4. Ergebnisse. Spezies Signalpeptid B- C-Domäne A-Domäne Total a
|
|
|
- Artur Jaeger
- vor 6 Jahren
- Abrufe
Transkript
1 4. Ergebnisse 4.1. Relaxin Equiden Molekulare Charakterisierung Prorelaxin cdna-sequenz Mit zwei degenerierten Oligonukleotidprimern, die entsprechend der bekannten Peptid-sequenzen der A- und B-Domäne für equines Prorelaxin hergestellt wurden (siehe ; Stewart et al., 1991), erfolgte der RT-PCR-Nachweis eines 428 bp Amplifikates aus Gesamt-RNA von Pferdeplazenten am Tag 120 und 300 der Trächtigkeit sowie von zwei Präimplan-tationsembryonen am Tag 30 der Gravidität, bei denen die Choriongürtelzellen zuvor abpräpariert worden waren. In den Zellen des Allantochorions, nicht jedoch in den Choriongürtelzellen, wurde Relaxin-mRNA detektiert (Abb. 2). Die partielle cdna-sequenz für equines Prorelaxin kodierte für die C- terminalen 22 Aminosäuren (66 bp) der B-Domäne, eine C-Domäne von 109 Aminosäuren (327 bp) und die N-terminalen 12 Aminosäuren (36 bp) der A-Domäne (Abb. 3). Tabelle 12. Aminosäure-Homologien des equinen Präprorelaxins und der einzelnen Relaxin- Domänen mit Relaxinen anderer Spezies. Spezies Signalpeptid B- C-Domäne A-Domäne Total a Domäne Schwein 66,7 64,3 67,6 54,5 65,6 Dromedar 76,0 69,2 61,9 50,0 65,4 H1 b 52,0 57,1 53,8 45,5 52,2 H2 b 50,0 53,6 50,5 50,0 50,0 Katze 60,0 65,4 61,2 59,1 59,9 Hund 72,0 44,5 23,7 59,1 48,0 Ratte 57,1 42,9 40,4 45,5 41,2 Maus 54,5 42,9 48,5 27,3 40,1 a Prozentsatz identischer Aminosäuren b H1 und H2: die zwei humanen Präprorelaxine Die potentielle Rezeptorbindungssequenz in der B-Domäne wich in einer Aminosäure (V A) vom klassischen Rezeptorbindungsmotif (-G-R-E-L-V-R-) ab (Abb. 3) und war identisch mit der kürzlich publizierten equinen Präprorelaxinsequenz (Min et al., 1996). Equines Prorelaxin zeigte die 36
 mit total RNA einer Pferdeplazenta am Tag 300 (2) und 120 (3) der Gravidität. Total RNA aus dem Cerebellum einer Stute diente als Negativkontrolle (1). 4.1.1.2. Expression und Lokalisation im Gewebe 4.")

2 höchste Homologie zu den Prorelaxinen des Schweines und des Dromedars (Tab. 12; Klonisch et al., 1995). Abb. 1. Northern Analyse Die Northern Analyse ergab ein spezifisches Hybridisierungssignal für Relaxin bei 1 kb (Abb. 1) mit total RNA einer Pferdeplazenta am Tag 300 (2) und 120 (3) der Gravidität. Total RNA aus dem Cerebellum einer Stute diente als Negativkontrolle (1) Expression und Lokalisation im Gewebe Präimplantationsembryonen Es gelang der RT-PCR Nachweis spezifischer Amplifikate für Prorelaxin im Allantochorion, zwei Präimplantationsembryonen des Pferdes am Tag 30 der Gravidität. In Choriongürtelzellen zweier Präimplantationsembryonen am Tag 33 der Trächtigkeit sowie in den aus diesen invasiven Trophoblastzellen hervorgehenden endometrialen Cupzellen konnten keine Relaxin-Transkripte oder immunreaktives Relaxin nachgewiesen werden. Abb. 2. Ergebnisse der RT-PCR für Prorelaxin an Präimplantationsembryonen des Pferdes am Tag 30 der Gravidität. Keine Expression von Relaxin-mRNA in Trophoblasten des Choriongürtels Expression von Relaxin-mRNA im Allantochorion 37
3 Abb. 3. Die partielle cdna-sequenz für equines Prorelaxin. Die verwendeten Primer sind durch Pfeile gekennzeichnet und die potentielle Rezeptorbindungsstelle in der B-Domäne ist hervorgehoben. B-Domäne 32 Ile Lys Ala Cys Gly Arg Glu Leu Ala Arg Leu Arg Ile Glu Ile Cys Gly Ser Leu Ser Trp Lys Lys ATT AAA GCA TGC GGC CGC GAA TTA GCA CGC CTG CGG ATC GAG ATC TGC GGC TCC CTC TCC TGG AAG AAA 162 C-Domäne 55 Thr Val Leu Arg Leu Glu Glu Pro Gly Leu Glu Val Gly Gln Pro Val Glu Ile Val Ser Ser Ser Ile ACG GTT CTG AGG CTG GAG GAG CCT GGG CTG GAA GTC GGA CAA CCC GTA GAA ATT GTG TCA TCC TCC ATC Ser Lys Asp Ale Glu Ala Leu Asn Thr Lys Leu Gly Leu Asn Ser Asn Leu Pro Lys Glu Gln Lys Ala AGC AAA GAT GCA GAA GCT TTA AAT ACG AAG TTG GGA CTC AAT TCT AAT TTG CCA AAA GAG CAG AAA GCA Thr Leu Ser Glu Arg Gln Pro Ser Trp Arg Glu Leu Leu Gln Gln Pro Ala Leu Lys Asp Ser Asn Leu ACA CTG TCT GAG AGG CAG CCA TCA TGG AGG GAG CTG CTA CAA CAA CCT GCA TTA AAG GAT TCA AAT CTT Asn Leu Glu Glu Phe Glu Glu Thr Ile Gln Lys Thr Gln Ser Glu Val Glu Asp Asp Ser Leu Ser Glu AAC TTG GAA GAA TTT GAG GAA ACC ATT CAG AAG ACA CAA AGT GAA GTA GAA GAT GAC AGT CTT TCA GAA 438 A-Domäne 147 Leu Lys Asn Leu Gly Leu Asp Lys His Ser Arg Lys Lys Arg Met Ile Gln Leu Ser His Lys Cys Cys TTA AAG AAC CTA GGC TTA GAT AAA CAT TCT CGT AAA AAG AGA ATG ATT CAA CTG AGT CAT AAA TGC TGC Tyr Trp Gly Cys Thr TAC TGG GGT TGT AC
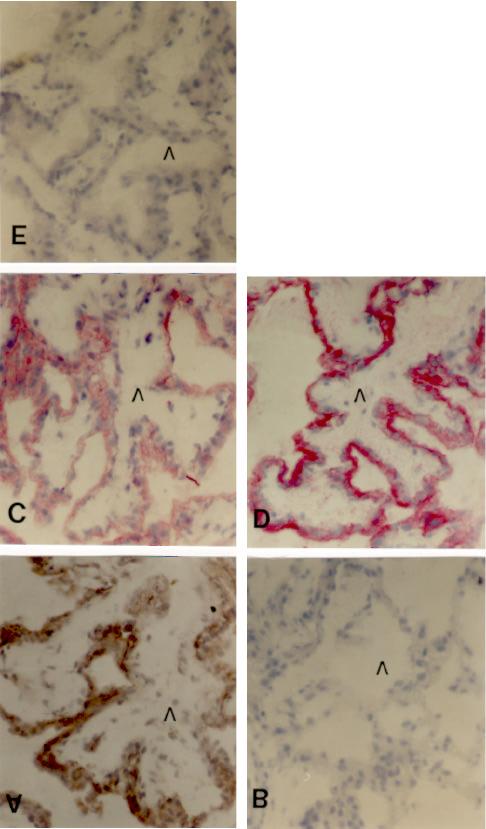
4 Uteroplazentares Gewebe Relaxin mrna (Abb. 4a, b, d) und immunreaktives Relaxin (Abb. 4e) waren in den Plazenten der intra-, inter- und extraspezifischen Graviditäten in fetalen Zellen exprimiert, die mit dem monoklonalen Antikörper F102.1 als equine Trophoblasten identifiziert wurden (Abb. 4c). Weder der Genotyp noch die immunologischen Abwehrmechanismen gegen den Eselembryo während der frühen Phase der extraspezifischen Gravidität schienen die Relaxin-Expression zu beeinflussen. In den Plazenten dieser Graviditätstypen (Tab. 3) konnten bezüglich ihrer Morphologie und ihres Relaxin- Expressionsmusters zwei Trophoblast-Populationen unter-schieden werden. Kuboidale Trophoblasten der frühen (Abb. 4a) und reifen Mikrokotyledonen (Abb. 4d) exprimierten Relaxin mrna und Protein (Abb. 4e). Elongierte, Relaxin-negative Trophoblasten waren während der Plazentation zwischen Bereichen Relaxin-positiver kuboidaler Trophoblasten nachweisbar (Abb. 4a, b, d und Abb. 5b) und in der reifen equinen Plazenta ausschließlich an der Basis der Mikrokotyledonen gegenüber der Öffnung uteriner Drüsen lokalisiert (Abb. 4d und Abb. 5b; Klonisch et al., 1997). Elongierte Trophoblasten der Mikrokotyledonen präsentierten spezifisch immunreaktive Major Histocompatibility Complex (MHC) Klasse I Moleküle (Abb. 5a). Abb. 4. Nachweis von Relaxin mrna in fetalen Trophoblasten (Tb) von Gefrierschnitten uteroplazentaren Gewebes am Tag 45 (a) und Tag 60 (b) einer interspezifischen Gravidität. Elongierte Trophoblasten (pseudostratified; Ps) exprimierten keine Relaxin mrna (Pfeile). (c) Kuboidale und elongierte (Ps) Trophoblasten wurden mit dem für equine Trophoblasten spezifischen monoklonalen Antikörper F102.1 identifiziert. (d) Lokalisation von Relaxin-mRNA in kuboidalen Trophoblasten der Mikrokotyledonen (Mc) am Tag 153 einer intraspezifischen Gravidität. Elongierte Trophoblasten (Ps) an der Basis der Mikrokotyledonen (Mc) exprimierten keine Relaxin mrna. (e) Nachweis immunoreaktiven Relaxins mit einem Antiserum gegen equines Relaxin (Nr. 3150) am Tag 153 der intraspezifischen Gravidität. Bereiche elongierter Trophoblasten (Ps) gegenüber der Öffnung endometrialer Drüsen (endometrial glands; Eg) zeigten keine Färbung. (f) In der Negativkontrolle zeigten fetales Allantochorion (Ac) und maternales Endometrium (E) einer Plazenta am Tag 60 der intraspezifischen Gravidität keine immunhistochemische Färbung. Maßstab: 100 µm. 39

5 40
 Klasse I-Molekülen (a) und Relaxin mrna-exprimie-renden Trophoblasten (b) an")
6 Abb. 5. Vergleichende Darstellung der zellulären Lokalisation von immunoreaktiven Major Histocompatibility Complex (MHC) Klasse I-Molekülen (a) und Relaxin mrna-exprimie-renden Trophoblasten (b) an Kryoschnitten einer Pferdeplazenta am Tag 153 der intraspezifischen Gravidität. Der Nachweis der MHC Klasse I-Moleküle erfolgte mit dem monoklonalen Rattenantikörper JK 56 in elongierten fetalen Trophoblasten (Ps; pseudo-stratified) der Areolae des Allantochorion sowie in uterinen Stromazellen in den Mikrokotyledonen (Mc; a). Kuboidale Trophoblasten der Mikrokotyledonen sowie fetale Stromazellen exprimierten keine MHC Klasse I- Moleküle (a). Relaxin-Transkripte wurden ausschließlich in kuboidalen Trophoblasten der Mikrolotyledonen (Mc), nicht jedoch in den MHC Klasse I-positiven, elongierten Trophoblasten (Ps) nachgewiesen. Die elongierten Trophoblasten (Ps) waren häufig gegenüber der Öffnung endometrialen Drüsen (ed) lokalisiert (a, b). Vergrößerung: a, b x
.")
. Abb. 6.")

7 Ovar In ovariellen Follikeln verschiedener Größen sowie in Corpora albicantia der zyklischen Stute konnte mit RT-PCR keine Relaxin-mRNA nachgewiesen werden. Der PCR-Nachweis für Relaxin mrna gelang in 3-17 Tage alten Corpora lutea des zyklischen Ovars und wurde durch Southern-Analyse bestätigt (Abb. 6). Gleiche Mengen an Gesamt-RNA waren für die RT-PCR eingesetzt worden. Als Positivkontrolle diente eine cdna von plazentarem Gewebe am Tag 120 der Trächtigkeit (Abb. 6). Negativkontrollen unter Ausschluß der reversen Transkriptase oder dem Einsatz von Gesamt-RNA als Template ergaben keine Amplifikate oder Hybridisierungssignale (nicht gezeigt). Abb. 6. Southern Analyse mit einer Digoxigenin-markierten antisense crna Sonde zur Verifikation der RT-PCR-Amplifikate für equines Prorelaxin in den Corpora lutea. 1-5: 1, 2, 3, 4 und > 5 cm große Follikel 6: frühes Corpus luteum 7-8: 3-7 Tage altes Corpus luteum 9: 9-11 Tage altes Corpus luteum; 10-11: Tage altes Corpus luteum; 12: Corpus albicans; 13: Pferdeplazenta am Tag 120 der Gravidität (Positivkontrolle; Ryan et al., 1997). Abb. 7. Mit einer Digoxigeninmarkierten antisense crna- Sonde für equines Prorelaxin erfolgte der spezifische Nachweis von Hybridisierungssignalen in großen Lutealzellen (Pfeile) eines reifen equinen Corpus luteum (ca. Tag 9) (a). Inkubation mit einer sense crna Relaxin-Sonde ergab keine Hybridisierungssignale (b). Maßstab: 133 µm. 42
 zeigten, vermutlich aufgrund zu geringer Expressionsmengen, keine Hybridisierungssignale für RelaxinmRNA.")
8 In großen Lutealzellen der untersuchten Corpora lutea gelang der spezifische Nachweis von Relaxin- Transkripten (Abb. 7a+b). Die Follikel verschiedener Reifegrade (1-5 cm im Durchmesser) zeigten, vermutlich aufgrund zu geringer Expressionsmengen, keine Hybridisierungssignale für RelaxinmRNA. Der Nachweis von immunreaktivem Relaxin mit dem Kaninchen-Antiserum gegen equines Relaxin Nr (Stewart, 1986) gelang in den Zellen der Theca interna und den Granulosazellen reifer Follikel (ca. 5,5 cm; Abb. 8a) und war besonders stark in Lutealzellen des reifen Corpus luteum (Tag 9-11; Abb. 8c). Immunreaktives Relaxin war nicht im ovariellen Stroma und seriellen Schnitten von Follikeln und Corpora lutea nach Inkubation mit Nichtimmunserum des Kaninchens (Abb. 8b+d) nachweisbar. Abb. 8. Lokalisation von immunoreaktivem equinem Relaxin in Formalin-fixierten, paraffinierten Schnitten (10 µm) eines reifen Follikels (> 5,5 cm; a, b) sowie eines reifen Corpus luteum (ca. Tag 9; c,d). Die Schnitte wurden mit einem Antiserum gegen Pferderelaxin (Nr. 3150; a, c) oder mit Nichtimmunserum vom Kaninchen (b, d) als Negativkontrolle inkubiert. Im Follikel war immunoreaktives Relaxin in den Zellen der Theca interna (T) und der Granulosazellschicht (G) nachweisbar (a). Im Corpus luteum wurde immunoreaktives Relaxin in den Lutealzellen (Lc) detektiert (c). Das ovarielle Stroma (St), die Blutgefäße (Bv) sowie die Negativkontrollen (b, d) zeigten keine Färbung. Maßstab: a, b: 64µm; c, d: 80 µm. 43
9 Relaxinkonzentrationen in ovarieller Follikelflüssigkeit Der Relaxingehalt der Flüssigkeit in ovariellen Follikeln unterschiedlicher Größe wurde im homologen Radioimmunoassay (RIA) für equines Relaxin (Tab. 11; Stewart 1986) und durch High-Performance Liquid Chromatography (HPLC) bestimmt (Abb. 9a-d). Obwohl die durch RIA pro Milliliter Follikelflüssigkeit bestimmten Konzentrationen für Relaxin in Follikeln unterschiedlicher Größe gleich waren, stieg die Relaxin-Gesamtmenge mit zunehmendem Flüssigkeitsgehalt bis zum reifen präovulatorischen Follikel um das ca. 10-fache an (Tab. 13). Das HPLC-Absorptionsspektrum der 40%-Acetonitril-Fraktion der Follikelflüssigkeiten zeigte ein vergleichbares Spektrum wie gereinigtes equines Relaxin in 30% Acetonitril, welches als Standard bei allen HPLC-Analysen diente (Fig. 9ad). Die durch RIA ermittelten Fraktionen von immunoreaktivem equinem Relaxin in einem präovulatorischen Follikel korrelierten gut mit den zwischen min eluierten HPLC-Fraktionen. HPLC-Analyse der Flüssigkeit eines Corpus haemorrhagicum ergab im Vergleich zu einem großen Follikel ( 5 cm; 1,8 ng/ml) einen ca. 6-fache erhöhten (10,7 ng/ml) Relaxingehalt (Ryan et al., 1997). Tabelle 13. Gehalt an immunoreaktivem Relaxin in der Flüssigkeit equiner ovarieller Follikel unterschiedlicher Größe. Kategorie Anzahl der Konzentration Gesamtmenge Follikelgröße Follikelflüssig- (cm) Follikel (n) (ng/ml) (ng) (cm) keit (ml) sehr klein ( 2) 9 2,1 ± 0,3 7,9 ± 1,3 1,9 ± 0,06 3,8 ± 0,1 klein (> 2 3) 15 2,5 ± 0,3 18,6 ± 2,2 2,8 ± 0,06 7,4 ± 0,6 mittel(> 3 4) 13 3,1 ± 0,5 60,5 ± 9,7 3,8 ± 0,07 20,1 ± 1,4 groß (> 4) 16 2,4 ± 0,3 91,5 ± 9,2 5,3 ± 0,2 37,6 ± 1,5 44
10 Abb 9. Reverse-phase Chromatogramme für gereinigtes equines Relaxin als Standard (a) und follikuläre Flüssigkeit von ca. 2 cm, 3 cm und 5,5 cm großen ovariellen Follikeln zyklischer Stuten (b-d). Die Profile in (b + c) resultierten von gepoolter Follikelflüssigkeit aus zwei bis drei Follikeln. Die im RIA ermittelte Werte immunoreaktiven Relaxins (in pg) für die Elutionsspitzen der Follikelflüssigkeit eines 5,5 cm Follikels sind angegeben (d). 45
11 Zusammenfassung Im equinen Präimplantationsembryo exprimiert nur der nicht-invasive Trophoblast RelaxinmRNA, nicht jedoch die MHC Klasse I-Moleküle immunopositiven invasiven Tropho-blasten des Choriongürtels oder die endometrialen Cupzellen (Kydd et al., 1991). In der Plazenta von Equiden existieren zwei Populationen nicht-invasiver Trophoblasten, die sich bezüglich ihrer Morphologie und der Expression für Relaxin und MHC-Klasse I-Molekülen unterscheiden. Relaxin wird während des ovariellen Zyklus in den reifen ovariellen Follikeln sowie dem Corpus luteum exprimiert Dromedar Molekulare Charakterisierung Präprorelaxin cdna-sequenz Die Präprorelaxin cdna des Dromedars wurde aus uteroplazentarem Gewebe am Tag 300 der Trächtigkeit kloniert. Die 600 bp lange cdna (200 Aminosäuren) gliederte sich in ein Signalpeptid von 25 Aminosäuren (75 bp), eine B-Domäne von 28 Aminosäuren (84 bp), eine C-Domäne von 122 Aminosäuren (366 bp) und eine A-Domäne von 24 Aminosäuren (72 bp) (Abb. 10). Die B- Domäne enthielt das klassische Rezeptorbindungsmotiv (-GRELVR-). Das Prorelaxin des Dromedars hatte mit 122 Aminosäuren die längste C-Domäne aller bekannten Prorelaxine und resultierte aus einem bislang in keinem anderen Prorelaxin nachgewiesenen repetitiven Aminosäuremotiv (-RPAP) 3 -(K/RPAL-) 2 (Abb. 10). Das Präprorelaxin des Dromedars zeigte höchste Aminosäure-Homologie von 74.7% zum Relaxin des Schweines (Tab. 14) Eine Northern Analyse zur Bestimmung der Transkriptgröße war aufgrund der geringen Menge verfügbarer RNA nicht möglich. 46
12 Tabelle 14. Aminosäure-Homologie des Präprorelaxins und der einzelnen Relaxin-Domänen des Dromedars mit Relaxinen anderer Spezies. Spezies Signalpeptid B- C-Domäne A-Domäne Total a Domäne Schwein 79,2 88,9 78,1 54,2 74,6 Pferd 76,0 69,2 61,9 50,0 65,4 H1 b 48,0 52,0 65,4 33,3 52,7 H2 b 54,2 50,0 62,0 37,5 51,7 Katze 60,0 63,0 65,3 41,7 54,5 Hund 72,0 57,1 51,6 33,3 48,3 Ratte 54,2 33,3 55,0 41,7 45,4 Maus 50,0 37,0 58,3 29,2 39,0 a Prozentsatz identischer Aminosäuren b H1 und H2: die zwei humanen Präprorelaxine Expression und Lokalisation im Gewebe Uteroplazentares Gewebe In Plazentagewebe am Tag 93 der Gravidität wurde eine schwache und in den Plazenten am Tag 170, 196, 213, 240, 340 und 372 wurde eine starke Expression von Relaxin-mRNA und immunreaktivem Relaxin im luminalen uterinen Epithel detektiert (Abb. 11). Endometriale Drüsen und das Myometrium sowie die mit einer Digoxigenin-markierten sense crna inkubierten Plazentaschnitte zeigten keine Hybridisierungssignale (Abb. 11). Trotz des fehlenden Nachweises von Relaxin mrna konnte selbst bei einer Verdünnung des R6 Relaxin-Antiserums von 1: in den Trophoblasten der Plazentagewebe vom Tag 170 bis 372 der Gravidität immunreaktives Relaxin detektiert werden (Abb. 11). Aufgrund der Morphologie und der Expression immunreaktiven Relaxins waren in den Plazenten des Dromedars drei verschiedene Cytokeratin-positive Trophoblast-Populationen zu unterscheiden (Abb. 11). Abb. 10. Nukleinsäure- und Aminosäuresequenz des Präprorelaxins des Dromedars. Die verwendeten Primer sind durch Pfeile gekennzeichnet. Numerierung der Primer siehe Tab. 4. Die Rezeptorbindungssequenz in der B-Domäne ist kursiv gekennzeichnet. Das Aminosäuremotiv (- RPAP) 3 -(K/RPAL-) 2 im Prorelaxin des Dromedars am Beginn des N-Terminus der C-Domäne ist hervorgehobenen. 47
13 Signalpeptid 1 Met Pro Arg Leu Leu Leu Ser His Leu Leu Gly Val Trp Leu Leu Leu Ser Gln Leu Pro Lys Glu Thr 23 1 ATG CCG CGC CTG CTC TTG TCC CAC CTG CTA GGT GTC TGG CTG CTA CTG AGC CAA CTT CCT AAG GAG ACC 69 (3) B-Domäne 24 Ser Gly Glu Arg Ser Asn Asp Phe Val Lys Ala Cys Gly Arg Glu Leu Val Arg Leu Trp Ile Glu Ile TCA GGC GAG CGT TCA AAC GAC TTC GTT AAG GCA TGC GGC CGA GAA TTA GTC CGC CTC TGG ATC GAA ATC 138 (1) C-Domäne 47 Cys Gly Ser Val Ser Trp Gly Arg Pro Ala Pro Arg Pro Ala Pro Arg Pro Ala Pro Lys Pro Ala Leu TGT GGC TCT GTC TCC TGG GGG AGG CCG GCT CCG AGA CCG GCT CCG AGA CCG GCT CCA AAA CCC GCT CTG 207 (8) (9) 70 Arg Pro Ala Leu Ser Gln Asp Lys Lys Pro Arg Leu Arg Ser Gly Pro Pro Ala Glu Ile Met Pro Ser AGA CCC GCT CTG AGC CAG GAC AAG AAG CCT CGG CTG AGA TCT GGC CCG CCA GCA GAA ATC ATG CCA TCC 276 (9) (10) 93 Ser Ile Thr Lys Asp Ala Glu Thr Leu Thr Thr Met Leu Glu Phe Thr Pro Asn Leu Pro Gln Glu Leu TCC ATC ACC AAA GAT GCA GAA ACC TTA ACT ACG ATG TTG GAA TTC ACC CCT AAT TTG CCA CAG GAG CTG Thr Ala Thr Leu Ser Glu Arg Gln Pro Ser Ala Glu Pro Gln Gln Pro Ala Leu Lys Asp Ser Asn Leu ACG GCC ACA CTG TCT GAG AGG CAG CCA TCG GCA GAG CCA CAA CAA CCT GCT TTA AAG GAT TCA AAT CTT 414 (5) 139 Asn Phe Glu Glu Phe Lys Lys Ile Ile Phe Asp Arg Gln Asn Glu Glu Glu Asp Glu Ser Leu Ser Glu AAT TTC GAA GAA TTT AAG AAA ATC ATT TTT GAC AGA CAA AAT GAG GAA GAA GAC GAG AGT CTT TCA GAA 483 (6) (7) A-Domäne 162 Leu Lys Asn Leu Gly Leu Asp Lys His Ser Glu Lys Lys Arg Gln Leu Gln Met Thr Leu Gly Glu Arg TTA AAA AAC TTA GGC TTA GAT AAA CAT TCC GAA AAA AAG AGA CAG TTA CAA ATG ACA CTG GGT GAG AGA Cys Cys Gln Lys Gly Cys Ser Arg Lys Glu Met Ala Thr Ala Cys Stop TGT TGT CAA AAA GGT TGT AGC AGA AAA GAA ATG GCT ACA GCA TGC TGA 600 (2) (4) 48
14 Neben kuboidalen villösen Trophoblasten, die schwach immunreaktives Relaxin exprimierten (Abb. 11), wurden multinukleäre Trophoblast-Riesenzellen sowie häufig gegenüber der Öffnung uteriner Drüsen gruppierte elongierte Trophoblasten identifiziert (Abb. 11), die beide kein Relaxin exprimierten. Mit den zur Verfügung stehenden Antikörpern gelang bislang kein Nachweis von MHC Klasse I-Molekülen in Trophoblasten der Dromedar-Plazenta. Die Kontrolle der Spezifität der immunhistochemischen Färbungen erfolgte durch Ausschluß des Primärantikörper und ergab in allen Fällen keine spezifische Färbung (Abb. 11) Ovar In den untersuchten Ovarien vom Tag 93, 170, 213, 240, 340 und 372 der Gravidität erfolgte der spezifische Nachweis von immunreaktivem Relaxin, nicht jedoch von Relaxin mrna, in den follikulären Granulosazellen und Theca interna-zellen (Abb. 12). In den Lutealzellen der Corpora lutea aller Graviditätsstadien wurden Relaxin Transkripte sowie immunreaktives Relaxin detektiert (Abb. 12) Zusammenfassung Das Präprorelaxin des Dromedars ist das erste in einem Wiederkäuer klonierte und bislang größte Relaxin. Das Prorelaxin des Dromedars besitzt im N-Terminus der C-Domäne das neuartige Aminosäuremotiv (-RPAP) 3 -(K/RPAL-) 2. In ovariellen Follikeln und dem Corpus luteum des trächtigen Dromedars wird Relaxin im Untersuchungszeitraum vom Tag der Gravidität exprimiert. In der epitheliochorialen Plazenta des Dromedars erfolgt die Induktion des Relaxingenes im luminalen uterinen Epithel ab dem Tag 93 der Gravidität. Multinukleäre Riesenzellen und elongierte Trophoblasten exprimieren kein Relaxin. 49
 und fetales Stroma, uterinen Drüsen, fetale Trophoblasten sowie mit sense crna inkubierte Schnitte zeigten keine Hybridisierungssignale (B).")
15 Abb. 11. An plazentarem Gewebe eines Dromedars am Tag 196 (A, B) und am Tag 213 (C-F) der Gravidität erfolgte der Nachweis von Relaxin-mRNA (A) in uterinen luminalen Epithelzellen. Maternales (m) und fetales Stroma, uterinen Drüsen, fetale Trophoblasten sowie mit sense crna inkubierte Schnitte zeigten keine Hybridisierungssignale (B). Die Relaxin-mRNA-positiven uterinen Epithelzellen exprimierten auch immunreaktives Relaxin (C, E). Eine schwache Färbung für immunreaktives Relaxin wurde in fetalen Trophoblasten der plazentaren Villi (v) detektiert (D, E). Areolae-artige Areale elongierter Trophoblasten (Pfeile) sowie multinukleäre Riesenzellen (nicht gezeigt) waren immunnegativ für Relaxin (E). Das uterine luminale Epithel sowie die fetalen Trophoblasten waren immunpositiv für Cytokeratin (F). Die immunhistochemische Negativkontrolle zeigte keine Färbung (D). Vergrößerungen: A, B, E, F x132; C; D x
 sowie Theca interna (ti; A) und im Corpus luteum in großen Luteinzellen (E) nachgewiesen.")
16 Abb. 12. In einem Bouin-fixierten ovariellen Follikel sowie Corpus luteum eines Dromedars am Tag 213 der Gravidität erfolgte der Nachweis immunreaktiven Relaxins. Im Follikel wurde immunreaktives Relaxin in den Zellen der Granulosa (g) sowie Theca interna (ti; A) und im Corpus luteum in großen Luteinzellen (E) nachgewiesen. Der Nachweis von Relaxin-mRNA gelang im Corpus luteum (C). Mit Digoxigenin-markierter sense-crna wurden keine Hybridisierungssignale detektiert (D). Die immunhistochemischen Negativkontrollen für den ovariellen Follikel (B) und das Corpus luteum (F) ergaben keine Färbung. Vergrößerungen: A, B x109; C-F x
17 Die Halbaffen G. crassicaudatus und V. variegata Molekulare Charakterisierung Präprorelaxin cdna-sequenzen Die cdna-sequenzen für die Präprorelaxine von Galago crassicaudatus und Varecia variegata wurden aus mrna von Plazenten ausgetragener Trächtigkeiten kloniert. Aufgrund der geringen isolierten RNA-Mengen kam die Capfinder cdna-synthese mit anschließender RT- und RACE- PCR zum Einsatz. Die cdna-sequenzen für die Präprorelaxine beider Halbaffenspezies waren bis auf eine konservative Transversion (V.variegata-Sequenz: A 507 C; Ser 169 ) identisch und kodierten für ein Peptid von 188 Aminosäuren (567 bp) mit einem Signalpeptid mit 21 Aminosäuren (63 bp), einer B-Domäne mit 34 Aminosäuren (102 bp), einer C-Domäne mit 109 Aminosäuren (327 bp) und einer A-Domäne mit 24 Aminosäuren (75 bp; Abb. 13). Die Rezeptorbindungssequenz in der B-Domäne beider Präprorelaxine (-GRRLIR-) wich in zwei Positionen (E R 40 und V I 42 ) vom klassischen Motiv (-GRELVR-) ab. Potentielle enzymatische Prozessierungsstellen waren Ala 22 zwischen dem Signalpeptid und der B-Domäne, Arg 55 zwischen B- und C-Domäne und die -K-R- R-K- ( ) Prozessierungssequenz für Furin. Die Relaxinsequenzen beider Halbaffen zeigten höchste Aminosäurehomologie (50,6%) zum Präprorelaxin des Schweines (Tab. 15). Tabelle 15. Homologievergleich der Aminosäuresequenzen der Präprorelaxine sowie deren Domänen von Galago crassicaudatus mit Präprorelaxinen anderer Tierspezies. Spezies Signalpeptid B- C-Domäne A-Domäne Total a Domäne Schwein 70,0 50,0 47,6 37,5 50,6 Dromedar 66,7 46,2 42,5 29,2 43,7 Katze 66,7 26,9 54,1 33,3 41,1 Ratte 42,9 40,6 44,8 41,7 41,4 Maus 52,4 2,9 39,8 37,5 34,2 Hund 66,7 29,6 14,0 41,7 33,9 a Prozentsatz identischer Aminosäuren Abb. 13. cdna-sequenz und Aminosäuresequenz der Präprorelaxine von Galago crassicaudatus und Varecia variegata. Die Domänen und Primerbindungsstellen sind durch Pfeile gekennzeichnet sind. Die Primernumern entsprechen denen in Tab. 7. Das Rezeptorbindungsmotiv in der B-Domäne und die konservative Transversion in Position 507 in der cdna-sequenz für das Präprorelaxin von V. variegata (A C; Ser 169 ) sind hervorgehoben. 52
18 Signalpeptid 1 Met Pro Arg Leu Phe Phe Phe His Leu Leu Gly Val Trp Leu Leu Leu Thr Gln Ile Ser Arg Ala Lys 23 1 ATG CCT CGC CTG TTT TTT TTC CAC CTG CTA GGA GTC TGG CTG CTA CTG ACC CAA ATT TCC AGA GCA AAA 69 (1;3) (5) B-Domäne 24 Met Asp Lys Gly Glu Asn Leu Asn Gln Ile Ile Phe Ala Cys Gly Arg Arg Leu Ile Arg Ile Trp Val ATG GAT AAG GGG GAG AAT CTG AAC CAA ATT ATT TTT GCA TGT GGC CGC CGA CTA ATA CGC ATT TGG GTT 138 (6) C-Domäne 47 Glu Val Cys Gly Ser Thr Gly Phe Arg Gly Arg Ala Lys Asn Gln Thr Glu His Gln Pro Gly Ser Glu GAG GTT TGC GGC TCT ACA GGT TTC AGA GGA AGA GCT AAG AAC CAG ACA GAG CAC CAA CCT GGA TCT GAA Pro Phe Ser Glu Ile Val Pro Ser Ser Phe Ile Asn Lys Asp Ala Glu Thr Ile Asn Met Met Ser Glu CCT TTC TCA GAA ATT GTG CCA TCA TCC TTC ATC AAC AAA GAT GCA GAA ACT ATA AAT ATG ATG TCA GAA 276 (8) 93 Phe Ile Ala Asn Leu Pro Gln Lys Gln Lys Thr ThrGln Ser Glu Met Asn Leu Pro Ser Pro Glu Leu TTC ATT GCT AAT TTA CCA CAG AAG CAG AAG ACA ACACAG TCT GAG ATG AAT CTA CCT TCA CCA GAG CTA Gln Gln Tyr Pro Pro Thr Leu Lys Gly Ser Asp Ile Ser Phe Glu Glu Val Lys Asn Asn Ile His Asn CAG CAA TAT CCA CCC ACC TTA AAG GGT TCA GAT ATT AGC TTT GAA GAA GTT AAG AAT AAT ATT CAT AAT 414 (9) 139 Glu Gln Gly Glu Ala Glu Asp Asn Ser His Ser Glu Leu Gln Asn Leu Gly Leu Asp Thr His Ser Arg GAA CAA GGT GAA GCA GAA GAC AAC AGT CAT TCA GAA TTA CAA AAC TTA GGA TTG GAT ACT CAT TCC CGA 483 (7) (10) A-Domäne 162 Lys Lys Arg Glu Arg Tyr Met Ser Pro Leu Gln Lys Cys Cys Arg Ile Gly Cys Thr Lys Arg Ser AAA AAG AGA GAG CGC TAT ATG TCA/C CCA TTG CAG AAA TGT TGC CGA ATT GGT TGT ACA AAA AGA TCT 549 (2) (4) 184 Leu Ala Arg Phe Cys stop CTT GCT AGA TTT TGC TGA 565 (4) 53
19 Ein Homologievergleich mit Relaxinsequenzen anderer Primatenspezies ergab höchste Homologie von 58,4% bzw. 56,8% Homologie für die Präprorelaxine H2 des Menschen und des Rhesusaffen. Eine deutlich geringere Aminosäurehomologie von 44,9% bestand zum Präprorelaxin des Neuweltaffen Callithrix jacchus (Marmoset). Die Aminosäuresequenzen der Signalpeptide und der A-Domänen waren in den untersuchten Präprorelaxinen der Primaten am stärksten und die der B- Domäne am geringsten konserviert (Tab. 16). Tabelle 16. Homologievergleich der Aminosäuresequenzen der Präprorelaxine und ihrer Domänen von Galago crassicaudatus mit den Präprorelaxinen anderer Primaten. Spezies Signalpeptid B- C-Domäne A-Domäne Total a Domäne H1 b 71,4 40,7 53,9 58,3 50,3 H2 b 85,7 34,5 57,4 62,5 58,4 Rhesus 81,0 40,7 57,4 62,5 56,8 Marmoset 76,2 34,5 54,8 50,0 44,9 C1 c - 33,3 52,9 58,3 - C2 c - 33,3 57,4 68,2 - G1 d - 41,4-58,3 - G2 d - 37,9-62,5 - a Prozentsatz identischer Aminosäuren b H1 und H2: die zwei humanen Präprorelaxine c C1 und C2: die beiden Relaxine des Chimpansen d G1 und G2: die beiden Relaxine des Gorillas - Sequenz nicht bekannt Die phylogenetischen Analysen der Aminosäuresequenzen des Präprorelaxins von Galago crassicaudatus erfolgte mit den Präprorelaxinen des Menschen (H1 und H2), des Rhesusaffen und des Marmoset sowie den Prorelaxinen (C1 und C2) des Chimpansen. Die Präprorelaxine des Pferdes und des Schweines dienten als Referenzgruppen. Grundlage für die Erstellung des Phylogrammes war das Alignment der Relaxinsequenzen in Abb. 14. Im Vergleich zu den Relaxinsequenzen der anderen Alt- und Neuweltprimaten zeigten die Präprorelaxine des lorisoiden G. crassicaudatus und des Lemuren V. variegata eine hohe Aminosäure-Austauschfrequenz durch erhöhte Mutationsrate (Abb. 15). 54
20 Abb. 14. Alignment der Aminosäuresequenzen der Präprorelaxine des Halbaffen Galago crassicaudatus mit dem Neuweltaffen Callithrix jacchus (Callith; Marmoset) und den Präprorelaxinen der Altweltprimaten Mensch (H1 und H2) und Rhesusaffe (Rhesus) sowie den Prorelaxinen des Chimpansen (Chimp1 und Chimp2) für die Erstellung des Phylogrammes. Die einzelnen Relaxindomänen sind gekennzeichnet. Die Rezeptorbindungssequenz in der B-Domäne des Relaxins ist rot und die konservierten Cysteine sind grün markiert. Gelb hervorgehobene Aminosäuren sind bei allen Relaxinsequenzen identisch. Signalpeptid B-Domäne C-Domäne Galago MPRLFFFHLLGVWLLLTQISRAKMDKGENLNQIIFACGRRLIRIWVEVCGSTGFRGRAKNQTEHQPGSEPFSEIVPSSFINKDAETINMMSEF Human1 MPRLFLFHLLEFCLLLNQFSRAVAAKW--KDDVIKLCGRELVRAQIAICGMSTWSKRSLSQEDAPQTPRPVAEIVPS-FINKDTETIIIMLEF Human2 MPRLFFFHLLGVCLLLNQFSRAVADSW--MEEVIKLCGRELVRAQIAICGMSTWSKRSLSQEDAPQTPRPVAEIVPS-FINKDTETINMMSEF Callith MSRLFLFHLLGVCLLLNQIFRAVATNG--KDDVIKVCGRELVRAQIDACGMSTVDKSTLSREDASPQPELVTEIVPS-FNSKDTETINMMSEL Rhesus MPRLFLFHLLGVCLLLNQFSRAVAAKW--MDDVIKACGRELVRAQIAICGKSTLGKRSLNQEDAPLKPRPAAEIVPS-LINQDTETINMMSEF Chimp RAVADSW--MDEVIKLCGRELVRAQIAICGKSTWSKRSLSQEDAPQTPRPVAEIVPS-FINKDTETINMMSEF Chimp RAVADSW--MDEVIKLCGRELVRAQIAICGMSTWSKRSLSQEDAPQTPRPVAEIVPS-FINKDTETIIIMLEF C-Domäne A-Domäne Galago IANLPQKQKTTQSEMNLPSPELQQYPPTLKGSDISFEEVKNNIHNEQGEAEDNSHSELQNLGLDTHSRKKRERYMSPLQKCCRIGCTKRSLARFC Human1 IANLPPELKAALSERQPSLPELQQYVPALKDSNLSFEEFKKLIRNRQSEAADSNPSELKYLGLDTHSQKKRRPYVALFEKCCLIGCTKRSLAKYC Human2 VANLPQELKLTLSEMQPALPQLQQHVPVLKDSSLLFEEFKKLIRNRQSEAADSSPSELKYLGLDTHSRKKRQLYSALANKCCHVGCTKRSLARFC Callith IANLPQELKAAPSGRQSLLPELQQHVPILKDSDLSSEEFKEIIRNRQSEAADSSPSELKYLGLHAHSRRKRQFSLALYNQCCLIGCTKRALAEFC Rhesus VANLPQELKLTLSERQPALSELQQHVPVLKDSNLSFEEFKKIIRKRQSEATDSSPSELRSLGLDTHSRRKRQLYMTLSNKCCHIGCTKKSLAKFC Chimp1 IANLPPELKAALSERQPSLPEPQQYVPALKDSNLSFEEFKKLIRNRQSEAADSNPSELKYLGLDTHSQKKRQPYVALFEKCCLIGCTKRSLANYC Chimp2 VANLPQELKLTLSEMQPALPQLQQYVPVLKDSSLLFEEFKKLIRNRQSEAADSSPSELKYLGLDTHSRKKRQLYSALANKCCHVGCTKRSLARFC 55
21 Abb. 15. Phylogramm der Präprorelaxine von G. crassicaudatus (Galago), Rhesusaffe (Rhesus), Callithrix jacchus (Callith; Marmoset), Mensch (Human 1+2) und den Prorelaxinen des Chimpansen (Chimp 1+2). Die Präprorelaxine des Schweines und Pferdes dienten als Referenzsequenzen. Die Berechnung der Aminosäure-Substitutionen erfolgte mit der Jones-Matrix (Jones et al., 1992). Die Zweiglänge ist proportional zur Anzahl der Aminosäuresubstitutionen in dieser Sequenz. Die Länge 0,1 entspricht 1 Aminosäure-substitution pro 10 Aminosäuren. Die prozentuale Wahrscheinlichkeit für die korrekte Lage der Knotenpunkte im Phylogramm sind an den Aufzweigungen angegeben. Schwein Pferd Galago 100 Rhesus 100 Callith Chimp Human1 77 Chimp Human2 56
22 Expression und Lokalisation im Gewebe Plazentares Gewebe An Kryostatschnitten einer Plazenta von Galago crassicaudatus gelang der Nachweis von Präprorelaxin Transkripten (Abb. 16A) und immunreaktivem Relaxin (Abb. 16C) in fetalen Zellen, welche die plumpen Chorioallantoiszotten bedeckten. Die Relaxin-exprimierenden Zellen waren immunpositiv für Cytokeratin und wurden als Trophoblasten identifiziert (Abb. 16D). Die Stromazellen der plazentaren Zotten exprimierten immunreaktives Vimentin aber kein Relaxin (nicht gezeigt). Mit Digoxigenin-markierter sense crna behandelte Schnitte zeigten keine Hybridisierungssignale (Abb. 16B). Keine spezifische immunhistochemische Färbungen war nach Ausschluß des Primärantikörpers nachweisbar (Abb. 16E) Zusammenfassung Die Aminosäuresequenzen des Präprorelaxins des in Afrika und Asien beheimateten Galago crassicaudatus und des nur in Madagaskar vorkommenden Lemuren Varecia variegata sind identisch und zeigen eine phylogenetisch entfernte Verwandtschaft mit den Altweltaffen Afrikas. Relaxin wird in fetalen Trophoblasten der epitheliochorialen Plazenta von Galago crassicaudatus exprimiert. Abb. 16. An Kryoschnitte einer ausgetragenen Plazenta von Galago crassicaudatus erfolgte der Nachweis von Relaxin mrna (A) in Zellen des fetalen Villus (v). Das villöse Stroma und mit Digoxigenin-markierter sense crna inkubierte Schnitte zeigten keine Hybridisierungs-signale (B). In den Relaxin mrna-positiven villösen Zellen erfolgte der Nachweis immunoreaktiven Relaxins mit dem polyklonalen Relaxin-Antiserum R6 (C). Die gleichen Zellen wurden durch den Nachweis immunoreaktiven Cytokeratins (D) als Trophoblasten der epitheliochorialen Galago-Plazenta identifiziert. Eine typische immunohistochemische Negativkonrolle ist in (E) gezeigt. Vergrößerung: A-E x216 57
23 58
24 Katze Molekulare Charakterisierung Präprorelaxin cdna-sequenz Die cdna für das Präprorelaxin der Katze wurde aus mrna von uteroplazentarem Gewebe am Tag 35 der Trächtigkeit durch RT- und RACE-PCR kloniert. Mit einem degenerierten Oligonukleotid-Primerpaar (Tab. 5) wurde zunächst ein 430 bp Fragment des Prorelaxins amplifiziert. Die komplette Kodierungssequenz des Präprorelaxins der Katze betrug 540 bp und gliederte sich in ein Signalpeptid von 25 Aminosäuren (75 bp), eine B-Domäne von 33 Aminosäuren (99 bp), eine C-Domäne von 98 Aminosäuren (294 bp) und eine A-Domäne von 24 Aminosäuren (72 bp; Abb. 17). Das Rezeptorbindungsmotiv in der B-Domäne des Katzenrelaxins wich in einer Position (Leu Phe 37 ) von der konservierten Peptidsequenz (-GRELVR-) ab. Das Präprorelaxin der Katze zeigte höchste Homologie zu den Präprorelaxinen des Pferdes und des Schweines (Tab. 17; Klonisch et al., 1999a). Tabelle 17. Homologie des Präprorelaxins der Katze und der einzelnen Domänen mit Relaxinen anderer Spezies. Spezies Signalpeptid B- C-Domäne A-Domäne Total a Domäne Schwein 60,0 54,5 59,2 45,8 56,6 Dromedar 60,0 63,0 65,3 41,7 54,5 H1 b 52,0 48,5 50,0 37,5 48,3 H2 b 52,0 54,5 50,0 41,7 50,0 Hund 80,0 48,9 52,2 66,7 58,3 Pferd 60,0 54,0 51,2 53,4 56,6 Ratte 40,0 27,3 39,8 41,7 37,8 Maus 48,0 36,4 43,9 41,7 42,8 a Prozentsatz identischer Aminosäuren b H1 und H2: die zwei humanen Präprorelaxine Abb. 17. Die Nukleinsäure- und kodierende Peptidsequenz für das Präprorelaxin der Katze. Die Primerbindungsstellen sind durch Pfeile markiert und das Rezeptorbindungsmotiv in der B-Domäne ist hervorgehoben. Numerierung der Primer siehe Tab
25 Signalpeptid 1 Met Leu Arg Leu Phe Leu Ser His Leu Leu Gly Val Trp Leu Leu Leu Ser Leu Arg Ala Arg Lys Ile 23 1 ATG CTG CGC CTG TTC TTA TCC CAC CTG CTG GGA GTC TGG CTG CTA CTG AGC CTA CGT GCC AGA AAG ATC 69 (3) B-Domäne 24 Pro Ala Gln Glu Glu Val Leu Lys Ala Cys Gly Arg Glu Phe Val Arg Leu Gln Ile Arg Ile Cys Gly CCA GCC CAG GAG GAA GTG TTA AAA GCA TGC GGC CGA GAG TTT GTC CGA CTA CAA ATC CGG ATC TGC GGC 138 (1) (5) C-Domäne 47 Ser Leu Ser Trp Gly Lys Ser Ser Gln Gln His Arg Glu Pro Arg Gln Ala Pro Ala Ala Leu Pro Glu TCA TTG TCC TGG GGA AAA AGC AGT CAA CAA CAC CGG GAA CCT CGG CAG GCA CCC GCC GCA CTC CCA GAA 207 (5) (6) 70 Ile Val Ser Ser Ser Ile Thr Ser Gly Ala Glu Ala Leu Asn Gly Met Leu Glu Tyr Ile Pro Asp Leu ATC GTG TCC TCC TCC ATC ACC AGT GGT GCA GAA GCC TTG AAT GGG ATG TTG GAA TAC ATT CCT GAT TTG Pro Gln Glu Leu Lys Ala Thr Leu Ser Glu Arg Glu Pro Ser Phe Arg Glu Leu Gln Pro Ser Leu Lys CCA CAA GAG CTG AAG GCA ACA CTG TCT GAG AGG GAG CCA TCA TTC AGA GAA CTA CAA CCT TCA TTG AAG Asp Ser Asn Leu Asn Leu Glu Glu Val Glu Lys Ser Ile Leu Gly Arg Gln Asn Glu Ala Glu Asp Gln GAT TCT AAT CTT AAC TTG GAA GAA GTG GAG AAA AGT ATT CTT GGT AGA CAA AAT GAA GCT GAA GAC CAA 414 (7) 139 Ser Leu Ser Gln Leu Gly Arg Ser Arg Leu Asp Ala His Ser Arg Ile Lys Arg Ser Asp Tyr Ile Arg AGT CTT TCA CAA TTA GGA AGA TCA AGG TTA GAT GCA CAT TCT CGA ATA AAG AGA TCA GAC TAT ATA AGA 483 (8) A-Domäne 162 Tyr Ser Asp Arg Cys Cys Asn Val Gly Cys Thr Arg Lys Glu Leu Ala Asp Leu Cys stop TAC AGT GAC AGG TGT TGT AAC GTA GGT TGT ACC AGG AAA GAG CTT GCT GAT TTA TGC TGA 543 (2) (4) 60
26 Abb 18. Northern Analyse zum Nachweis spezifischer Präprorelaxin-Transkripte der Katze in uteroplazentarem Gewebe am Tag 35 der Gravidität. Ein spezifisches Hybridisierungsignal bei 1 kb wurde mit der Northern 1 2 Analyse in Gesamt-RNA uteroplazentaren Gewebes am Tag 35 der Gravidität (1) nachgewiesen, während Gesamt-RNA aus Lebergewebe keine Hybridisierungssignale zeigte (2; Abb. 18). 28 S 18 S 1 kb Expression und Lokalisation im Gewebe Uteroplazentares Gewebe In-situ Hybridisierungen mit einer Digoxigenin-markierten antisense crna-sonde für Präpro-relaxin wurden auf Kryostatschnitten von Katzenplazenten am Tag 35 der Gravidität durchge-führt. Im lamellären Chorioallantois-Anteil der zonaren Gürtelplazenta wurden spezifische Hybridisierungsignale für Relaxin detektiert (Abb. 19A, E, G). Die paraplazentaren uterinen Areale, die Zellen der plazentaren Transitionszone, die uterinen Drüsen, das Myometrium sowie mit sense crna behandelte Gewebeschnitte zeigten keine Hybridisierungssignale (Abb. 19B). Die villösen, Relaxin mrna-exprimierenden Zellen wurden aufgrund ihrer immunpositiven Färbung für Cytokeratin als Trophoblasten identifiziert (Abb. 19F, H). Trophoblasten an den fetalen Zottenspitzen, die in der plazentaren Transitionszone lokalisiert waren und das maternale Endometrium invadierten exprimierten kein Relaxin (Abb. 19A, G). Extravillöse Trophoblasten, die sich von der Zottenspitze losgelöst hatten und im endometrialen Stroma lokalisiert waren, exprimierten ebenfalls keine Relaxin mrna (Abb. 19H). Mit dem Relaxin-Antiserum R6 gelang der Nachweis von immunreaktivem Relaxin in den Relaxin mrna-exprimierenden villösen Trophoblastzellen des plazentaren Labyrinths (Abb. 19C). Die Trophoblastzellen der invadierenden Zottenspitzen sowie die extravillösen Trophoblasten zeigten keine Immunreaktivität für Relaxin. Experimente zur Kontrolle der Spezifität der durchgeführten immunhistochemischen Färbungen erfolgten unter Ausschluß der entsprechenden Primärantikörper und zeigten keine Färbung (Abb. 19D, I). 61
27 Zusammenfassung Die cdna für das Präprorelaxin der Katze besitzt die für alle bekannten Relaxine charakteristischen Struktur ( N Signalpeptid-B-C-A C ). In der Katzenplazenta wird Relaxin ausschließlich von villösen Trophoblasten im lamellären Labyrint der Gürtelplazenta exprimiert. Invadierende Trophoblasten an der fetalen Zotten-spitze sowie extravillöse Trophoblasten im maternalen Endometrium exprimieren kein Relaxin. Abb. 19. An Gefrierschnitten uteroplazentaren Gewebes einer Katze am Tag 35 der Gravidität erfolgte der spezifische Nachweis von Relaxin Transkripten mit der nichtradioaktiven in-situ Hybridisierung (A, B, E, G). Der Nachweis von immunreaktivem Relaxin mit einem polyklonalen Antiserum aus dem Kaninchen (R6; C) und von immunreaktivem Cytokeratin mit dem Maus monoklonalen Antikörper MNF-116 (F, H) erfolgte mit der Kaninchen- bzw. Maus-APAAP indirekten Immunhistochemie. Die Trophoblasten in den Villi (v) des lamellären plazentaren Labyrints (l) exprimierten spezifisch Relaxin-mRNA (A, E, G). Die Zellen in der Transitionszone (t), die uterinen Drüsen (u), das Myometrium (m), die paraplazentaren uterinen Areale sowie die mit der Digoxigenin-markierten sense crna inkubierten Schnitte zeigten keine Hybridisierungssignale (A, B). Die Relaxin-mRNA-exprimierenden Zellen waren immunpositiv für Cytokeratin (F, H) und wurden als villöse Trophoblasten identifiziert. Trophoblasten an der Spitze der invadierenden Villi (v) in der Transitionszone (t) exprimierten immunoreaktives Cytokeratin (Pfeile; H) aber keine Relaxin Transkripte (A, G). Immunoreaktives Relaxin wurde im plazentaren Labyrint (l) lokalisiert, in denen auch Relaxin-mRNA-exprimierende Trophoblasten nachweisbar waren (C). Typische immunhistochemische Negativkontrollen für die Kaninchen- und Maus-APAAP Methode wurden unter Ausschluß des entsprechenden Primärantikörpers durchgeführt und ergaben keine Färbung (D, I). Vergrößerungen: A, B x29; E, F x145; C, D, G-I x

28 63
29 Hund Molekulare Charakterisierung Präprorelaxin cdna-sequenz Die cdna für das Präprorelaxin des Hundes wurde aus mrna uteroplazentaren Gewebes am Tag 35 der Trächtigkeit kloniert. Mit degenerierten Oligonukleotidprimern, die entsprechend der Aminosäuresequenz für die A- und B-Domäne des Hunderelaxins (Stewart et al.,1992b) hergestellt waren (Tab. 6), gelang die Amplifikation eines 424 bp Fragmentes für das Prorelaxin. Die cdna- Sequenz des Präprorelaxins betrug 531 bp und kodierte für ein Signalpeptid von 25 Aminosäuren (75 bp), eine B-Domäne von 35 Aminosäuren (105 bp), eine C-Domäne von 93 Aminosäuren (279 bp) und eine A-Domäne von 24 Aminosäuren (72 bp; Abb. 20). Die aus der cdna-sequenz abgeleiteten Aminosäuresequenzen für die A- und B-Domäne waren identisch mit den publizierten Sequenzen für das Hunderelaxin (Stewart et al., 1992b). Das Rezeptorbindungsmotiv der B- Domäne zeigte mit Glu Asp 37 und Leu Tyr 38 zwei Substitutionen im klassischen GRELVR-Motiv. Das Präprorelaxin des Hundes zeigte höchste Homologie zum Präprorelaxin des Schweines (Tab. 18; Klonisch et al., 1999b). Tabelle 18. Homologie des Präprorelaxins des Hundes und der einzelnen Domänen mit Relaxinen anderer Spezies. Spezies Signalpeptid B- C-Domäne A-Domäne Total a Domäne Schwein 72,0 51,4 50,0 41,7 52,3 Dromedar 72,0 57,1 51,6 33,3 48,3 H1 b 52,0 34,3 37,0 45,8 39,8 H2 b 52,0 31,4 34,8 50,0 38,6 Katze 80,0 48,9 52,2 66,7 58,3 Pferd 64,0 39,9 48,9 54,2 50,0 Ratte 44,0 42,9 33,7 41,7 38,1 Maus 52,0 34,3 31,5 45,8 36,9 a Prozentsatz identischer Aminosäuren b H1 und H2: die zwei humanen Präprorelaxine Abb. 20. Nukleinsäure- und kodierende Aminosäuresequenz für das Präprorelaxin des Hundes. Die Domänen des Präprorelaxins sowie die Primerbindungsstellen sind durch Pfeile markiert (siehe auch Tab 6). Das potentielle Rezeptorbindungsmotiv in der B-Domäne ist hervorge-hoben. 64
30 Signalpeptid 1 Met Leu Arg Trp Phe Leu Ser His Leu Leu Gly Val Trp Leu Leu Leu Ser Gln Leu Pro Arg Glu Ile 23 1 ATG CTG CGC TGG TTC TTG TCC CAC CTG CTA GGT GTC TGG CTG CTA CTA AGC CAA CTT CCC AGA GAG ATC 69 (3) B-Domäne 24 Pro Ala Thr Asp Asp Lys Lys Leu Lys Ala Cys Gly Arg Asp Tyr Val Arg Leu Gln Ile Glu Val Cys CCA GCC ACG GAT GAC AAG AAA CTT AAG GCA TGT GGT CGT GAT TAT GTC CGC CTA CAG ATT GAG GTC TGC 138 (5) (1) (6) (7) C-Domäne 47 Gly Ser Ile Trp Trp Gly Arg Lys Ala Gly Gln Leu Arg Glu Arg Arg Gln Ile Ser Glu Pro Leu Ala GGC TCC ATC TGG TGG GGG AGA AAG GCT GGC CAG CTG CGG GAA CGT CGG CAG ATA TCC GAA CCC CTG GCA 207 (7) 70 Glu Val Val Pro Ser Ser Ile Ile Asn Asp Pro Glu Ile Leu Ser Leu Met Leu Gln Ser Ile Pro Gly GAA GTT GTG CCA TCC TCC ATC ATC AAT GAT CCA GAA ATC TTA AGT TTG ATG TTG CAA TCC ATT CCC GGT 276 (9) 93 Met Pro Gln Glu Leu Arg Ile Ala Thr Arg Ser Gly Lys Glu Lys Leu Leu Arg Glu Leu His Phe Val ATG CCA CAG GAA CTG AGG ATA GCA ACA CGG TCT GGG AAG GAG AAG TTA TTA AGA GAG CTA CAC TTT GTA 345 (9) (8) 116 Leu Glu Asp Ser Asn Leu Asn Leu Glu Glu Met Lys Lys Thr Phe Leu Asn Thr Gln Phe Glu Ala Glu CTG GAA GAT TCC AAT CTT AAC TTG GAA GAA ATG AAG AAA ACT TTT CTT AAC ACA CAA TTT GAA GCT GAA 414 A-Domäne 139 Asp Lys Ser Leu Ser Lys Leu Asp Lys His Pro Arg Lys Lys Arg Asp Asn Tyr Ile Lys Met Ser Asp GAC AAA AGC CTT TCA AAA TTA GAT AAA CAT CCC CGA AAA AAG AGA GAT AAC TAC ATA AAA ATG AGT GAT Lys Cys Cys Asn Val Gly Cys ThrArg Arg Glu Leu Ala Ser Arg Cys stop AAA TGT TGT AAT GTA GGT TGT ACCAGA AGA GAG CTT GCT AGC CGA TGC TGA 528 (2) (4) 65
31 Abb. 21. Northern Analyse zum Nachweis spezifischer Präprorelaxin-Transkripte im uteroplazentaren Gewebe des Hundes. 1 2 Ein spezifisches Hybridisierungssignal von 1 kb wurde in der Northern Analyse mit Gesamt-RNA aus uteroplazentarem Gewebe am Tag 35 der Gravidität (2), nicht jedoch aus der Niere (1), nachgewiesen (Abb. 21). 28 S 18 S 1 kb Expression und Lokalisation im Gewebe Uteroplazentares Gewebe Uteroplazentares Gewebe am Tag 30 und 35 der Gravidität von je zwei Hündinnen diente zur Gewebelokalisation der Relaxin-Transkripte. Spezifische Hybridisierungssignale waren auf die labyrintäre zonare Gürtelplazenta beschränkt (Abb. 22A, B). Die paraplazentaren uterinen Anteilen, die plazentare Transitionszone, das Myometrium sowie mit Digoxigenin-markierter sense crna inkubierte Gewebeschnitte zeigten keine Hybridisierungssignale (Abb. 22C). Das maternale Endometrium invadierende fetale Zotten waren von einer basalen Schicht isoprismatischer Zellen umgeben, die keine Relaxin-mRNA exprimierten (Abb 22A, B). Diese Zellen waren immunpositiv für Cytokeratin und wurden als Cytotrophoblasten identifiziert (Abb. 22G). Diese Cytotrophoblasten waren von einer dichte Zellschicht umgeben, die eine starke spezifische Hybridisierung für RelaxinmRNA zeigte (Abb. 22A, B). In den plazentaren Paraffinschnitten waren starke Hybridisierungssignale für Relaxin-mRNA an der Spitze der fetalen Villi und schwächere netzartige Hybridisierungssignale an der Basis der Villi nachweisbar (Abb. 22A). In den entsprechenden Kryoschnitten waren die netzartigen Hybridisierungssignale deutlicher sichtbar. Die Relaxin-mRNAexprimierenden Zellen waren ebenfalls immunpositiv für Cytokeratin aber immunnegativ für Vimentin und wurden als Syncytiotrophoblasten identifiziert (Abb. 22G, H). Mit dem für Hunderelaxin spezifischen Kaninchen-Antiserum No (Steinetz et al., 1996) gelang der Nachweis immunreaktiven Relaxins in den Relaxin-mRNA-exprimierenden Syncytiotrophoblasten (Abb. 22E). Mit einem Antikörper gegen von-willebrand-faktor, einem Markermolekül für vaskuläre 66
32 Endothelzellen, konnte eine intensive Vaskularisation im Bereich des Relaxin-sezernierenden Syncytiotropho-blasten immunhistochemisch nachgewiesen werden (Abb. 22I). Kontrollexperimente unter Ausschluß des Primärantikörpers ergaben keine spezifische Färbungen (Abb. 22F) Zusammenfassung Das Präprorelaxin des Hundes kodiert für 177 Aminosäuren und besitzt zwei Aminosäureaustausche in der Rezeptorbindungssequenz der B-Domäne. In der zonaren Gürtelplazenta des Hundes exprimiert der Syncytiotrophoblast, nicht jedoch der Cytotrophoblast, Präprorelaxin. Im Bereich der Relaxin-sezernierenden Syncytiotrophoblasten war ein ausgedehntes kapilläres Gefäßnetz nachweisbar. Abb. 22. Bouin-fixiertes und paraffiniertes uteroplazentares Gewebe einer Hündin am Tag 30 der Trächtigkeit diente dem Nachweis von Relaxin-mRNA mit der nichtradioaktiven in-situ Hybridisierung (A-C). Mit einer Kaninchen- oder Maus-APAAP indirekten Immunhistochemie wurden immunreaktives Relaxin mit einem polyklonalen Antiserum spezifisch für Hunderelaxin (Nr ; D-E), Cytokeratin (G) oder Vimentin (H) mit einem monoklonalen Mausantikörper und von-willebrand Faktor (I) mit einem polyklonalen Kaninchen Antiserum detektiert. Spezifische Hybridisierungssignale für Relaxin mrna wurden im Syncytio-trophoblast der fetalen Villi (v) des plazentaren Labyrints nachgewiesen (A, B). Der Cytotrophoblast der fetalen Villi (A, B), das maternale Endometrium, die paraplazentaren uterinen Areale sowie die mit Digoxigenin-markierter sense crna inkubierten Gewebeschnitte zeigten keine Hybridisierungssignale für Relaxin mrna (C). Das Cytokeratin-positive (G) und Vimentin-negative (H) fetale Syncytium exprimierte auch immunreaktives Relaxin (D, E). Endometriale Stromazellen zeigten eine starke Expression für immunreaktives Vimentin (nicht gezeigt), während vaskuläres Endothel eine schwache Färbung aufwies (Pfeil; H). Mit dem Nachweis immunreaktiven von-willebrand Faktors gelang die Darstellung eines dichten kapillären Netzwerk im Syncytium der Hundeplazenta (I). Eine typische immunhistochemische Negativkontrolle ist in (F) dargestellt. Maßstab: A, C, D, F, H, I: x131; B, E, G: x

33 68
34 Zusammenfassung der Ergebnisse der Relaxin-Studien Die in dieser Arbeit klonierten cdna-moleküle kodieren für Relaxine mit den strukturellen Voraussetzungen für eine intakte Bioaktivität als sekretierte Hormone. Die phylogenetischen Analysen der Präprorelaxine der beiden Halbaffen Galago crassicaudatus und Varecia variegata zeigen a.) eine erstärkte Aminosäuresubstitutionsrate und folglich erhöhte Mutationsrate im Vergleich zu den Präprorelaxinen anderer Primaten und b.) deuten auf einen gemeinsamen afrikanischen Ursprung aller Primaten, einschließlich der Halbaffen, hin. Immunreaktives Relaxin wird in den ovariellen Follikeln und Relaxin-mRNA und immunreaktives Relaxin werden in Luteinzellen des Corpus luteum der nichhtträchtigen Stute und des trächtigen Dromedars exprimiert. Relaxin ist ein frühes Produkt nicht-invasiver Trophoblasten des Allantochorions im Pferdeembryo. Relaxin wird in den epitheliochorialen Plazenten der Equiden, des Dromedars und des Halbaffen Galago crassicaudatus exprimiert. Jedoch bestehen bezüglich der Relaxin-exprimierenden Zellen und des zeitlichen Expressionsmusters erhebliche Unterschiede in den verschiedenen Spezies: Equines Relaxin wird früh während der Plazentation im nicht-invasiven kuboidalen Trophoblasten der reifenden Mikrokotyledonen nachgewiesen. MHC Klasse I-immunpositive invasive equine Trophoblasten und elongierte Trophoblasten exprimieren kein Relaxin. Die Relaxin-Expression in der epitheliochorialen beginnt beim Dromedar erst um den Tag 93 der Gravidität in uterinen luminalen Epithelzellen. Fetale kuboidale Trophoblasten exprimieren schwach immunreaktives Relaxin. Elongierte Trophoblasten der Areolae und multinukleäre Riesenzellen exprimieren kein Relaxin. In einer postpartalen Plazenta von Galago crassicaudatus gelang der Relaxin-Nachweis in fetalen villösen Trophoblasten. In den endotheliochorialen Plazenten der Katze und des Hundes wird Relaxin ausschließlich in fetalen Trophoblasten des plazentaren Anteils der Gürtelplazenta, nicht jedoch in den paraplazentaren und uterinen Regionen exprimiert. In der Katzenplazenta exprimieren Cytotrophoblasten Relaxin, während in der Hundeplazenta Relaxin ein frühes Sekretions-produkt des Syncytiotrophoblasten darstellt. 69
35 4.2. Relaxin-like Faktor (RLF) Ziege Molekulare Charakterisierung RLF cdna Die 396 bp lange cdna für den Relaxin-like faktor (RLF) der Ziege wurde aus mrna eines adulten Ziegenhodens kloniert und kodierte für ein Polypeptid von 131 Aminosäuren (Abb. 23). Entsprechend der potentiellen enzymatischen Prozessierungsstellen in RLF-Molekülen anderer Spezies (Ivell, 1997a), gliederte sich das Ziegen-RLF in ein Signalpeptid von 24 Aminosäuren (72 bp), eine B-Domäne von 31 Aminosäuren (93 bp), eine C-Domäne von 50 Aminosäuren (150 bp) und eine A-Domäne von 26 Aminosäuren (78 bp; Abb. 23). Die Lokalisation und Anzahl der Cysteine sowie die potentielle Rezeptorbindungssequenz RALVR in der B-Domäne waren konserviert (Abb. 23). Von allen RLF-Domänen war die Peptidsequenz der A-Domäne am stärksten konserviert (Abb. 23). Ziegen-RLF zeigte 98% Homologie mit den RLF-Sequenzen anderer Widerkäuer und eine geringere Homologie zum RLF des Schweines (86%) und des Menschen (70%) (Tab. 19; Hombach-Klonisch et al., 1999). Tabelle 19. Homologievergleich der Aminosäuresequenz des Ziegen-RLF mit RLF-Molekülen anderer Spezies. Höchste Homologie bestand zum RLF der Wiederkäuer Rind und Schaf. Spezies Signalpeptid B-Domäne C-Domäne A-Domäne Total a Rind 100% 93,6% 100% 100% 98,5% Schaf b 93,3% 93,6% 98,0% 96,2% 95,9% Schwein 75,0% 87,1% 87,8% 92,3% 86,3% Mensch 75,0% 71,0% 62,0% 80,8% 70,2% Marmoset 75,0% 67,7% 58,0% 80,0% 67,7% Galago 66,7% 74,2% 68,0% 80,8% 71,8% Maus 53,3% 67,7% 47,8% 61,5% 55,7% a Prozent identischer Aminosäuren b keine komplette Signalpeptidsequenz kloniert (Roche et al., 1996) 70
36 Abb. 23. cdna und kodierende Aminosäuresequenz für den Relaxin-like Faktor (RLF) der Ziege. Die Einzeldomänen des RLF sowie die Primerbindungsstellen sind durch Pfeile gekennzeichnet (siehe auch Tab. 8). Das potentielle Rezeptorbindungsmotiv in der B-Domäne ist hervorgehoben. Signalpeptid 1 Met Asp Arg Arg Pro Leu Thr Trp Ala Leu Val Leu Leu Gly Pro Ala Leu Ala Ile Ala Leu Gly 22 1 ATG GAC CGT CGT CCG CTC ACC TGG GCT CTG GTG CTG CTG GGC CCG GCC CTT GCA ATC GCC CTC GGT 66 (3) (1) B-domain 23 Pro Ala Ala Ala Gln Glu Ala Pro Glu Lys Leu Cys Gly His His Ser Val Arg Ala Leu Val Arg CCT GCA GCC GCG CAG GAG GCG CCG GAG AAA CTG TGT GGC CAC CAC TCC GTG CGC GCG CTC GTG CGG 132 (5) (6) C-domain 45 Leu Cys Gly Gly Pro Arg Trp Ser Ser Glu Ser Gly Arg Pro Val Ala Gly Gly Asp Arg Glu Leu CTG TGC GGC GGC CCG CGC TGG TCT TCC GAG TCC GGG CGA CCT GTG GCT GGC GGC GAC CGT GAG CTC 198 (6) 67 Leu Arg Trp Leu Glu Gly Gln His Leu Leu His Gly Leu Met Ala Ser Gly Asp Pro Val Leu Val CTA CGG TGG CTG GAA GGA CAA CAT CTC CTC CAT GGG CTG ATG GCC AGT GGG GAC CCC GTG CTG GTA 264 (7) A-domain 89 Leu Ala Pro Gln Pro Leu Pro Gln Ala Ser Arg His His His His Arg Arg Ala Thr Ala Ile Asn CTG GCC CCA CAG CCC CTG CCC CAG GCT TCT CGC CAT CAC CAC CAC CGC CGT GCA ACT GCC ATC AAC 330 (7) (8) 111 Pro Ala Arg His Cys Cys Leu Ser Gly Cys Thr Arg Gln Asp Leu Leu Thr Leu Cys Pro His stop CCT GCC CGC CAC TGC TGC CTC AGC GGC TGC ACC CGG CAA GAC CTG CTG ACC CTC TGT CCC CAC TGA 396 (9) (2,4) 71
.")
37 Abb. 24. Northern Analyse für Ziegen-RLF. Northern Analyse mit einer Digoxigenin-markierten antisense crna für Ziegen-RLF zeigte ein spezifisches Hybridisierungssignal von 0.9 kb mit Gesamt-RNA des Hodens (1). RNA des Nebenhodens (2), der Leber (3), des Muskels (4) sowie mit einer Digoxigenin-markierten sense crna für Ziegen-RLF inkubierte Northern Blots ergaben keine Hybridisierungssignale. Identische Ergeb-nisse wie in Abb. 24 wurden mit Gesamt-RNA von entsprechenden Damwild-Geweben erzielt (nicht ge-zeigt). 28S 18S 0.9 kb Genomische Struktur Southern Analyse einer mit den Restriktionsenzymen HindIII (1) oder EcoRI (2) verdauten genomischen DNA ergab nach Inkubation mit einem 32 P-markierten PCR-Fragment für Ziegen- RLF spezifische Hybridisierungssignale bei 4,5 kb (HindIII) und 3 kb (EcoRI; Abb. 25). Southern Analysen genomischer DNA des Damwildes mit einem 32 P-markierten Damwild-RLF PCR-Fragment ergaben identische Ergebnisse (nicht gezeigt). Die Ziege und das Damwild besitzten somit nur ein RLF-Gen. 4.5 kb 3.0 kb 1 2 Abb. 25. Southern Blot genomischer DNA der Ziege Expression und Lokalisation im Gewebe Hoden In Hoden von drei 27 Monate alten geschlechtsreifen Ziegenböcken wurden spezifische Hybridisierungssignale für RLF-Transkripte in den interstitiellen Leydigzellen nachgewiesen (Abb. 72

38 27A-C). Die Zellen der Tubuli seminiferi und des Nebenhodens sowie Hodenschnitte, die mit der Digoxigenin-markierten sense crna für Ziegen-RLF behandelt wurden, zeigten keine Hybridisierungssignale (Abb. 27E). Die Leydigzellen waren immunpositiv für die zwei Markermoleküle des Steroidstoffwechsels, 3β-Hydroxysteroid Dehydrogenase (3β-HSD) und 17α-Hydroxylase (Abb. 27D). Kontrollexperimente zur Spezifität der durchgeführten immunhistochemischen Färbungen erfolgten unter Ausschluß der entsprechenden Primär-antikörpers und ergaben keine Färbung (Abb. 27F) Uteroplazentares Gewebe Mit spezifischen Primern für Ziegen-RLF (Tab. 8) gelang der RT-PCR Nachweis eines 396 bp langen Amplifikates in Ziegen-Plazenta zum Zeitpunkt der Geburt (Abb. 26). Das Amplifikat wurde durch Sequenzanalyse als RLF der Ziege identifiziert. Abb. 26. RT-PCR Nachweis von RLF-mRNA in der Plazenta der Ziege. Nachweis eines spezifischen Amplifikates für RLF in cdna einer Ziegenplazenta (1). Eine RT-PCR mit plazentarer Gesamt-RNA anstelle von cdna als Template diente als Negativkontrolle. 100 bp- Marker (3). 396 bp Zusammenfassung Die RLF-Sequenz der Ziege ist, wie die RLF-Sequenzen anderer Widerkäuer, hoch konserviert. Die Ziege besitzt nur ein Gen für RLF. Ziegen-RLF wird in den testikulären Leydigzellen und in der Plazenta exprimiert. Abb. 27. An Paraffinschnitten des adulten Ziegenhodens erfolgte der spezifische Nachweis von RLF-Transkripten mit nichtradioaktiver in-situ Hybridisierung in interstitiellen Leydig-zellen (A-C). Mit einer sense crna inkubierte Schnitte zeigten keine Hybridisierungssignale (E). Die Leydigzellen wurden durch den Nachweis des immunreaktiven 3β-HSD charakteri-siert (D). Eine typische immunhistochemische Negativkontrolle ist in (F) gezeigt. Vergrößerungen: A, E, F x132; B, C, D x

39 74
40 Damwildes (Dama dama) Molekulare Charakterisierung RLF cdna und genomische Struktur Die RLF-cDNA des Damwildes wurde aus testikulärer mrna eines adulten Damwildbockes kloniert und kodierte für ein Polypeptid von 131 Aminosäuren (396 bp) mit einem Signalpeptid von 24 Aminosäuren (72 bp), einer B-Domäne von 31 Aminosäuren (93 bp), einer C-Domäne von 50 Aminosäuren (150 bp) und einer A-Domäne von 26 Aminosäuren (78 bp; Abb. 28). Damwild-RLF zeigte ca. 96% Nukleinsäure- und Aminosäurehomologie (Tab. 20) zu den RLF Molekülen der Widerkäuer Rind, Schaf und Ziege. Die RLF-Peptidsequenzen der Wiederkäuer zeigte eine Sequenzheterogenität an der Grenze zwischen der B- und C-Domäne, wobei das RLF des Rindes eine zusätzliche Glutaminsäure (Glu 55 ) enthielt (Abb. 29). Identisch zu den Northern Analysen bei der Ziege war ein spezifisches Hybridisierungssignal bei 0,9 kb in Gesamt-RNA des Hodens nachweisbar, während der Nebenhoden, die Leber und der Muskel keine RLF-Transkripte enthielten (siehe Abb. 24). Southern Analyse einer mit den Restriktionsenzymen HindIII (1) oder EcoRI (2) verdauten genomischen DNA ergab nach Inkubation mit einem 32 P-markierten PCR- Fragment für Damwild-RLF spezifische Hybridisierungssignale bei 4,5 kb (HindIII) und 3 kb (EcoRI). Diese Ergebnisse waren identisch zu den Southern Analysen bei der Ziege (siehe Abb. 25). Tabelle 20. Homologievergleich der Aminosäuresequenz des Damwild-RLF mit RLF-Molekülen anderer Spezies. Höchste Homologie bestand zum RLF der Widerkäuer Rind und Schaf. Spezies Signalpeptid B-Domäne C-Domäne A-Domäne Total a Rind 95,8% 96,8% 100% 100% 96,9% Ziege 95,8% 90,3% 98,0% 100% 96,2% Schaf b 86,7% 96,8% 98,0% 96,2% 95,1% Schwein 75,0% 90,3% 87,8% 92,3% 86,3% Mensch 70,8% 74,2% 64,0% 80,8% 71,0% Marmoset 70,8% 71,0% 56,0% 80,0% 77,7% Maus 53,3% 64,5% 47,8% 61,5% 64% a Prozent identischer Aminosäuren b keine komplette Signalpeptidsequenz vorhanden 75
41 Abb. 28. cdna und kodierende Aminosäuresequenz für das RLF des Damwildes. Die Einzeldomänen des RLF und die Primerbindungsstellen sind durch Pfeile gekennzeichnet (siehe auch Tab. 8). Das potentielle Rezeptorbindungsmotiv in der B-Domäne ist hervorgehoben. Signalpeptid 1 Met Asp Arg Arg Pro Leu Thr Trp Ala Leu Val Leu Leu Gly Pro Ala Leu Ala Ile Ala Leu Ser 22 1 ATG GAC CGT CGT CCG CTC ACC TGG GCC CTG GTG CTG CTG GGC CCG GCC CTT GCG ATC GCC CTC AGT 66 (3) (1) (5) B-Domäne 23 Pro Ala Ala Ala Gln Glu Val Pro Glu Lys Leu Cys Gly His His Phe Val Arg Ala Leu Val Arg CCT GCA GCC GCG CAG GAG GTG CCT GAG AAA CTG TGT GGC CAC CAC TTC GTG CGC GCG CTC GTG CGG132 (5) (6) C-Domäne 45 Leu Cys Gly Gly Pro Arg Trp Ser Ser Glu Asp Arg Arg Pro Val Ala Gly Gly Asp Arg Glu Leu CTG TGC GGC GGC CCG CGC TGG TCT TCC GAG GAC CGG CGA CCT GTG GCT GGC GGC GAC CGT GAG CTC198 (6) 67 Leu Arg Trp Leu Glu Gly Gln His Leu Leu His Gly Leu Met Ala Ser Gly Asp Pro Val Leu Val CTA CGG TGG CTG GAA GGA CAA CAT CTC CTC CAT GGG CTG ATG GCC AGT GGG GAC CCT GTG CTG GTT264 (7) A-Domäne 89 Leu Ala Pro Gln Pro Leu Pro Gln Ala Ser Arg His His His His Arg Arg Ala Thr Ala Ile Asn CTG GCC CCA CAA CCC CTG CCC CAG GCT TCT CGC CAT CAC CAC CAC CGC CGG GCA ACT GCC ATC AAC330 (7) (8) 111 Pro Ala Arg His Cys Cys Leu Ser Gly Cys Thr Arg Gln Asp Leu Leu Thr Leu Cys Pro His & CCT GCC CGC CAC TGC TGC CTC AGC GGC TGC ACC CGG CAA GAC CTG CTG ACC CTC TGT CCC CAC TGA396 (9) (2;4) 76
42 Abb. 29. Schematische Darstellung des Homologievergleiches der bislang klonierten RLF-Moleküle von Widerkäuern. Identische Aminosäuren sind durch einen Punkt gekennzeichnet. B-Domäne Ziege Damwild Rind Schaf AAQEAPEKLCGHHSVRALVRLCGGPRWSSES-...V...F...-D...F...ED...F...-D C-Domäne Ziege GRPVAGGDRELLRWLEGQHLLHGLMASGDPVLVLAPQPLPQASRHHHHRR Damwild R... Rind... Schaf...A... A-Domäne Ziege ATAINPARHCCLSGCTRQDLLTLCPH Damwild... Rind... Schaf...V Expression und Lokalisation im Gewebe Hoden Spezifische Hybridisierungssignale für RLF-mRNA wurden in den interstitiellen Leydigzellen des adulten Damwildhodens lokalisiert, die auch immunoreaktive 3β-HSD und 17α-Hydroxylase exprimierten. Die Zellen der Tubuli seminiferi, der Nebenhoden und mit Digoxigenin-markierte sense crna inkubierte Schnitte zeigten keine Hybridisierungssignale. Die Ergebnisse zur testikulären RLF- Expression im Damwild waren identisch mit denen aus dem Ziegenhoden (siehe Abb. 27) Ovar In den ovariellen Follikeln dreier adulter Tiere exprimierten die Theca interna-zellen RLF- mrna (Abb. 30A). Die Expression war variabel, da nicht in allen Follikeln RLF-Transkripte detektiert wurden. Die Luteinzellen der Corpora lutea exprimierten ebenfalls RLF-mRNA (Abb. 30C). Die mit 77
. Abb. 30.")
43 Digoxigenin-markierter sense RLF-cRNA inkubierten Schnitte zeigten keine Hybridisierungssignale (Abb. 30B, F). Abb. 30. Mit einer Digoxigenin-markierten antisense crna für das Damwild-RLF erfolgte der Nachweis spezifischer Hybridisierungssignale in den Zellen der Theca interna (ti; A) des Ovars und den Luteinzellen des Corpus luteum (C). Granulosazellen (g), ovariellen Stromazellen sowie mit einer sense crna behandelte Schnitte zeigten keine Hybridisierungssignale (B). In uteroplazentarem Gewebe am Tag 80 der Gravidität wurden RLF-Transkripte in den endometrialen Drüsen nachgewiesen (D). Im nichtträchtigen Uterus wurden spezifische Hybridisierungssignale für RLF-mRNA im luminalen uterinen Epithel sowie in vornehmlich basal lokalisierten endometrialen Drüsen (Pfeile) detektiert (E). Mit einer sense-crna inkubierte Schnitte zeigten keine spezifischen Hybridisierungssignale (B, F). Vergrößerungen: A-D x216; E, F x
44 Uterus und uteroplazentares Gewebe Im nichtträchtigen Uterus und in interplazentaren Uterusarealen wurde RLF-mRNA im uterinen Epithel und in den vornehmlich basal liegenden uterinen Drüsen detektiert (Abb. 30 E). In Plazenten am Tag 80 und 110 der Trächtigkeit war RLF-mRNA sowohl in den uterinen Drüsen (Abb. 30D), im luminalen Epithel und in fetalen Trophoblasten nachweisbar (Abb. 31A, B). Fetale Stromazellen exprimierten ebenfalls RLF-Transkripte, während das maternale Stroma frei von RLF-mRNA war. Die in den fetalen Villi lokalisierten kuboidalen, uninukleären Trophoblasten exprimierten schwach RLF-mRNA sowie immunreaktives Cytokeratin (Abb. 31A, B, D). Binukleäre Trophoblasten (BNC) wurden mit den beiden Antikörpern gegen ovines plazentares Laktogen (opl; Abb. 31E) und Pregnancy-Specific-Protein-60 (PSP-60; nicht gezeigt) identifiziert. Die basal dem fetalen Zottenstroma aufliegenden BNC zeigten eine starke Expression für RLF-Transkripte, während die apikal dem uterinen Epithel anliegenden BNC häufig keine RLF-mRNA exprimierten (Abb. 31A, B). Elongierte, Cytokeratin-positive Trophoblasten (Abb. 31G) an der Basis der fetalen Zotten exprimierten schwach RLF Transkripte. Mit dem monoklonalen Antikörper 87G (Blaschitz et al., 1997) gelang der Nachweis immunreaktiver MHC Klasse Ib-Antigene auf der Oberfläche der elongierten Trophoblasten (Abb. 31H). Kontrollexperimente mit einer Digoxigenin-markierten sense crna zeigten keine Hybridisierungssignale (Abb. 31C). Der Ausschluß der entsprechenden Primärantikörper diente als Negativkontrolle und ergab keine Färbung (Abb. 31H, I). Abb. 31. In Damwild-Plazenten am Tag 80 (B, D, E, F) und am Tag 120 (A, C, G-I) der Gravidität erfolgte der Nachweis von RLF-mRNA im uterinen luminalen Epithel der maternalen plazentaren Septen (m) sowie in uni- und binukleären Trophoblasten (BNC; Pfeile) entlang der plazentaren Villi (v; A) und in den Zottenspitzen (B). RLF-mRNA-exprimierende BNC waren vornehmlich basal (A, B) und BNC mit Expression für immunreaktives plazentares Laktogen vor allem apikal (E) im villösen Zottenepithel lokalisiert. Elongierte Trophoblasten an der Basis der plazentaren Villi (I) exprimierten schwach RLF-mRNA (nicht gezeigt). Elongierte Trophoblasten (G) und villöse uninukleäre Trophoblasten, nicht jedoch BNC (D), exprimierten immunreaktives Cytokeratin. Ausschließlich elongierte Trophoblasten exprimierten immunreaktiven MHC Klasse Ib Moleküle (H). Die mit sense crna inkubierten Schnitte zeigten keine Hybridisierungssignale (C). Immunhistochemische Negativkontrollen zeigten ebenfalls keine Färbung (F, I). Vergrößerungen: A-I x

45 Zusammenfassung Im Hoden ist RLF-mRNA ein spezifisches Transkript adulter testikulärer Leydigzellen. RLF-mRNA wird im nichtträchtigen Uterus und im uteroplazentaren Gewebe exprimiert. Binukleäre Trophoblasten exprimieren stark RLF-mRNA. Elongierte Trophoblasten exprimieren RLF-mRNA und präsentieren MHC Klasse Ib- Antigene auf ihrer Zelloberfläche. Die Theca interna Zellen und die Granulosazellen der ovariellen Follikel sowie die Luteinzellen des Corpus luteum exprimieren RLF-Transkripte. 80
46 Galago crassicaudatus Molekulare Chrakterisierung RLF cdna Die RLF-cDNA wurde aus Hodengewebe eines geschlechtsreifen Galago crassicaudatus kloniert und kodierte für ein RLF-Peptid von 131 Aminsäuren (396 bp), das sich in ein Signalpeptid von 24 Aminosäuren (72 bp), eine B-Domäne von 31 Aminosäuren (93 bp), eine C-Domäne von 50 Aminosäuren (150 bp) und eine A-Domäne von 26 Aminosäuren (78 bp) (Abb. 32) gliederte. Das RLF von G. crassicaudatus enthielt die konservierte potentielle Rezeptorbindungssequenz RALVR in der B-Domäne (Abb. 32). RT-PCR ergab Amplifikate für RLF im Hoden, aber nicht in der Plazenta von G. crassicaudatus (nicht gezeigt). Höchste Aminosäurehomologie bestand zu den beiden Primaten-RLF Sequenzen des Menschen (79,4%) und des Neuweltaffen Callithrix jacchus (74,6%; Tab. 21). Northern-Analysen waren wegen der geringen zur Verfügung stehenden RNA- Mengen nicht möglich. Tabelle 21. Homologievergleich der Aminosäuresequenz des RLF von Galago crassicaudatus mit RLF Molekülen anderer Spezies. Höchste Homologie bestand zum RLF der Menschen. Spezies Signalpeptid B-Domäne C-Domäne A-Domäne Total a Mensch 83,3% 80,6% 76,0% 80,8% 79,4% Marmoset 83,3% 77,4% 68,0% 76,0% 74,6% Schwein 58,3% 80,6% 64,6% 88,5% 72,5% Rind 66,7% 77,4% 67,3% 80,8% 70,2% Ziege 66,7% 74,2% 68,0% 80,8% 71,8% Schaf b 66,7% 77,4% 67,3% 80,8% 73,0% Maus 26,7% 77,4% 56,5% 73,1% 61,5% a Prozent identischer Aminosäuren b keine komplette Signalpeptidsequenz vorhanden 81
47 Abb. 32. cdna und kodierende Aminosäuresequenz für das RLF von Galago crassicaudatus. Die Einzeldomänen des RLF und die Primerbindungsstellen sind durch Pfeile gekennzeichnet. Primer (1) bindet teilweise und Primer (8) vollständig ausserhalb des RLF-kodierenden cdna-bereiches (Numerierung siehe Tab. 9). Nur die innerhalb des kodierenden cdna-bereiches liegenden Primerbindungsstellen sind dargestellt. Das potentielle Rezeptorbindungsmotiv in der B-Domäne ist hervorgehoben. Signalpeptid 1 Met Asp Pro Arg Leu Ser Val Trp Ala Leu Val Leu Leu Gly Pro Ala Leu Val Phe Ala Leu His 22 1 ATG GAC CCC CGT CTG TCC GTC TGG GCG CTG GTG CTG CTG GGT CCT GCC CTA GTT TTC GCG CTG CAC 66 (3;5) (6) (1) B-Domäne 23 Pro Ser Leu Ser Leu Glu Thr Arg Glu Lys Leu Cys Gly His His Phe Val Arg Ala Leu Val Arg CCT AGT CTT TCC CTG GAG ACG CGG GAG AAG TTG TGT GGC CAC CAC TTC GTG CGC GCT CTG GTG CGC132 (7) C-Domäne 45 Leu Cys Gly Gly Pro Arg Trp Ser Pro Glu Ala Gly Thr Ser Ser Ala Gly Gly Asp Arg Glu Leu CTG TGC GGT GGC CCG CGC TGG TCC CCC GAG GCG GGG ACA TCC TCA GCT GGC GGT GAC CGT GAA CTG Leu Gln Trp Leu Glu Arg Pro His Leu Leu His Gly Leu Val Ala Glu His Asp Pro Ala Leu Val CTA CAG TGG CTG GAG AGA CCA CAT CTG CTC CAT GGG TTG GTG GCT GAG CAT GAC CCT GCA TTG GTA264 A-Domäne 89 Pro Gly Leu Gln Pro Leu Pro Gln Ala Ser His His His Arg His His Arg Ala Ala Thr Thr Asn CCT GGC CTG CAA CCC CTC CCC CAG GCT TCT CAC CAT CAT CGC CAT CAC CGG GCA GCT ACC ACC AAC Pro Ala His Arg Cys Cys Leu Ser Gly Cys Thr Arg Gln Asp Leu Leu Thr Leu Cys Pro His & CCT GCA CAC CGC TGC TGT CTC AGT GGC TGC ACC CGA CAA GAC CTG CTG ACC CTC TGT CCC CAC TGA396 (2;4) 82
48 Genomische Organisation Southern Analyse einer mit den Restriktionsenzymen EcoRI (1) oder HindIII (2) verdauten genomischen DNA ergab nach Inkubation mit dem 32 P-markierten RLF-Fragment spezifische Hybridisierungssignale bei 1,9 kb (EcoRI) und 13 kb (HindIII; Abb. 33). 13 kb 1 2 Abb. 33. Southern Analyse genomischer DNA von G. crassicaudatus. 1.9 kb Eine phylogenetische Analysen der RLF-Moleküle von G. crassicaudatus, dem Menschen und dem Neuweltaffen Callithrix jacchus (Abb. 34) deuteten auf einen gemeinsamen evolutionsbiologischen Ursprung dieser Primaten. Von den drei Primatensequenzen zeigte das RLF von G. crassicaudatus die höchste Anzahl an Aminosäuresubstitutionen. Abb. 34. Phylogramm der Peptidsequenzen aller bislang klonierten RLF-Moleküle. Das RLF des Schweines diente als Referenzsequenz. Die Berechnung der Aminosäure-Substitutions-matrix erfolgte nach Jones et al. (1992). Die Zweiglänge ist proportional zur Anzahl der Aminosäuresubstitutionen in dieser Sequenz, wobei eine Aminosäuresubstitution pro zehn Aminosäuren der angegebenen Zweiglänge von 0,1 entspricht. Die Zahl an der Verzweigungs-stelle entspricht der prozentualen Wahrscheinlichkeit für die korrekte Positionierung der Verzweigung im Phylogramm. 83
49 Expression und Lokalisation im Gewebe Hoden In der cdna aus testikulärer RNA, nicht jedoch aus plazentarer RNA, eines G. crassicaudatus wurde ein spezifisches Amplifikat für RLF isoliert und sequenziert (Abb. 32). Mit der nichtradioaktiven in-situ Hybridisierung erfolgte der Nachweis von RLF-Transkripten in interstitiellen testikulären Leydigzellen (Abb. 35a) in Hodengewebe dreier geschlechtsreifer Lemuren der in Madagaskar endemischen Spezies Varecia variegata. Das kryokonservierte testikuläre Gewebe von G. crassicaudatus eignete sich für diese Untersuchungen nicht. Die Zellen der Tubuli seminiferi, des Nebenhodens und die mit Digoxigenin-markierter sense RLF-cRNA inkubierten Gewebe- 84
50 schnitte zeigten keine Hybridisierungssignale (Abb. 35b). Die testikulären Leydigzellen waren immunpositiv für 17α-Hydroxylase (Abb. 35c). Abb. 35. In Bouin-fixiertem Hodengewebe dreier geschlechtsreifer Lemuren der Spezies Varecia variegata erfolgte der spezifische Nachweis von RLF-Transkripten in interstitiellen Leydigzellen (A). Das Keimepithel, die Zellen des Nebenhodens sowie mit Digoxigenin-markierter sense-crna inkubierte Paraffinschnitte (B) zeigten keine Hybridisierungssignale. Die RLF-mRNAexprimierenden interstitiellen testikulären Leydigzellen wurden durch den Nachweis von 17α- Hydroxylase (C) weiter charakterisiert. Die immunhistochemischen Negativkontrollen unter Ausschluß des Primärantikörpers ergaben keine Färbung (D). Vergrößerungen: A-D x216. A A B C C D D Zusammenfassung G. crassicaudatus besitzt nur ein Gen für RLF. Das Phylogramm zeigt eine relativ hohe Aminosäure-Substitutionsrate mit erhöhter Mutationsrate für das RLF von G. crassicaudatus. In der Lemurenspezies V. variegata und sehr wahrscheinlich bei allen Halbaffen ist RLF-mRNA spezifisch in adulten testikulären Leydigzellen exprimiert. Die Plazenta von G. crassicaudatus exprimiert keine RLF-Transkripte. 85
51 Mensch (Homo sapiens) Expression und Lokalisation im Gewebe Testikuläre Leydigzell-Neoplasien Der Nachweis von RLF-mRNA im humanen Hodengewebe erfolgte mit einer Digoxigeninmarkierten antisense crna eines für humanes RLF spezifischen 550 bp PCR-Fragment. Im normalen adulten menschlichen Hoden zeigten ausschließlich interstitielle Leydigzellen ein spezifisches Hybridisierungssignal (Abb. 36; 1). Humane testikuläre Leydigzell-Hyperplasien zeigten einen graduellen Verlust der RLF-Expression von der Peripherie zum Zentrum der Hyperplasie (Abb. 36; 3, 5), während benachbart gelegene nicht hyperplastische Leydigzellen eine normale Expression für RLF-mRNA zeigten. In den beiden untersuchten humanen Leydigzell-Adenomen waren RLF- Transkripte nur in Leydigzellen der Randzone, nicht jedoch im Zentrum der Adenome, nachweisbar (Abb. 36; 3; Klonisch et al., 1999c). Das Keimepithel der Tubuli seminiferi sowie die mit der Digoxigenin-markierten sense crna behandelten Hodenschnitte zeigten keine Hybridisierung (nicht gezeigt). Der Nachweis von immunreaktivem humanem RLF mit dem polyklonalen Rattenantiserum RA 15 (Ivell et al., 1997b) stimmte mit dem in der in-situ Hybridisierung ermittelten Expressionsmuster für RLF-mRNA überein (Abb. 36; 2, 4, 6). Das Expressionsprofil für immunreaktives 3β-HSD, einem Markermolekül für Leydigzellen, entsprach der RLF-Expression im Tumorgewebe (nicht gezeigt). Die RLF-Expression in den Leydigzell-Neoplasien beruhte nicht auf einem veränderten Proliferationsverhalten dieser Leydigzellen, da von diesen weniger als 1% den nukleären Proliferationsmarker Ki-67 exprimierten. Kontrollexperimente unter Ausschluß des primären Antikörpers ergaben keine Immunfärbung (nicht gezeigt). Abb. 36. Nichtradioaktive in-situ Hybridisierung mit einer Digoxigenin-markierten antisense-crna Sonde für humanes RLF im normalen humanen Hoden (1), einem Leydigzell-Adenom (3) und einer Leydigzell-Hyperplasie (5). Während Leydigzellen des normalen Hodens RLF-mRNA exprimierten (1), war in den Leydigzell-Neoplasien eine starke Abnahme der RLF-Transkriptmenge von der Peripherie zum Zentrum nachweisbar (3, 5). Die Ergebnisse des immunhistochemischen Nachweises von RLF mit dem Antikörper RA 15 im normalen Hoden (2), einem Leydigzell-Adenom (4) und einer Leydigzell-Hyperplasie (6) waren vergleichbar den Daten aus der in-situ Hybridisierung. Maßstab: 1, 2, 5, 6: 15 µm; 3, 4: 25 µm. 86
.")
52 Ovarieller Sertoli-Leydigzell-Tumor (SLCT) Der spezifische Nachweis von RLF erfolgte in Leydigzellen zweier Patientinnen mit virilisierendem ovariellem Sertoli-Leydigzell-Tumor (Abb. 37 A, C). In den ovariellen Leydig-zellnestern wurden sowohl mit der nichtradioaktiven in-situ Hybridisierung sowie beim immunhistochemischen RLF- Nachweis mit dem polyklonalen Rattenantiserum RA 15 (Ivell et al., 1997b) RLF-positive und RLFnegative Leydigzellen nachgewiesen. Die ovariellen Leydig-zellen wurden durch ihre typische Histologie und den spezifischen Nachweis immunreaktiver 3β-HSD und 17α-Hydroxylase identifiziert (Abb. 37 E, G). RLF-negative ovarielle Leydigzellen exprimierten weder immunreaktive 3β-HSD noch 17α-Hydroxylase. Eine kombinierte in-situ Hybridisierung für RLF mit anschließender Immunhistochemie für Cytokertain 18 zeigte, daß die RLF-mRNA-exprimierenden ovariellen Leydigzellen, im Unterschied zu RLF-positiven testikulären Leydigzellen, immunreaktives 87
. Abb. 37.")
53 Cytokeratin 18 exprimierten (Abb. 37 F; Klonisch et al., 1999d). Vergleichbar den Verhältnissen im humanen Hoden konnte der Proliferationsmarker Ki-67 nur in sehr wenigen ovariellen Leydigzellen nachgewiesen werden (nicht gezeigt; Klonisch et al., 1999d). Abb. 37. In einem ovariellen Sertoli-Leydigzell-Tumor (SLCT) erfolgte der spezifische Nachweis von RLF-mRNA (A, F) und immunreaktivem RLF (C) in der Leydigzell-Komponente. Die Sertoli Zellen, das ovarielle Stroma sowie die mit einer sense-crna Sonde für RLF inkubierten Schnitte (B) zeigten keine Hybridisierungssignale. Die Leydigzellen des SLCT wurden durch den Nachweis von immunreaktiver 3β-HSD (E) und 17α-Hydroxylase (G) charakterisiert. Der Nachweis der Koexpression von RLF-mRNA und immunreaktivem Cytokeratin 18 in den ovariellen Leydigzellen (F) gelang mit einer Kombination aus in-situ Hybridisierung und Immunhistochemie. Vergrößerungen: A, C, E, F: x550; B, D: x
54 Uteroplazentares Gewebe In Plazenten zweier nichtpathologischer Schwangerschaften der 9. und 13. SSW erfolgte der Nachweis von RLF-mRNA im villösen und extravillösen Cytotrophoblasten (Abb. 38 A, D). Bezüglich der maternalen Deciduazellen und des Syncytiotrophoblasten kann derzeit keine eindeutige Aussage zur RLF-mRNA-Expression gemacht werden. Es erscheint jedoch möglich, daß die maternale Deciduazellen schwach und der Syncytiotrophoblast keine RLF-mRNA exprimieren (Abb. 38 J). Extravillöse Cytotrophoblasten in der gesamten Invasionszone der Haftzotten und in der Decidua basalis exprimierten RLF-mRNA (Abb. 38 A, H). Auch intravaskulär gelegene Cytotrophoblasten exprimierten RLF-mRNA (Abb. 38 H). Syncytio- und Cytotrophoblasten wurden durch den Nachweis immunreaktiven Cytokeratins (Abb. 38 B, I) und die maternalen Deciduazellen durch den immunhistochemischen Nachweis von Vimentin (Abb. 38 C) charakterisiert. Mit dem monoklonalen Mausantikörper HLA-A2, der spezifisch HLA-A und HLA- G erkennt, erfolgte der Nachweis von HLA-G auf der Zelloberfläche einer Subpopulation extravillöser RLF-mRNA-exprimierender Cytotrophoblasten (Abb. 38 E). In der humanen Plazenta wird kein HLA-A präsentiert. Negativkontrollen zur in-situ Hybridisierung (Abb. 38 D, G) und Immunhistochemie (Abb. 38 F) zeigten keine Färbung. Abb. 38. In einer humanen Plazenta der 13. SSW (A-I) und 9. SSW (J) erfolgte der Nachweis von RLF-mRNA mit einer Digoxigenin-markierten antisense crna für humanes RLF in den Zotten und in der Decidua basalis (d; H). In den Haftzotten (A) waren RLF-exprimierende Zellen entlang des Villus (v) nachweisbar (A). Diese Zellen waren immunpositiv für Cytokeratin (B) und immunnegativ für Vimentin (C) und wurden als Cytotrophoblasten identifiziert. Die Expression von immunoreaktivem HLA-G war auf den extravillösen Cytotrophoblasten beschränkt (E). In der Decidua basalis (d) und im Lumen decidualer Gefäße wurden RLF-mRNA-exprimierende interstitielle sowie vaskuläre Cytotrophoblasten (vt; H) lokalisiert und durch den Nachweis immunreaktiven Cytokeratins identifiziert (I). In einer Plazenta der 9. SSW exprimierten villöse (v) Cytotrophoblasten RLF-mRNA, während der Syncytiotrophoblast (Pfeil) keine Hybridisierungssignale zeigte (J). Mit sense crna inkubierte Schnitte zeigten keine Hybridisierungssignale (D, G) und die immunhistochemischen Negativkontrollen ergaben keine Färbung (F). Vergrößerungen: A-F x109; G-J x

55 Zusammenfassung In testikulären Leydigzell-Hyperplasien und -Adenomen erfolgt eine graduelle Abnahme der Genexpression für RLF von der Peripherie zum Zentrum der Neoplasie. Leydigzellen ovarieller SLCT zeigen ein heterogenes RLF-Expressionsmuster und unterscheiden sich von testikulären Leydigzellen durch die Expression von Cytokertain 18. In der normalen humanen Plazenta der 9. und 13. SSW exprimieren villöse und extravillöse Cytotrophoblasten RLF mrna. Die RLF-Expression ist konstitutiv und unabhängig vom jeweiligen Proliferations- und Differenzierungszustand des extravillösen Cytotrophoblasten. 90
A- Zugabe von 180µl Pufferlösung und Proteinase K, Inkubation bei 56 C. B- Zugabe von 200µl einer zweiten Pufferlösung, Inkubation bei 70 C
 3 Methoden 3.1 Extraktion der DNA aus den Paraffinschnitten 3.1.1 Extraktions-Kit Das QIAamp DNA Mini Kit- System beruht auf dem Prinzip der Auflösung der Eiweiße mit Proteinase K, DNA Bindung an eine
3 Methoden 3.1 Extraktion der DNA aus den Paraffinschnitten 3.1.1 Extraktions-Kit Das QIAamp DNA Mini Kit- System beruht auf dem Prinzip der Auflösung der Eiweiße mit Proteinase K, DNA Bindung an eine
Sigma-Aldrich Chemie GmbH, München Merck Biosciences, Darmstadt. Thermo Hybaid GmbH, Ulm Roche Diagnostics GmbH, Mannheim
 3 Materialien 28 3 Materialien 3.1 Chemikalien Acrylamid Agar Agarose Ammoniumsulfat Ampicillin, Natriumsalz L-Arabinose Bradford-Reagenz Bromphenolblau Caseinhydrolysat Chloramphenicol Chloroform Coomassie-Brillant-Blau
3 Materialien 28 3 Materialien 3.1 Chemikalien Acrylamid Agar Agarose Ammoniumsulfat Ampicillin, Natriumsalz L-Arabinose Bradford-Reagenz Bromphenolblau Caseinhydrolysat Chloramphenicol Chloroform Coomassie-Brillant-Blau
Patente in der Biotechnologie. VPP-Düsseldorf. 2002-März-20
 Patente in der Biotechnologie VPP-Düsseldorf 2002-März-20 25 März, 2002 KB-RP / PL / PL4 / Biotechnology 1 Patente in der Biotechnologie Große Tiere, kleine Tiere, Gene, Proteine und Patente 25 März, 2002
Patente in der Biotechnologie VPP-Düsseldorf 2002-März-20 25 März, 2002 KB-RP / PL / PL4 / Biotechnology 1 Patente in der Biotechnologie Große Tiere, kleine Tiere, Gene, Proteine und Patente 25 März, 2002
Praktikum Molekularbiologie, 5. Semester, BT 12 Martin Sievers, Gottfried Dasen und Tobias Wermelinger RT 418
 Praktikum Molekularbiologie, 5. Semester, BT 12 Martin Sievers, Gottfried Dasen und Tobias Wermelinger RT 418 Datum Gruppe Thema 18.09.14 I Forensik: PCR und Restriktionsanalyse am Beispiel der ITS-Region
Praktikum Molekularbiologie, 5. Semester, BT 12 Martin Sievers, Gottfried Dasen und Tobias Wermelinger RT 418 Datum Gruppe Thema 18.09.14 I Forensik: PCR und Restriktionsanalyse am Beispiel der ITS-Region
Alternative Diabetesdiagnostik
 Köln 05.-07. Mai 2006 Alternative Diabetesdiagnostik Wolfgang Heiden FH Bonn-Rhein-Sieg Sankt Augustin, Germany Wolfgang Heiden wolfgang.heiden@fh-brs.de 2006 zur Person: Wolfgang Heiden wolfgang.heiden@fh-bonn
Köln 05.-07. Mai 2006 Alternative Diabetesdiagnostik Wolfgang Heiden FH Bonn-Rhein-Sieg Sankt Augustin, Germany Wolfgang Heiden wolfgang.heiden@fh-brs.de 2006 zur Person: Wolfgang Heiden wolfgang.heiden@fh-bonn
Aminosäurenanalytik. Probenvorbereitung Eiweißfällung, Oxidation und Hydrolyse Karl-Heinz Jansen SYKAM CHROMATOGRAPHIE
 Aminosäurenanalytik Probenvorbereitung Eiweißfällung, Oxidation und Hydrolyse Karl-Heinz Jansen SYKAM CHROMATOGRAPHIE Proteinfällung 2 Proteinfällung 3 Proteinfällung 4 Proteinfällung 5 Proteinfällung
Aminosäurenanalytik Probenvorbereitung Eiweißfällung, Oxidation und Hydrolyse Karl-Heinz Jansen SYKAM CHROMATOGRAPHIE Proteinfällung 2 Proteinfällung 3 Proteinfällung 4 Proteinfällung 5 Proteinfällung
Transkriptmengen und Apocarotinoidgehalte der chimären Pflanzen von. VII.4 Mykorrhizierungsgrad und Arbuskelvorkommen in Pflanzen des
 VII VII.1 VII.2 VII.3 Anhang Oligonucleotide p35suida-int Transkriptmengen und Apocarotinoidgehalte der chimären Pflanzen von Experiment IV VII.4 Mykorrhizierungsgrad und Arbuskelvorkommen in Pflanzen
VII VII.1 VII.2 VII.3 Anhang Oligonucleotide p35suida-int Transkriptmengen und Apocarotinoidgehalte der chimären Pflanzen von Experiment IV VII.4 Mykorrhizierungsgrad und Arbuskelvorkommen in Pflanzen
Molekularbiologie II. Rekombinante PCR Zielgerichtete Mutagenese Heterologe Expression His-Tag Aufreinigung BIOCHEMISCHES ANFÄNGERPRAKTIKUM
 V 1.8 Molekularbiologie II BIOCHEMISCHES ANFÄNGERPRAKTIKUM FÜR CHEMIKER UND BIOLOGEN Rekombinante PCR Zielgerichtete Mutagenese Heterologe Expression His-Tag Aufreinigung Abteilung für Biochemie und Molekularbiologie,
V 1.8 Molekularbiologie II BIOCHEMISCHES ANFÄNGERPRAKTIKUM FÜR CHEMIKER UND BIOLOGEN Rekombinante PCR Zielgerichtete Mutagenese Heterologe Expression His-Tag Aufreinigung Abteilung für Biochemie und Molekularbiologie,
Erfassung und Bewertung von Genexpressionsmustern von Zebrabärblingen (Danio rerio) nach Belastung mit östrogenen Substanzen
 nach Belastung mit östrogenen Substanzen") Lehrstuhl für Zellbiologie der Technischen Universität München Wissenschaftszentrum Weihenstephan Univ.-Prof. Dr. Bertold Hock Erfassung und Bewertung von Genexpressionsmustern von Zebrabärblingen (Danio
Lehrstuhl für Zellbiologie der Technischen Universität München Wissenschaftszentrum Weihenstephan Univ.-Prof. Dr. Bertold Hock Erfassung und Bewertung von Genexpressionsmustern von Zebrabärblingen (Danio
3. Methoden. 3.1 Chemikalien und Geräte
 3. Methoden Aufgabenstellung: Ziel der Arbeit war es eine eigene Multiplexreaktion zu erstellen, die im Rechtsmedizinischen Institut der Martin-Luther-Universität Halle-Wittenberg zur Durchführung von
3. Methoden Aufgabenstellung: Ziel der Arbeit war es eine eigene Multiplexreaktion zu erstellen, die im Rechtsmedizinischen Institut der Martin-Luther-Universität Halle-Wittenberg zur Durchführung von
8 Anhang. 8.1 Vektorkarten und verwendete Sequenzen 8.1.1 Vektorkarten. Anhang 90
 Anhang 90 8 Anhang 8.1 Vektorkarten und verwendete Sequenzen 8.1.1 Vektorkarten 8.1.1.1 Expression in Sf9-Zellen durch Baculoviren pfastbac1 mit GST (Firma Invitrogen) Anhang 91 pbsv-8his mit Signalpeptid
Anhang 90 8 Anhang 8.1 Vektorkarten und verwendete Sequenzen 8.1.1 Vektorkarten 8.1.1.1 Expression in Sf9-Zellen durch Baculoviren pfastbac1 mit GST (Firma Invitrogen) Anhang 91 pbsv-8his mit Signalpeptid
Anmeldenummer: 87113445.8 Int. Cl.4: C12N 15/00, C07H 21/04, - C07K7/36, C12N 1/20, Anmeldetag: 15.09.87 //C12P21/02.C12R1 :19
 Europaishes Patentamt i European Patent Offie Veröffentlihungsnummer: 0 261 552 Offie europeen des brevets AI EUROPÄISCHE PATENTANMELDUNG Anmeldenummer: 87113445.8 Int. Cl.4: C12N 15/00, C07H 21/04, -
Europaishes Patentamt i European Patent Offie Veröffentlihungsnummer: 0 261 552 Offie europeen des brevets AI EUROPÄISCHE PATENTANMELDUNG Anmeldenummer: 87113445.8 Int. Cl.4: C12N 15/00, C07H 21/04, -
ScieNews. ScieTalk Der bts-wissenschaftskongress. Das Magazin der bts. Juli 2013. Wissenschaft. bts aktuell
 Juli 2013 Das Magazin der bts ScieTalk Der bts-wissenschaftskongress bts aktuell Wissenschaft Ansturm auf den Schwarzwald - Die bts in Freiburg Organs on a Chip - Vorstellung des ScieTalk Siegerprojektes
Juli 2013 Das Magazin der bts ScieTalk Der bts-wissenschaftskongress bts aktuell Wissenschaft Ansturm auf den Schwarzwald - Die bts in Freiburg Organs on a Chip - Vorstellung des ScieTalk Siegerprojektes
2 Materialien. Materialien 15. 2.1 Histondeacetylaseinhibitoren (HDIs) Sigma-Aldrich, Deutschland. 2.2 Zelllinien und Zellkultur.
 Sigma-Aldrich, Deutschland. 2.2 Zelllinien und Zellkultur.") Materialien 15 2 Materialien 2.1 Histondeacetylaseinhibitoren (HDIs) Substanz Butyrat Trichostatin A (TSA) Valproat Hersteller Sigma-Aldrich, Sigma-Aldrich, Sigma-Aldrich, 2.2 Zelllinien und Zellkultur
Materialien 15 2 Materialien 2.1 Histondeacetylaseinhibitoren (HDIs) Substanz Butyrat Trichostatin A (TSA) Valproat Hersteller Sigma-Aldrich, Sigma-Aldrich, Sigma-Aldrich, 2.2 Zelllinien und Zellkultur
Aufgabe 2: (Aminosäuren)
") Aufgabe 2: (Aminosäuren) Aufgabenstellung Die 20 Aminosäuren (voller Name, 1- und 3-Buchstaben-Code) sollen identifiziert und mit RasMol grafisch dargestellt werden. Dann sollen die AS sinnvoll nach ihren
Aufgabe 2: (Aminosäuren) Aufgabenstellung Die 20 Aminosäuren (voller Name, 1- und 3-Buchstaben-Code) sollen identifiziert und mit RasMol grafisch dargestellt werden. Dann sollen die AS sinnvoll nach ihren
PATENTSCHRIFT. int.ci.6; A61K 39/395
 Europäisches Patentamt European Patent Office Office europeen des brevets Veröffentlichungsnummer: 0 548 194 B1 EUROPÄISCHE PATENTSCHRIFT Veröffentlichungstag der Patentschrift: 24.05.95 int.ci.6; A61K
Europäisches Patentamt European Patent Office Office europeen des brevets Veröffentlichungsnummer: 0 548 194 B1 EUROPÄISCHE PATENTSCHRIFT Veröffentlichungstag der Patentschrift: 24.05.95 int.ci.6; A61K
Metasystemische Ergebnisse zum Rezeptor-Protein Npr21 und dem molekularen Schalter cgki2.
 Metasystemische Ergebnisse zum Rezeptor-Protein Npr21 und dem molekularen Schalter cgki2. Verschaltung des Nervensystems Forscher auf der Spur eines Schlüsselprozesses Nervenzellen müssen sich verschalten,
Metasystemische Ergebnisse zum Rezeptor-Protein Npr21 und dem molekularen Schalter cgki2. Verschaltung des Nervensystems Forscher auf der Spur eines Schlüsselprozesses Nervenzellen müssen sich verschalten,
Funktionelle Genomik. praktische Anwendungen
 Funktionelle Genomik praktische Anwendungen Schmelzpunkt der DNA Einzelsträngige DNA absorbiert UV-Licht effektiver SYBR Green fluoresziert nur wenn es an doppelsträngiger DNA bindet Fluoreszenz erhöht
Funktionelle Genomik praktische Anwendungen Schmelzpunkt der DNA Einzelsträngige DNA absorbiert UV-Licht effektiver SYBR Green fluoresziert nur wenn es an doppelsträngiger DNA bindet Fluoreszenz erhöht
2. Material und Methoden
 2. Material und Methoden 2.1 Verwendete Materialien 2.1.1 Chemikalien und Enzyme Alle verwendeten Chemikalien wurden, sofern nicht anders angegeben, bei Sigma (München), Roth (Karlsruhe), Fluka (Buchs,
2. Material und Methoden 2.1 Verwendete Materialien 2.1.1 Chemikalien und Enzyme Alle verwendeten Chemikalien wurden, sofern nicht anders angegeben, bei Sigma (München), Roth (Karlsruhe), Fluka (Buchs,
Aminosäurebestimmung im Kochschinken
 1. Gegenstand der Untersuchung Es wurden 4 verschiedene Kochschinken eines Discounters hinsichtlich deren Gehalt an freien und NPN-gebundenen sowie möglichen Zusätzen untersucht. Darüber hinaus dienen
1. Gegenstand der Untersuchung Es wurden 4 verschiedene Kochschinken eines Discounters hinsichtlich deren Gehalt an freien und NPN-gebundenen sowie möglichen Zusätzen untersucht. Darüber hinaus dienen
Vorläufige Empfehlung des UAM zum Nachweis von SMRV in Zellinien Stand: Mai 2007
 Vorläufige Empfehlung des Unterausschuss Methodenentwicklung (UAM) der Bund/Länder-AG Gentechnik (LAG) zur Untersuchung von Zelllinien auf Kontamination mit dem Squirrel Monkey Retrovirus Aus der Überwachung
Vorläufige Empfehlung des Unterausschuss Methodenentwicklung (UAM) der Bund/Länder-AG Gentechnik (LAG) zur Untersuchung von Zelllinien auf Kontamination mit dem Squirrel Monkey Retrovirus Aus der Überwachung
Die Rolle der Autoimmunreaktion gegen das SLA/LP - Molekül bei autoimmuner Hepatitis
 Die Rolle der Autoimmunreaktion gegen das SLA/LP - Molekül bei autoimmuner Hepatitis Dissertation zur Erlangung des Grades Doktor der Naturwissenschaften am Fachbereich Biologie der Johannes Gutenberg-Universität
Die Rolle der Autoimmunreaktion gegen das SLA/LP - Molekül bei autoimmuner Hepatitis Dissertation zur Erlangung des Grades Doktor der Naturwissenschaften am Fachbereich Biologie der Johannes Gutenberg-Universität
Biologie. Ausbildungsrichtung: Agrarwirtschaft. Dienstag, 26. Mai 2009
 ABITURPRÜFUNG 2009 ZUM ERWERB DER FACHGEBUNDENEN HOCHSCHULREIFE AN FACHORSCHULEN UND RUFSORSCHULEN Biologie Ausbildungsrichtung: Agrarwirtschaft Prüfungstag: Arbeitszeit: Dienstag, 26. Mai 2009 180 Minuten
ABITURPRÜFUNG 2009 ZUM ERWERB DER FACHGEBUNDENEN HOCHSCHULREIFE AN FACHORSCHULEN UND RUFSORSCHULEN Biologie Ausbildungsrichtung: Agrarwirtschaft Prüfungstag: Arbeitszeit: Dienstag, 26. Mai 2009 180 Minuten
Universitätsklinikum Essen Medizinische Fakultät der Universität-Gesamthochschule Duisburg-Essen. Aus dem Institut für Humangenetik
 Universitätsklinikum Essen Medizinische Fakultät der Universität-Gesamthochschule Duisburg-Essen Aus dem Institut für Humangenetik Deletionen und Insertionen des RB1-Gens bei Patienten mit Retinoblastom:
Universitätsklinikum Essen Medizinische Fakultät der Universität-Gesamthochschule Duisburg-Essen Aus dem Institut für Humangenetik Deletionen und Insertionen des RB1-Gens bei Patienten mit Retinoblastom:
Genotypisierung des telogenen Haares
 Genotypisierung des telogenen Haares Inhalt 1. Genotypisierung des telogenen Haares 2. Die Entscheidung Sequenzierung des mitochondrialen Genom oder STR-Analyse 3. Das telogene Haar und seine Entwicklung
Genotypisierung des telogenen Haares Inhalt 1. Genotypisierung des telogenen Haares 2. Die Entscheidung Sequenzierung des mitochondrialen Genom oder STR-Analyse 3. Das telogene Haar und seine Entwicklung
Optimierte konstante Domänen für die effektive Expression von Fab-Fragmenten in E. coli
 Aus dem Institut für Biochemie der Medizinischen Fakultät Charité Universitätsmedizin Berlin DISSERTATION Optimierte konstante Domänen für die effektive Expression von Fab-Fragmenten in E. coli zur Erlangung
Aus dem Institut für Biochemie der Medizinischen Fakultät Charité Universitätsmedizin Berlin DISSERTATION Optimierte konstante Domänen für die effektive Expression von Fab-Fragmenten in E. coli zur Erlangung
MOL.504 Analyse von DNA- und Proteinsequenzen
 MOL.504 Analyse von DNA- und Proteinsequenzen Kurs 1 Monika Oberer, Karl Gruber MOL.504 Modul-Übersicht Einführung, Datenbanken BLAST-Suche, Sequenzalignment Proteinstrukturen Virtuelles Klonieren Abschlusstest
MOL.504 Analyse von DNA- und Proteinsequenzen Kurs 1 Monika Oberer, Karl Gruber MOL.504 Modul-Übersicht Einführung, Datenbanken BLAST-Suche, Sequenzalignment Proteinstrukturen Virtuelles Klonieren Abschlusstest
Supporting Information
 Supporting Information Wiley-VCH 2013 69451 Weinheim, Germany Recoding the Genetic Code with Selenocysteine** Markus J. Brçcker, Joanne M. L. Ho, George M. Church, Dieter Sçll,* and Patrick ODonoghue*
Supporting Information Wiley-VCH 2013 69451 Weinheim, Germany Recoding the Genetic Code with Selenocysteine** Markus J. Brçcker, Joanne M. L. Ho, George M. Church, Dieter Sçll,* and Patrick ODonoghue*
MOL.504 Analyse von DNA- und Proteinsequenzen
 MOL.504 Analyse von DNA- und Proteinsequenzen Vorbereitung: Start des Serial Cloners ( unter Laufwerk K: im Ordner Win_SerialCloner2-5 Programm direkt im Ordner starten) K:\Win_SerialCloner2-5 (oder 2-6
MOL.504 Analyse von DNA- und Proteinsequenzen Vorbereitung: Start des Serial Cloners ( unter Laufwerk K: im Ordner Win_SerialCloner2-5 Programm direkt im Ordner starten) K:\Win_SerialCloner2-5 (oder 2-6
4. ERGEBNISSE. 4.2. Untersuchung des umgelagerten IgH Gens in Hodgkinzelllinien
 36 4. ERGEBNISSE 4.1. Immunglobulin Gentranskripte in Hodgkinzelllinien Mit Hilfe der RT PCR untersuchten wir die Expression umgelagerter Ig Gene in den Hodgkinzelllinien L1236, L428, L591 und KM-H2 sowie
36 4. ERGEBNISSE 4.1. Immunglobulin Gentranskripte in Hodgkinzelllinien Mit Hilfe der RT PCR untersuchten wir die Expression umgelagerter Ig Gene in den Hodgkinzelllinien L1236, L428, L591 und KM-H2 sowie
Evidence of strobilurine resistant isolates of A. solani and A. alternata in Germany
 Technische Universität München Evidence of strobilurine resistant isolates of A. solani and A. alternata in Germany H. Hausladen, B. Adolf and J. Leiminger outline introduction material and methods results
Technische Universität München Evidence of strobilurine resistant isolates of A. solani and A. alternata in Germany H. Hausladen, B. Adolf and J. Leiminger outline introduction material and methods results
Klinik und Poliklinik für Psychiatrie und Psychotherapie der Universität Würzburg Direktor: Professor Dr. med. H. Beckmann
 Klinik und Poliklinik für Psychiatrie und Psychotherapie der Universität Würzburg Direktor: Professor Dr. med. H. Beckmann Mutationsanalyse der Gene Connexin 36 (CX36) und Tyrosinkinase 3 (TYRO3) als Kandidatengene
Klinik und Poliklinik für Psychiatrie und Psychotherapie der Universität Würzburg Direktor: Professor Dr. med. H. Beckmann Mutationsanalyse der Gene Connexin 36 (CX36) und Tyrosinkinase 3 (TYRO3) als Kandidatengene
2. Material und Methoden
 Material und Methoden 13 2. Material und Methoden 2.1 Material 2.1.1 Gewebe 2.1.1.1 Gefriermaterial Für die Einzelzellanalyse wurden fünf Fälle des klassischen Hodgkin-Lymphoms (chl) ausgewählt. Dabei
Material und Methoden 13 2. Material und Methoden 2.1 Material 2.1.1 Gewebe 2.1.1.1 Gefriermaterial Für die Einzelzellanalyse wurden fünf Fälle des klassischen Hodgkin-Lymphoms (chl) ausgewählt. Dabei
Kaninchen: 1 cagattgaggagagcaccaccatcgtcacctcgcagttggcggaggtcagcgcagccgag 60
 Ergebnisse 6 3. Ergebnisse 3. Charakterisierung der zelllinienspezifischen cdn Die Entwicklung zweier Zelllinien, des epithelialen Trophoblasten und des pluripotenten Embryoblasten, ist der erste Differenzierungsschritt
Ergebnisse 6 3. Ergebnisse 3. Charakterisierung der zelllinienspezifischen cdn Die Entwicklung zweier Zelllinien, des epithelialen Trophoblasten und des pluripotenten Embryoblasten, ist der erste Differenzierungsschritt
Modul 8: Bioinformatik A. Von der DNA zum Protein Proteinsynthese in silicio
 Modul 8: Bioinformatik A. Von der DNA zum Protein Proteinsynthese in silicio Ein Wissenschaftler erhält nach einer Sequenzierung folgenden Ausschnitt aus einer DNA-Sequenz: 5 ctaccatcaa tccggtaggt tttccggctg
Modul 8: Bioinformatik A. Von der DNA zum Protein Proteinsynthese in silicio Ein Wissenschaftler erhält nach einer Sequenzierung folgenden Ausschnitt aus einer DNA-Sequenz: 5 ctaccatcaa tccggtaggt tttccggctg
DNS-Modell Best.-Nr. 2015801
 DNS-Modell Best.-Nr. 2015801 1. Produktvorstellung Ziel des Produktes Dieses Modell soll das DNS-Molekül visualisieren: es soll die Doppelspirale, Stickstoffbasen mit Wasserstoffbrückenbindung, Zucker-Phosphatskelette
DNS-Modell Best.-Nr. 2015801 1. Produktvorstellung Ziel des Produktes Dieses Modell soll das DNS-Molekül visualisieren: es soll die Doppelspirale, Stickstoffbasen mit Wasserstoffbrückenbindung, Zucker-Phosphatskelette
Die Rolle der microrna-29 während der Leberfibrogenese
 Die Rolle der microrna-29 während der Leberfibrogenese Inaugural-Dissertation zur Erlangung des Doktorgrades der Mathematisch-Naturwissenschaftlichen Fakultät der Universität zu Köln vorgelegt von Monika
Die Rolle der microrna-29 während der Leberfibrogenese Inaugural-Dissertation zur Erlangung des Doktorgrades der Mathematisch-Naturwissenschaftlichen Fakultät der Universität zu Köln vorgelegt von Monika
Versuch 2: Polymerasekettenreaktion. Betreuer: Knut Jahreis. Versuchsinhalt. Polymerasekettenreaktion, Gelelektrophorese auf Agarosegelen
 Versuch 2: Polymerasekettenreaktion Betreuer: Knut Jahreis Versuchsinhalt Polymerasekettenreaktion, Gelelektrophorese auf Agarosegelen Zwei verschiedene Plasmide werden als Matrizen für eine Polymerasekettenreaktion
Versuch 2: Polymerasekettenreaktion Betreuer: Knut Jahreis Versuchsinhalt Polymerasekettenreaktion, Gelelektrophorese auf Agarosegelen Zwei verschiedene Plasmide werden als Matrizen für eine Polymerasekettenreaktion
Charakterisierung der mitochondrialen RNA-Prozessierungs-Faktoren 4 und 6 in Arabidopsis thaliana
 Charakterisierung der mitochondrialen RNA-Prozessierungs-Faktoren 4 und 6 in Arabidopsis thaliana Dissertation zur Erlangung des Doktorgrades Dr. rer. nat. der Fakultät für Naturwissenschaften der Universität
Charakterisierung der mitochondrialen RNA-Prozessierungs-Faktoren 4 und 6 in Arabidopsis thaliana Dissertation zur Erlangung des Doktorgrades Dr. rer. nat. der Fakultät für Naturwissenschaften der Universität
TEPZZ 684965A_T EP 2 684 965 A1 (19) (11) EP 2 684 965 A1 (12) EUROPÄISCHE PATENTANMELDUNG. (43) Veröffentlichungstag: 15.01.2014 Patentblatt 2014/03
 (11) EP 2 684 965 A1 (12) EUROPÄISCHE PATENTANMELDUNG. (43) Veröffentlichungstag: 15.01.2014 Patentblatt 2014/03") (19) TEPZZ 68496A_T (11) EP 2 684 96 A1 (12) EUROPÄISCHE PATENTANMELDUNG (43) Veröffentlichungstag:.01.14 Patentblatt 14/03 (1) Int Cl.: C12Q 1/68 (06.01) (21) Anmeldenummer: 1318471.9 (22) Anmeldetag:
(19) TEPZZ 68496A_T (11) EP 2 684 96 A1 (12) EUROPÄISCHE PATENTANMELDUNG (43) Veröffentlichungstag:.01.14 Patentblatt 14/03 (1) Int Cl.: C12Q 1/68 (06.01) (21) Anmeldenummer: 1318471.9 (22) Anmeldetag:
B. Material und Methoden
 B. Material und Methoden I. Arbeitsmethoden I.1 Nabelschnurblut I.1.1 Sammlung und Lagerung von Nabelschnurblut (CB) Das in Rahmen dieser Arbeit verwendete Nabelschnurblut (engl. cord blood, CB) erhielt
B. Material und Methoden I. Arbeitsmethoden I.1 Nabelschnurblut I.1.1 Sammlung und Lagerung von Nabelschnurblut (CB) Das in Rahmen dieser Arbeit verwendete Nabelschnurblut (engl. cord blood, CB) erhielt
Etablierung einer. Homemade - PCR
 Etablierung einer Homemade - PCR Anja Schöpflin Institut für Pathologie Universitätsklinikum Freiburg Überblick: Anwendungsgebiete der PCR Anforderungen an Primer Auswahl geeigneter Primer / Primerdesign
Etablierung einer Homemade - PCR Anja Schöpflin Institut für Pathologie Universitätsklinikum Freiburg Überblick: Anwendungsgebiete der PCR Anforderungen an Primer Auswahl geeigneter Primer / Primerdesign
 Die vorliegende Arbeit entstand in der Zeit von April 2001 bis April 2004 an der Klinik und Poliklinik für Chirurgie des Klinikums der Universität Regensburg. Die externe Betreuung übernahm Herr PD Dr.
Die vorliegende Arbeit entstand in der Zeit von April 2001 bis April 2004 an der Klinik und Poliklinik für Chirurgie des Klinikums der Universität Regensburg. Die externe Betreuung übernahm Herr PD Dr.
Andrée, S. und F. Schwägele (2011) Mitteilungsblatt Fleischforschung Kulmbach 50, Nr. 193, 163-171
 Mitteilungsblatt Fleischforschung Kulmbach 50, Nr. 193, 163-171") , 163-171 Authentifizierung ausgewählter Geflügelspezies mittels Pentaplex-Real-Time-PCR Authentication of selected poultry species by means of pentaplex real-time PCR S. ANDRÉE und F. SCHWÄGELE Zusammenfassung
, 163-171 Authentifizierung ausgewählter Geflügelspezies mittels Pentaplex-Real-Time-PCR Authentication of selected poultry species by means of pentaplex real-time PCR S. ANDRÉE und F. SCHWÄGELE Zusammenfassung
Bedeutung der N-Glykosylierung der Chemokinrezeptoren CXCR4 und CCR5 für die Infektion mit dem Humanen Immundefizienzvirus Typ-1
 Bedeutung der N-Glykosylierung der Chemokinrezeptoren CXCR4 und CCR5 für die Infektion mit dem Humanen Immundefizienzvirus Typ-1 Dissertation zur Erlangung des Doktorgrades der Mathematischen-Naturwissenschaftlichen
Bedeutung der N-Glykosylierung der Chemokinrezeptoren CXCR4 und CCR5 für die Infektion mit dem Humanen Immundefizienzvirus Typ-1 Dissertation zur Erlangung des Doktorgrades der Mathematischen-Naturwissenschaftlichen
Aminosäuren - Proteine
 Aminosäuren - Proteine ÜBERBLICK D.Pflumm KSR / MSE Aminosäuren Überblick Allgemeine Formel für das Grundgerüst einer Aminosäure Carboxylgruppe: R-COOH O Aminogruppe: R-NH 2 einzelnes C-Atom (α-c-atom)
Aminosäuren - Proteine ÜBERBLICK D.Pflumm KSR / MSE Aminosäuren Überblick Allgemeine Formel für das Grundgerüst einer Aminosäure Carboxylgruppe: R-COOH O Aminogruppe: R-NH 2 einzelnes C-Atom (α-c-atom)
Peptide Proteine. 1. Aminosäuren. Alle optisch aktiven proteinogenen Aminosäuren gehören der L-Reihe an: 1.1 Struktur der Aminosäuren
 1. Aminosäuren Aminosäuren Peptide Proteine Vortragender: Dr. W. Helliger 1.1 Struktur 1.2 Säure-Basen-Eigenschaften 1.2.1 Neutral- und Zwitterion-Form 1.2.2 Molekülform in Abhängigkeit vom ph-wert 1.3
1. Aminosäuren Aminosäuren Peptide Proteine Vortragender: Dr. W. Helliger 1.1 Struktur 1.2 Säure-Basen-Eigenschaften 1.2.1 Neutral- und Zwitterion-Form 1.2.2 Molekülform in Abhängigkeit vom ph-wert 1.3
Analyse der in organello RNA-Prozessierung nach Elektroporation von Mitochondrien
 Analyse der in organello RNA-Prozessierung nach Elektroporation von Mitochondrien Dissertation zur Erlangung des Doktorgrades der Mathematisch-Naturwissenschaftlichen Fakultät der Christian-Albrechts-Universität
Analyse der in organello RNA-Prozessierung nach Elektroporation von Mitochondrien Dissertation zur Erlangung des Doktorgrades der Mathematisch-Naturwissenschaftlichen Fakultät der Christian-Albrechts-Universität
X - chromosomal dominanter hypophosphatämischer Rachitis
 Aus der Klinik für Kinder und Jugendmedizin der Universität zu Lübeck Direktor : Prof. Dr. E. Herting Molekulargenetische Untersuchungen bei Patienten mit X - chromosomal dominanter hypophosphatämischer
Aus der Klinik für Kinder und Jugendmedizin der Universität zu Lübeck Direktor : Prof. Dr. E. Herting Molekulargenetische Untersuchungen bei Patienten mit X - chromosomal dominanter hypophosphatämischer
2. Konstruktion und Charakterisierung einer stx 2 -Mutante im E. coli-stamm O157:H7 86-24
 IV. Ergebnisse 90 2. Konstruktion und Charakterisierung einer stx 2 -Mutante im E. coli-stamm O157:H7 86-24 Die Konstruktion einer isogenen Toxinmutante des Stx2-produzierenden EHEC- Stamm O157:H7 86-24
IV. Ergebnisse 90 2. Konstruktion und Charakterisierung einer stx 2 -Mutante im E. coli-stamm O157:H7 86-24 Die Konstruktion einer isogenen Toxinmutante des Stx2-produzierenden EHEC- Stamm O157:H7 86-24
Tanzania Wildlife Discussion Paper No. 25. Ludwig Siege, Rolf D. Baldus (Eds.)
") Tanzania Wildlife Discussion Paper No. 25 Ludwig Siege, Rolf D. Baldus (Eds.) Phylogenetische Differenzierung von Subpopulationen der Rappenantilope (Hippotragus Niger) in Ostafrika with an english foreword/summary
Tanzania Wildlife Discussion Paper No. 25 Ludwig Siege, Rolf D. Baldus (Eds.) Phylogenetische Differenzierung von Subpopulationen der Rappenantilope (Hippotragus Niger) in Ostafrika with an english foreword/summary
Molekulare Präsentationstechniken Phage Display
 Molekulare Präsentationstechniken Phage Display Gentechnik und Genomics WiSe 2007/2008 Kristian M. Müller Institut für Biologie III Albert-Ludwigs-Universität Freiburg Pharma-Biotech Umsatz in den USA
Molekulare Präsentationstechniken Phage Display Gentechnik und Genomics WiSe 2007/2008 Kristian M. Müller Institut für Biologie III Albert-Ludwigs-Universität Freiburg Pharma-Biotech Umsatz in den USA
Aus der Medizinischen Universitätsklinik - Abteilung Innere Medizin IV - der Albert-Ludwigs-Universität Freiburg im Breisgau
 I Aus der Medizinischen Universitätsklinik - Abteilung Innere Medizin IV - der Albert-Ludwigs-Universität Freiburg im Breisgau Das Phäochromozyttom beii Neurofiibromattose Typ 1 Klliiniik und Mollekullargenettiik
I Aus der Medizinischen Universitätsklinik - Abteilung Innere Medizin IV - der Albert-Ludwigs-Universität Freiburg im Breisgau Das Phäochromozyttom beii Neurofiibromattose Typ 1 Klliiniik und Mollekullargenettiik
MOLEKULARE DIAGNOSTIK,THERAPIE UND EPIDEMIOLOGIE VON ERREGERN VIRALER BERÜCKSICHTIGUNG DES LASSA-VIRUS HÄMORRHAGISCHER FIEBER UNTER BESONDERER
 MOLEKULARE DIAGNOSTIK,THERAPIE UND EPIDEMIOLOGIE VON ERREGERN VIRALER HÄMORRHAGISCHER FIEBER UNTER BESONDERER BERÜCKSICHTIGUNG DES LASSA-VIRUS Dissertation vorgelegt dem Fachbereich 2 (Biologie/Chemie)
MOLEKULARE DIAGNOSTIK,THERAPIE UND EPIDEMIOLOGIE VON ERREGERN VIRALER HÄMORRHAGISCHER FIEBER UNTER BESONDERER BERÜCKSICHTIGUNG DES LASSA-VIRUS Dissertation vorgelegt dem Fachbereich 2 (Biologie/Chemie)
Vom Gen zum Naturstoff!
 Pierre Stallforth // HKI Jena Vom Gen zum Naturstoff Sequenzierung: Wie erhalte ich genomische Informationen eines Organismus Genetische Knockouts, (CRISPR-Cas9) Veränderung: Bioinformatik: Datenbanken,
Pierre Stallforth // HKI Jena Vom Gen zum Naturstoff Sequenzierung: Wie erhalte ich genomische Informationen eines Organismus Genetische Knockouts, (CRISPR-Cas9) Veränderung: Bioinformatik: Datenbanken,
Bitte lesen Sie die folgenden Hinweise sorgfältig durch bevor Sie mit dem Lösen der Aufgaben beginnen.
 GYMNASIUM MUTTENZ MATURITÄTSPRÜFUNGEN 2008 FACH: Biologie TITEL EF: Verhalten Bitte lesen Sie die folgenden Hinweise sorgfältig durch bevor Sie mit dem Lösen der Aufgaben beginnen. Die Prüfung umfasst
GYMNASIUM MUTTENZ MATURITÄTSPRÜFUNGEN 2008 FACH: Biologie TITEL EF: Verhalten Bitte lesen Sie die folgenden Hinweise sorgfältig durch bevor Sie mit dem Lösen der Aufgaben beginnen. Die Prüfung umfasst
Klassifizierung: noch nicht abgeschlossen, Verwandtschaftsverhältnis zur Familie: Desulvovibrionaceae - Vertreter Desulvovibrio desulfuricans
 AVID X / 1998 Lawsonia intracellularis Seite -1- Klassifizierung: noch nicht abgeschlossen, Verwandtschaftsverhältnis zur Familie: Desulvovibrionaceae - Vertreter Desulvovibrio desulfuricans 1. ALLGEMEINES
AVID X / 1998 Lawsonia intracellularis Seite -1- Klassifizierung: noch nicht abgeschlossen, Verwandtschaftsverhältnis zur Familie: Desulvovibrionaceae - Vertreter Desulvovibrio desulfuricans 1. ALLGEMEINES
Entwicklung von Replikonsystemen mit subgenomischen Abschnitten von HCV-RNA Genotyp 3a Isolaten klinisch charakterisierter Patienten
 Entwicklung von Replikonsystemen mit subgenomischen Abschnitten von HV-RNA enotyp 3a Isolaten klinisch charakterisierter Patienten INAUURAL-DISSERTATION zur Erlangung des rades eines Dr. med. vet. beim
Entwicklung von Replikonsystemen mit subgenomischen Abschnitten von HV-RNA enotyp 3a Isolaten klinisch charakterisierter Patienten INAUURAL-DISSERTATION zur Erlangung des rades eines Dr. med. vet. beim
ScieCon. Messekatalog. ScieCon München 2014. ScieCon Dein Jobpromotor! Viele Firmen Ein Weg Dein Job. Biowissenschaften I Chemie I Pharmazie I Medizin
 ScieCon Dein Jobpromotor! Biowissenschaften I Chemie I Pharmazie I Medizin Messekatalog ScieCon München 2014 Viele Firmen Ein Weg Dein Job ScieCon München 2014 Klinikum Großhadern 29. Januar 2014, 10-17
ScieCon Dein Jobpromotor! Biowissenschaften I Chemie I Pharmazie I Medizin Messekatalog ScieCon München 2014 Viele Firmen Ein Weg Dein Job ScieCon München 2014 Klinikum Großhadern 29. Januar 2014, 10-17
LIETUVOS STANDARTAS LST EN ISO 21570/AC
 LIETUVOS STANDARTAS LST EN ISO 21570/AC PATAISA AC 2007 m. vasaris ICS 67.050 Maisto produktai. Analiz s metodai aptikti genetiškai modifikuotus organizmus ir jų produktus. Kiekybiniai nukleorūgščių nustatymu
LIETUVOS STANDARTAS LST EN ISO 21570/AC PATAISA AC 2007 m. vasaris ICS 67.050 Maisto produktai. Analiz s metodai aptikti genetiškai modifikuotus organizmus ir jų produktus. Kiekybiniai nukleorūgščių nustatymu
4.1. Herstellung des CH2/CH3-trunkierten dimerisierten anti-cd30-igg1-tnf- Fusionsproteins
 ERGEBNISSE 29 4. Ergebnisse 4.1. Herstellung des CH2/CH3-trunkierten dimerisierten anti-cd3-igg1-tnf- Fusionsproteins Im vorliegenden Immunzytokin wurden die Domänen CH2/CH3 des humanen Fc-Fragmentes durch
ERGEBNISSE 29 4. Ergebnisse 4.1. Herstellung des CH2/CH3-trunkierten dimerisierten anti-cd3-igg1-tnf- Fusionsproteins Im vorliegenden Immunzytokin wurden die Domänen CH2/CH3 des humanen Fc-Fragmentes durch
3 Material und Methoden
 17 3 Material und Methoden 3.1 Zellkultur H9c2-Kardiomyoblasten (ATCC, Rockville, USA) wurden in Dulbeccos modifiziertem Eagle Medium (Bio-Whittacker, Verviers, Belgien; mit 4,5 g/l D- Glucose, 3,7 g/l
17 3 Material und Methoden 3.1 Zellkultur H9c2-Kardiomyoblasten (ATCC, Rockville, USA) wurden in Dulbeccos modifiziertem Eagle Medium (Bio-Whittacker, Verviers, Belgien; mit 4,5 g/l D- Glucose, 3,7 g/l
2 Material und Methoden
 2 Material und Methoden 21 Material 211 Chemikalien und Enzyme Die im Rahmen dieser Arbeit eingesetzten Chemikalien waren von analytischem Reinheitsgrad und wurden von den Firmen Biomol (Hamburg), Fluka
2 Material und Methoden 21 Material 211 Chemikalien und Enzyme Die im Rahmen dieser Arbeit eingesetzten Chemikalien waren von analytischem Reinheitsgrad und wurden von den Firmen Biomol (Hamburg), Fluka
Charakterisierung funktioneller Domänen des Vpr-Proteins von SIV mac 239
 Charakterisierung funktioneller Domänen des Vpr-Proteins von SIV mac 239 Diplomarbeit aus dem Institut für Klinische und Molekulare Virologie der Medizinischen Fakultät der Friedrich-Alexander-Universität
Charakterisierung funktioneller Domänen des Vpr-Proteins von SIV mac 239 Diplomarbeit aus dem Institut für Klinische und Molekulare Virologie der Medizinischen Fakultät der Friedrich-Alexander-Universität
Genisolierung in 2 Stunden : Die Polymerase-Ketten-Reaktion (PCR)
") PCR Genisolierung in 2 Stunden : Die Polymerase-Ketten-Reaktion (PCR) von Kary B. Mullis entwickelt (1985) eigentlich im Mai 1983 bei einer nächtlichen Autofahrt erstes erfolgreiches Experiment am 16.12.1983
PCR Genisolierung in 2 Stunden : Die Polymerase-Ketten-Reaktion (PCR) von Kary B. Mullis entwickelt (1985) eigentlich im Mai 1983 bei einer nächtlichen Autofahrt erstes erfolgreiches Experiment am 16.12.1983
3 Methoden. Methoden 24. 3.1 Zellkultur. Kultivieren von Zellen
 Methoden 24 3 Methoden 3.1 Zellkultur Kultivieren von Zellen Die Nierenkarzinomzelllinien A498, SN12 und ACHN (Referenzen für die verwendeten Zelllinien siehe unter 2.2) wurden nach Anleitung der American
Methoden 24 3 Methoden 3.1 Zellkultur Kultivieren von Zellen Die Nierenkarzinomzelllinien A498, SN12 und ACHN (Referenzen für die verwendeten Zelllinien siehe unter 2.2) wurden nach Anleitung der American
Messekatalog. ScieCon NRW 2014. ScieCon. Your Life Sciences Career. Let s Start it! Viele Firmen Ein Weg Dein Job.
 Your Life Sciences Career. Let s Start it! Biowissenschaften I Chemie I Pharmazie I Medizin Messekatalog ScieCon NRW 2014 Viele Firmen Ein Weg Dein Job ScieCon NRW 2014 Veranstaltungszentrum Ruhr-Universität
Your Life Sciences Career. Let s Start it! Biowissenschaften I Chemie I Pharmazie I Medizin Messekatalog ScieCon NRW 2014 Viele Firmen Ein Weg Dein Job ScieCon NRW 2014 Veranstaltungszentrum Ruhr-Universität
Modulation der Masernvirusinfektion durch RNA-Interferenz mittels mirna Expressionskassetten
 Modulation der Masernvirusinfektion durch RNA-Interferenz mittels mirna Expressionskassetten Dissertation zur Erlangung des akademischen Grades Dr. rer. med. an der Medizinischen Fakultät der Universität
Modulation der Masernvirusinfektion durch RNA-Interferenz mittels mirna Expressionskassetten Dissertation zur Erlangung des akademischen Grades Dr. rer. med. an der Medizinischen Fakultät der Universität
Die RT-PCR Untersuchungen wurden anhand 27 Struma nodosa- (siehe Tab. 7) und 15 Adenom- (siehe Tab. 11) Präparaten durchgeführt.
 und 15 Adenom- (siehe Tab. 11) Präparaten durchgeführt.") - 22-4 Ergebnisse 4.1 RT-PCR Die RT-PCR Untersuchungen wurden anhand 27 Struma nodosa- (siehe Tab. 7) und 15 Adenom- (siehe Tab. 11) Präparaten durchgeführt. 4.1.1 Struma nodosa Histologie Fas Fas CD 97
- 22-4 Ergebnisse 4.1 RT-PCR Die RT-PCR Untersuchungen wurden anhand 27 Struma nodosa- (siehe Tab. 7) und 15 Adenom- (siehe Tab. 11) Präparaten durchgeführt. 4.1.1 Struma nodosa Histologie Fas Fas CD 97
Identifizierung eines am Steroid- und Tetrapyrroltransport beteiligten Proteins in Pflanzen
 Identifizierung eines am Steroid- und Tetrapyrroltransport beteiligten Proteins in Pflanzen Dissertation zur Erlangung des akademischen Grades doctor rerum naturalium (Dr. rer. nat.) vorgelegt der Mathematisch-Naturwissenschaftlich-Technischen
Identifizierung eines am Steroid- und Tetrapyrroltransport beteiligten Proteins in Pflanzen Dissertation zur Erlangung des akademischen Grades doctor rerum naturalium (Dr. rer. nat.) vorgelegt der Mathematisch-Naturwissenschaftlich-Technischen
Zuschriften. Oligofunktionale Konjugate aus DNA und Gold- Nanopartikeln** Christof M. Niemeyer,* Bülent Ceyhan und Pompi Hazarika
 Nanotechnologie Oligofunktionale Konjugate aus DNA und Gold- Nanopartikeln** Christof M. Niemeyer,* Bülent Ceyhan und Pompi Hazarika Professor Manfred T. Reetz zum 60. Geburtstag gewidmet Hybrid-Konjugate
Nanotechnologie Oligofunktionale Konjugate aus DNA und Gold- Nanopartikeln** Christof M. Niemeyer,* Bülent Ceyhan und Pompi Hazarika Professor Manfred T. Reetz zum 60. Geburtstag gewidmet Hybrid-Konjugate
Regulation des VEGF-Systems in endometrialen Stromazellen: Untersuchungen zur mrna-expression der Rezeptoren VEGFR1 und sflt-1
 Aus der Frauenklinik der Heinrich-Heine-Universität Düsseldorf Direktor: Universitätsprofessor Dr. med. W. Janni Regulation des VEGF-Systems in endometrialen Stromazellen: Untersuchungen zur mrna-expression
Aus der Frauenklinik der Heinrich-Heine-Universität Düsseldorf Direktor: Universitätsprofessor Dr. med. W. Janni Regulation des VEGF-Systems in endometrialen Stromazellen: Untersuchungen zur mrna-expression
Untersuchungen zur Struktur und Funktion der Glutathionsynthetase bei der Spalthefe Schizosaccharomyces pombe
 Untersuchungen zur Struktur und Funktion der Glutathionsynthetase bei der Spalthefe Schizosaccharomyces pombe Von der Fakultät für Mathematik, Informatik und Naturwissenschaften der Rheinisch-Westfälischen
Untersuchungen zur Struktur und Funktion der Glutathionsynthetase bei der Spalthefe Schizosaccharomyces pombe Von der Fakultät für Mathematik, Informatik und Naturwissenschaften der Rheinisch-Westfälischen
Ergebnisse. I. Identifizierung intrazellulärer Interaktionspartner. Ergebnisse I. Identifizierung intrazellulärer Interaktionspartner
 Ergebnisse I. Identifizierung intrazellulärer Interaktionspartner Es sollten mithilfe des Yeast-Two-Hybrid-Systems Proteine identifiziert werden, die mit der zytoplasmatischen Domäne von CEACAM1-4L aus
Ergebnisse I. Identifizierung intrazellulärer Interaktionspartner Es sollten mithilfe des Yeast-Two-Hybrid-Systems Proteine identifiziert werden, die mit der zytoplasmatischen Domäne von CEACAM1-4L aus
Aliphatische Aminosäuren. Aromatische Aminosäuren
 Prof. Dr..-U. eißig rganische Chemie I 17.1 Aliphatische Aminosäuren systematischer ame (Trivialname) truktur Vorkommen, Bedeutung 2-Aminoethansäure (α-aminoessigsäure, Glycin) 3 C 2 C 2 α Proteine, Peptide
Prof. Dr..-U. eißig rganische Chemie I 17.1 Aliphatische Aminosäuren systematischer ame (Trivialname) truktur Vorkommen, Bedeutung 2-Aminoethansäure (α-aminoessigsäure, Glycin) 3 C 2 C 2 α Proteine, Peptide
Molekulare Ansätze zur Reduktion des allergenen Potenzials von Tomatenfrüchten
 Molekulare Ansätze zur Reduktion des allergenen Potenzials von Tomatenfrüchten Der Naturwissenschaftlichen Fakultät der Friedrich-Alexander-Universität Erlangen-Nürnberg zur Erlangung des Doktorgrades
Molekulare Ansätze zur Reduktion des allergenen Potenzials von Tomatenfrüchten Der Naturwissenschaftlichen Fakultät der Friedrich-Alexander-Universität Erlangen-Nürnberg zur Erlangung des Doktorgrades
β2-microglobulin deficient mice lack CD4-8+cytolytic T cells
 β2-microglobulin deficient mice lack CD4-8+cytolytic T cells Mäuse mit einem Knock-out bezüglich ß-Microglobulin sind nicht in der Lage CD4-8+ cytotoxische T-Zellen zu bilden Nature,Vol 344, 19. April
β2-microglobulin deficient mice lack CD4-8+cytolytic T cells Mäuse mit einem Knock-out bezüglich ß-Microglobulin sind nicht in der Lage CD4-8+ cytotoxische T-Zellen zu bilden Nature,Vol 344, 19. April
Fortbildungskurs Klinische Diabetologie der DDG 8. Oktober2012 Insulin: Biosynthese und Sekretion
 Fortbildungskurs Klinische Diabetologie der DDG 8. Oktober2012 Insulin: Biosynthese und ekretion Thomas Meissner Universitäts Kinderklinik Düsseldorf DDG Kurs 2012 Gliederung Insulin Insulinbiosynthese
Fortbildungskurs Klinische Diabetologie der DDG 8. Oktober2012 Insulin: Biosynthese und ekretion Thomas Meissner Universitäts Kinderklinik Düsseldorf DDG Kurs 2012 Gliederung Insulin Insulinbiosynthese
Aus dem Lehrstuhl für Innere Medizin I Prof. Dr. med. Jürgen Schölmerich Der Medizinischen Fakultät Der Universität Regensburg
 Aus dem Lehrstuhl für Innere Medizin I Prof. Dr. med. Jürgen Schölmerich Der Medizinischen Fakultät Der Universität Regensburg Untersuchung der Expression und Regulation von Komponenten des Inflammasoms
Aus dem Lehrstuhl für Innere Medizin I Prof. Dr. med. Jürgen Schölmerich Der Medizinischen Fakultät Der Universität Regensburg Untersuchung der Expression und Regulation von Komponenten des Inflammasoms
Das Sap47-Gen aus Drosophila melanogaster:
 Das Sap47-Gen aus Drosophila melanogaster: Gezielte Mutagenisierung und Suche nach Interaktionspartnern Dissertation zur Erlangung des naturwissenschaftlichen Doktorgrades der Bayerischen Julius-Maximilians-Universität
Das Sap47-Gen aus Drosophila melanogaster: Gezielte Mutagenisierung und Suche nach Interaktionspartnern Dissertation zur Erlangung des naturwissenschaftlichen Doktorgrades der Bayerischen Julius-Maximilians-Universität
Etablierung eines Hefe-Zwei-Hybrid-Systems zur Suche nach zytoplasmatischen Interaktionspartnern der α-protocadherine
 Aus dem Institut für Laboratoriumsmedizin und Pathobiochemie der Medizinischen Fakultät Charité Universitätsmedizin Berlin DISSERTATION Etablierung eines Hefe-Zwei-Hybrid-Systems zur Suche nach zytoplasmatischen
Aus dem Institut für Laboratoriumsmedizin und Pathobiochemie der Medizinischen Fakultät Charité Universitätsmedizin Berlin DISSERTATION Etablierung eines Hefe-Zwei-Hybrid-Systems zur Suche nach zytoplasmatischen
Europäisches Patentamt European Patent Office Veröffentlichungsnummer: 0 276 723 Office europeen des brevets EUROPAISCHE PATENTANMELDUNG
 J Europäisches Patentamt European Patent Office Veröffentlichungsnummer: 0 276 723 Office europeen des brevets A2 EUROPAISCHE PATENTANMELDUNG Anmeldenummer: 88100647.2 Int. Cl.4: C07H 21/04 ^ Anmeldetag:
J Europäisches Patentamt European Patent Office Veröffentlichungsnummer: 0 276 723 Office europeen des brevets A2 EUROPAISCHE PATENTANMELDUNG Anmeldenummer: 88100647.2 Int. Cl.4: C07H 21/04 ^ Anmeldetag:
1. Stammbaum einer Familie, in der Mukoviszidose aufgetreten ist.
 Die Prüfungsarbeit besteht aus drei zu bearbeitenden Teilen Aufgabe I Aufgabe II A oder II B Aufgabe III A oder III B I Aufgabe I: Humangenetik / klassische Genetik / Molekulargenetik Mukoviszidose Mukoviszidose,
Die Prüfungsarbeit besteht aus drei zu bearbeitenden Teilen Aufgabe I Aufgabe II A oder II B Aufgabe III A oder III B I Aufgabe I: Humangenetik / klassische Genetik / Molekulargenetik Mukoviszidose Mukoviszidose,
Kursinhalte der Weiterbildung Molekulare Biotechnologie (MNr.: 237 / 0411 / 2010)
") Kursinhalte der Weiterbildung Molekulare Biotechnologie (MNr.: 237 / 0411 / 2010) Schwerpunkte im Bereich BIOANALYTIK Polyacrylamidelektrophorese von Nukleinsäuren im Vertikalsystem Agarosegelelektrophorese
Kursinhalte der Weiterbildung Molekulare Biotechnologie (MNr.: 237 / 0411 / 2010) Schwerpunkte im Bereich BIOANALYTIK Polyacrylamidelektrophorese von Nukleinsäuren im Vertikalsystem Agarosegelelektrophorese
Europäisches Patentamt 19 J» European Patent Office Office europeen des brevets Veröffentlichungsnummer : 0 531 300 B1 12 EUROPAISCHE PATENTSCHRIFT
 Eräishes atentamt 19 J» Erean atent Offie Offie ereen des brevs Veröffentlihngsnmmer : 0 51 00 B1 12 EUROASCHE ATENTSCHRFT @ Veröffentlihngstag der atentshrift : 08.06.94 atentblatt 94/2 @ Anmeldenmmer
Eräishes atentamt 19 J» Erean atent Offie Offie ereen des brevs Veröffentlihngsnmmer : 0 51 00 B1 12 EUROASCHE ATENTSCHRFT @ Veröffentlihngstag der atentshrift : 08.06.94 atentblatt 94/2 @ Anmeldenmmer
Online-Katalog zur Ausstellung LEBENS LANDSCHAFTEN <> GENESCAPES vom 11.09. bis 08.10.2011 KARSTEN K. PANZER PERZAN. Kunstverein Wesseling e.v.
 Kunstverein Wesseling e.v. Städtische Galerie & Scheunengalerie Schwingeler Hof Schwingelerweg 44, 50389 Wesseling Online-Katalog zur Ausstellung LEBENS LANDSCHAFTEN GENESCAPES vom 11.09. bis 08.10.2011
Kunstverein Wesseling e.v. Städtische Galerie & Scheunengalerie Schwingeler Hof Schwingelerweg 44, 50389 Wesseling Online-Katalog zur Ausstellung LEBENS LANDSCHAFTEN GENESCAPES vom 11.09. bis 08.10.2011
Studienkolleg der Technischen Universität Berlin. Biologie-Prüfung. für BewerberInnen mit Beruflicher Qualifikation nach 11 BerlHG
 Studienkolleg der Technischen Universität Berlin Biologie-Prüfung für BewerberInnen mit Beruflicher Qualifikation nach 11 BerlHG Teil 1 Markieren Sie bitte die richtige Antwort. (pro richtiger Antwort
Studienkolleg der Technischen Universität Berlin Biologie-Prüfung für BewerberInnen mit Beruflicher Qualifikation nach 11 BerlHG Teil 1 Markieren Sie bitte die richtige Antwort. (pro richtiger Antwort
Praktikum Biochemie Biotechnologie (Molekularbiologie & Biochemie) Bettina Siebers
 Bettina Siebers") Praktikum Biochemie Biotechnologie (Molekularbiologie & Biochemie) Bettina Siebers FIGURE 20.1 Biology 6/e Biotechnologie Protein Expression Genomische DNA PCR Vektormolekül (Plasmid) Escherichia coli
Praktikum Biochemie Biotechnologie (Molekularbiologie & Biochemie) Bettina Siebers FIGURE 20.1 Biology 6/e Biotechnologie Protein Expression Genomische DNA PCR Vektormolekül (Plasmid) Escherichia coli
Chemie für Biologen. Vorlesung im. WS 2004/05 V2, Mi 10-12, S04 T01 A02. Paul Rademacher Institut für Organische Chemie der Universität Duisburg-Essen
 Chemie für Biologen Vorlesung im WS 004/05 V, Mi 0-, S04 T0 A0 Paul Rademacher Institut für rganische Chemie der Universität Duisburg-Essen (Teil 3: 9.0.005) MILESS: Chemie für Biologen 36 D-Aldosen C
Chemie für Biologen Vorlesung im WS 004/05 V, Mi 0-, S04 T0 A0 Paul Rademacher Institut für rganische Chemie der Universität Duisburg-Essen (Teil 3: 9.0.005) MILESS: Chemie für Biologen 36 D-Aldosen C
Aus dem Institut für Physiologische Chemie I der Universität Würzburg Vorstand: Professor Dr. rer. nat. Manfred Schartl
 Aus dem Institut für Physiologische Chemie I der Universität Würzburg Vorstand: Professor Dr. rer. nat. Manfred Schartl Biochemische Charakterisierung der basischen Helix-Loop-Helix Transkriptionsfaktoren
Aus dem Institut für Physiologische Chemie I der Universität Würzburg Vorstand: Professor Dr. rer. nat. Manfred Schartl Biochemische Charakterisierung der basischen Helix-Loop-Helix Transkriptionsfaktoren
8. Translation. Konzepte: Translation benötigt trnas und Ribosomen. Genetischer Code. Initiation - Elongation - Termination
 8. Translation Konzepte: Translation benötigt trnas und Ribosomen Genetischer Code Initiation - Elongation - Termination 1. Welche Typen von RNAs gibt es und welches sind ihre Funktionen? mouse human bacteria
8. Translation Konzepte: Translation benötigt trnas und Ribosomen Genetischer Code Initiation - Elongation - Termination 1. Welche Typen von RNAs gibt es und welches sind ihre Funktionen? mouse human bacteria
4.2 Kokulturen Epithelzellen und Makrophagen
 Ergebnisse 4.2 Kokulturen Epithelzellen und Makrophagen Nach der eingehenden Untersuchung der einzelnen Zelllinien wurden die Versuche auf Kokulturen aus den A549-Epithelzellen und den Makrophagenzelllinien
Ergebnisse 4.2 Kokulturen Epithelzellen und Makrophagen Nach der eingehenden Untersuchung der einzelnen Zelllinien wurden die Versuche auf Kokulturen aus den A549-Epithelzellen und den Makrophagenzelllinien
PATENTANMELDUNG. int.ci.6:c12n 9/86, C12N 15/55, C12N 1/21, C12N 1/19, C12P 21/02, C12P 13/04
 Europäisches Patentamt European Patent Office Office europeen des brevets Veröffentlichungsnummer: 0 643 1 33 A2 EUROPAISCHE PATENTANMELDUNG Anmeldenummer: 94113110.4 @ Anmeldetag: 23.08.94 int.ci.6:c12n
Europäisches Patentamt European Patent Office Office europeen des brevets Veröffentlichungsnummer: 0 643 1 33 A2 EUROPAISCHE PATENTANMELDUNG Anmeldenummer: 94113110.4 @ Anmeldetag: 23.08.94 int.ci.6:c12n
Regulation kernlokalisierter mitochondrialer Gene bei der Muskeldifferenzierung
 Regulation kernlokalisierter mitochondrialer Gene bei der Muskeldifferenzierung I n a u g u r a l - D i s s e r t a t i o n zur Erlangung des Doktorgrades der Mathematisch-Naturwissenschaftlichen Fakultät
Regulation kernlokalisierter mitochondrialer Gene bei der Muskeldifferenzierung I n a u g u r a l - D i s s e r t a t i o n zur Erlangung des Doktorgrades der Mathematisch-Naturwissenschaftlichen Fakultät
uropaisches Patentamt. iuropean Patent Office )ffice europeen des brevets y Veröffentlichungsnummer: \2 PATcNTANIvibLUUNva EUROPÄISCHE
ffice europeen des brevets y Veröffentlichungsnummer: \2 PATcNTANIvibLUUNva EUROPÄISCHE") uropaisches Patentamt. J ) iuropean Patent Office )ffice europeen des brevets y Veröffentlichungsnummer: \2 EUROPÄISCHE PATcNTANIvibLUUNva i) Anmeldenummer: 89122438.8 ) Anmeldetag: 05.12.89 n) int. CIA
uropaisches Patentamt. J ) iuropean Patent Office )ffice europeen des brevets y Veröffentlichungsnummer: \2 EUROPÄISCHE PATcNTANIvibLUUNva i) Anmeldenummer: 89122438.8 ) Anmeldetag: 05.12.89 n) int. CIA
Rainer Merkl und Stephan Waack Bioinformatik Interaktiv
 Rainer Merkl und Stephan Waack Bioinformatik Interaktiv Beachten Sie bitte auch weitere interessante Titel zu diesem Thema Helms, V. Principles of Computational Cell Biology From Protein Complexes to Cellular
Rainer Merkl und Stephan Waack Bioinformatik Interaktiv Beachten Sie bitte auch weitere interessante Titel zu diesem Thema Helms, V. Principles of Computational Cell Biology From Protein Complexes to Cellular
2 Material. 2.1 Geräte. 2.1.1 Elektrophorese- und Blotapparaturen
 2 Material 2.1 Geräte 2.1.1 Elektrophorese- und Blotapparaturen Elekrophoreseapparatur Dual Vertical Mini Gel (C.B.S., Del Mar, CA, USA) Elekrophoreseapparatur Trans-Blot SD Semi-Dry Transfer Cell (Bio-Rad,
2 Material 2.1 Geräte 2.1.1 Elektrophorese- und Blotapparaturen Elekrophoreseapparatur Dual Vertical Mini Gel (C.B.S., Del Mar, CA, USA) Elekrophoreseapparatur Trans-Blot SD Semi-Dry Transfer Cell (Bio-Rad,
Anlage zur Akkreditierungsurkunde D PL 13372 01 00
 Deutsche Akkreditierungsstelle GmbH Anlage zur Akkreditierungsurkunde D PL 13372 01 00 nach DIN EN ISO/IEC 17025:2005 Gültigkeitsdauer: 22.05.2014 bis 25.03.2017 Ausstellungsdatum: 22.05.2014 Urkundeninhaber:
Deutsche Akkreditierungsstelle GmbH Anlage zur Akkreditierungsurkunde D PL 13372 01 00 nach DIN EN ISO/IEC 17025:2005 Gültigkeitsdauer: 22.05.2014 bis 25.03.2017 Ausstellungsdatum: 22.05.2014 Urkundeninhaber:
PCR und Fluoreszenz - DNA - Fragment - Analyse zum Nachweis einer monoklonalen B-Zell-Population zur Diagnostik der kutanen B-Zell Lymphome (CBCL)
") Aus der Klinik für Dermatologie der Medizinischen Fakultät Charité der Humboldt-Universität zu Berlin DISSERTATION PCR und Fluoreszenz - DNA - Fragment - Analyse zum Nachweis einer monoklonalen B-Zell-Population
Aus der Klinik für Dermatologie der Medizinischen Fakultät Charité der Humboldt-Universität zu Berlin DISSERTATION PCR und Fluoreszenz - DNA - Fragment - Analyse zum Nachweis einer monoklonalen B-Zell-Population
Aus dem Institut für Humangenetik der Medizinischen Fakultät der Charite- Universitätsmedizin Berlin. Dissertation
 Aus dem Institut für Humangenetik der Medizinischen Fakultät der Charite- Universitätsmedizin Berlin Dissertation Mutationsanalyse im NBS1-Gen bei Patientinnen mit BRCA1/2 negativem familiären und BRCA1/2
Aus dem Institut für Humangenetik der Medizinischen Fakultät der Charite- Universitätsmedizin Berlin Dissertation Mutationsanalyse im NBS1-Gen bei Patientinnen mit BRCA1/2 negativem familiären und BRCA1/2