BEREITS MIT HELM - UM 370.000 MEHR ALS IM VORJAHR
 1 MIO ÖSTERREICHISCHE SKIFAHRER SCHÜTZEN SICH BEREITS MIT HELM - UM 370.000 MEHR ALS IM VORJAHR 3/09 1 MIO ÖSTERREICHISCHE SKIFAHRER SCHÜTZEN SICH BEREITS MIT HELM - UM 370.000 MEHR ALS IM VORJAHR Mehr
1 MIO ÖSTERREICHISCHE SKIFAHRER SCHÜTZEN SICH BEREITS MIT HELM - UM 370.000 MEHR ALS IM VORJAHR 3/09 1 MIO ÖSTERREICHISCHE SKIFAHRER SCHÜTZEN SICH BEREITS MIT HELM - UM 370.000 MEHR ALS IM VORJAHR Mehr
Wissenschaftlicher Bericht
 Ein Auszug aus... Wissenschaftlicher Bericht Augmented Reality als Medium strategischer medialer Kommunikation Die komplette Studie ist bei amazon.de käuflich zu erwerben. Inhaltsverzeichnis 1 Einführung
Ein Auszug aus... Wissenschaftlicher Bericht Augmented Reality als Medium strategischer medialer Kommunikation Die komplette Studie ist bei amazon.de käuflich zu erwerben. Inhaltsverzeichnis 1 Einführung
LU-Zerlegung. Zusätze zum Gelben Rechenbuch. Peter Furlan. Verlag Martina Furlan. Inhaltsverzeichnis. 1 Definitionen.
 Zusätze zum Gelben Rechenbuch LU-Zerlegung Peter Furlan Verlag Martina Furlan Inhaltsverzeichnis Definitionen 2 (Allgemeine) LU-Zerlegung 2 3 Vereinfachte LU-Zerlegung 3 4 Lösung eines linearen Gleichungssystems
Zusätze zum Gelben Rechenbuch LU-Zerlegung Peter Furlan Verlag Martina Furlan Inhaltsverzeichnis Definitionen 2 (Allgemeine) LU-Zerlegung 2 3 Vereinfachte LU-Zerlegung 3 4 Lösung eines linearen Gleichungssystems
Lichtbrechung an Linsen
 Sammellinsen Lichtbrechung an Linsen Fällt ein paralleles Lichtbündel auf eine Sammellinse, so werden die Lichtstrahlen so gebrochen, dass sie durch einen Brennpunkt der Linse verlaufen. Der Abstand zwischen
Sammellinsen Lichtbrechung an Linsen Fällt ein paralleles Lichtbündel auf eine Sammellinse, so werden die Lichtstrahlen so gebrochen, dass sie durch einen Brennpunkt der Linse verlaufen. Der Abstand zwischen
BIS Infobrief November 2014
 Bayerisches Landesamt für Gesundheit und Lebensmittelsicherheit BIS Infobrief November 2014 Sehr geehrte Kolleginnen und Kollegen, wir bedanken uns ganz herzlich bei Ihnen für Ihre aktive Teilnahme am
Bayerisches Landesamt für Gesundheit und Lebensmittelsicherheit BIS Infobrief November 2014 Sehr geehrte Kolleginnen und Kollegen, wir bedanken uns ganz herzlich bei Ihnen für Ihre aktive Teilnahme am
InVo. Information zu Verordnungen in der GKV. Herstellung von Arzneimitteln durch Ärzte Anzeigepflicht bei Bezirksregierungen. Stand: Februar 2010
 Nr. 1 2010 InVo Information zu Verordnungen in der GKV Stand: Februar 2010 Herstellung von Arzneimitteln durch Ärzte Anzeigepflicht bei Bezirksregierungen Bisher konnten Sie als Arzt Arzneimittel (z. B.
Nr. 1 2010 InVo Information zu Verordnungen in der GKV Stand: Februar 2010 Herstellung von Arzneimitteln durch Ärzte Anzeigepflicht bei Bezirksregierungen Bisher konnten Sie als Arzt Arzneimittel (z. B.
Mobile Intranet in Unternehmen
 Mobile Intranet in Unternehmen Ergebnisse einer Umfrage unter Intranet Verantwortlichen aexea GmbH - communication. content. consulting Augustenstraße 15 70178 Stuttgart Tel: 0711 87035490 Mobile Intranet
Mobile Intranet in Unternehmen Ergebnisse einer Umfrage unter Intranet Verantwortlichen aexea GmbH - communication. content. consulting Augustenstraße 15 70178 Stuttgart Tel: 0711 87035490 Mobile Intranet
Inhalte unseres Vortrages
 Inhalte unseres Vortrages Vorstellung der beiden paper: Germ line transmission of a disrupted ß2 mirkroglobulin gene produced by homologous recombination in embryonic stem cells ß2 Mikroglobulin deficient
Inhalte unseres Vortrages Vorstellung der beiden paper: Germ line transmission of a disrupted ß2 mirkroglobulin gene produced by homologous recombination in embryonic stem cells ß2 Mikroglobulin deficient
Zeit lässt sich nicht wie Geld für schlechte Zeiten zur Seite legen. Die Zeit vergeht egal, ob genutzt oder ungenutzt.
 Zeitmanagement Allgemeine Einleitung Wie oft haben Sie schon gehört Ich habe leider keine Zeit? Und wie oft haben Sie diesen Satz schon selbst gesagt? Wahrscheinlich nahezu jeden Tag. Dabei stimmt der
Zeitmanagement Allgemeine Einleitung Wie oft haben Sie schon gehört Ich habe leider keine Zeit? Und wie oft haben Sie diesen Satz schon selbst gesagt? Wahrscheinlich nahezu jeden Tag. Dabei stimmt der
6 Schulungsmodul: Probenahme im Betrieb
 6 Schulungsmodul: Probenahme im Betrieb WIEDNER Wie schon im Kapitel VI erwähnt, ist die Probenahme in Betrieben, die Produkte nach dem Lebensmittel- und Futtermittelgesetzbuch herstellen oder in den Verkehr
6 Schulungsmodul: Probenahme im Betrieb WIEDNER Wie schon im Kapitel VI erwähnt, ist die Probenahme in Betrieben, die Produkte nach dem Lebensmittel- und Futtermittelgesetzbuch herstellen oder in den Verkehr
Persönliche Zukunftsplanung mit Menschen, denen nicht zugetraut wird, dass sie für sich selbst sprechen können Von Susanne Göbel und Josef Ströbl
 Persönliche Zukunftsplanung mit Menschen, denen nicht zugetraut Von Susanne Göbel und Josef Ströbl Die Ideen der Persönlichen Zukunftsplanung stammen aus Nordamerika. Dort werden Zukunftsplanungen schon
Persönliche Zukunftsplanung mit Menschen, denen nicht zugetraut Von Susanne Göbel und Josef Ströbl Die Ideen der Persönlichen Zukunftsplanung stammen aus Nordamerika. Dort werden Zukunftsplanungen schon
Handbuch. Artologik EZ-Equip. Plug-in für EZbooking version 3.2. Artisan Global Software
 Artologik EZ-Equip Plug-in für EZbooking version 3.2 Artologik EZbooking und EZ-Equip EZbooking, Ihre webbasierte Software zum Reservieren von Räumen und Objekten, kann nun durch die Ergänzung um ein oder
Artologik EZ-Equip Plug-in für EZbooking version 3.2 Artologik EZbooking und EZ-Equip EZbooking, Ihre webbasierte Software zum Reservieren von Räumen und Objekten, kann nun durch die Ergänzung um ein oder
Leseprobe. Thomas Konert, Achim Schmidt. Design for Six Sigma umsetzen ISBN: 978-3-446-41230-9. Weitere Informationen oder Bestellungen unter
 Leseprobe Thomas Konert, Achim Schmidt Design for Six Sigma umsetzen ISBN: 978-3-446-41230-9 Weitere Informationen oder Bestellungen unter http://www.hanser.de/978-3-446-41230-9 sowie im Buchhandel. Carl
Leseprobe Thomas Konert, Achim Schmidt Design for Six Sigma umsetzen ISBN: 978-3-446-41230-9 Weitere Informationen oder Bestellungen unter http://www.hanser.de/978-3-446-41230-9 sowie im Buchhandel. Carl