Bayerisches Staatsministerium tür Landwirtschaft und Forsten
|
|
|
- Gerburg Schwarz
- vor 7 Jahren
- Abrufe
Transkript


1
2 Alle Rechte vorbehalten. Nachdruck, auch auszugsweise, sowie fotomechanische und elektronische Wiedergabe nur mit Genehmigung des Herausgebers. Herausgeber: und Bezugsadresse: Nationalparkverwaltung Bayerischer Wald Freyunger Straße Grafenau Telefon / Telefax / poststelle@fonpv-bay.bayern.de Internet: Schriftleitung: Verantwortlich: Gesta Itu ng: Kartengestaltung : Litho u. Druck: Marco Heurich Der Leiter der Nationalparkverwaltung Bayerischer Wald, Karl Friedrich Sinner Annemarie Schmeller in Zusammenarbeit mit Atelier Et friends, Grafenau Marco Heurich, Arthur Reinelt, Michael Wotschikowsky Passavia Druckservice GmbH, Passau August 2001 ISBN Bayerisches Staatsministerium tür Landwirtschaft und Forsten
3 Waldentwicklung im Bergwald nach Windwurf und Borkenkäferbefall Nationalpark Bayerischer Wald
4 Vorwort Mit diesem Band wird die wissenschaftliche Schriften reihe des Nationalparks Bayerischer Wald nach mehrjähriger Pause wieder aufgenommen. Das Erscheinungsbild der Schriftenreihe und das Layout mussten komplett neu gestaltet werden, um das selbstgesteckte Ziel einer hochwertigen Fachpublikation, die sich auch an interessierte Nationalparkbesucher wendet, zu erreichen. Diese Forschungsfeldersind allerdings bei weitem noch nicht abgedeckt. Hier sind vermehrt interdisziplinäre Zusammenarbeit und Kooperation notwendig. Mit der wissenschaftlichen Schriftenreihe des Nationalparks Bayerischer Wald existiert eine Plattform, in welcher die Ergebnisse dieser Forschungsarbeiten publiziert und damit einem größeren Leserkreis zugänglich werden. In dieser ersten Neuausgabe wird das in den rückliegenden Jahren wichtigste Thema vorgestellt, die durch die Entwicklung des Borkenkäfers ausgelöste umfassende Veränderung des Waldes im Nationalpark. Diese säkulare Entwicklung der Nationalparkwälder hat eine gründliche Überarbeitung des Forschungskonzeptes des Nationalparks Bayerischer Wald notwendig gemacht. Viele neue Kooperationen mit den unterschiedlichsten Forschungseinrichtungen sind in den letzten Jahren entstanden, die gezielte Fragestellungen im Rahmen des Leitthemas der Forschung im Nationalpark "Steuergrößen natürlicher Waidentwicklung" bearbeiten. Ich danke allen Mitarbeiterinnen und Mitarbeitern des Nationalparks, die sich der nicht leichten Aufgabe gestellt haben, die Schriftenreihe in neuem Gewand und vor allem mit entsprechendem Inhalt wieder erscheinen zu lassen. Wir erhoffen uns durch diese Publikation eine Intensivierung der wissenschaftlichen Diskussion und Kooperation in dem so ungeheuer spannenden und facettenreichen Projekt Nationalpark Bayerischer Wald. Grafenau, Juli 2001 ~i~~~ Leiter der Nationalparkverwaltung Bayerischer Wald 4
5 Zur Einführung Die Bergfichtenwälder im Nationalpark durchlaufen eine dramatische Entwicklung, deren Dimension und Rasanz zwar von jedermann beobachtet werden können, die in ihrem Ursachen-Wirkungsgefüge aber nur bruchstückhaft verstanden wird. Speziell zur komplexen Verkettung von Sturmereignissen, Mehrung des Totholzanteils und Aufschwingen einer Borkenkäfer-Gradation auf der einen Seite und von schwindendem Widerstand gegen schädigende Umwelteinflüsse bei den Fichtenbeständen in den Hochlagen infolge von Überalterung - auch deren möglicher Benachteiligung durch ehemalige Wald pflege - auf der anderen Seite, bei gleichzeitigem Anstieg aerogener Schadstoffbelastung im Boden bzw. Wurzelraum sowie merklicher Verschiebung des Witterungsverlaufs (zu schneeärmeren Wintern und wärmeren Sommern) als drittem Faktorenkomplex, kann auf keinerlei Erfahrungswerte zurückgegriffen werden, weil derartige Entwicklungen in Mitteleuropa bisher einfach nicht zugelassen werden konnten. Hier eröffnet das Nationalpark-Konzept - gemäß dem Motto "Natur Natur sein lassen" - ganz neue Aspekte : Durch die Sicherung natürlicher, d. h. vom Menschen nicht willentlich gelenkter Abläufe in nutzungsfreier Waldlandschaft ermöglicht der Nationalpark Bayerischer Wald ein einmaliges Natur-Experiment, dessen wissenschaftliche Beobachtung und Dokumentation als außerordentliche Herausforderung für die Nationalpark-Forschung erkannt wurde. Der vorliegende Sammelband geht in drei Fachbeiträgen der zentralen Fragestellung nach : Wie verläuft der Umgestaltungs-Prozess eines bisher vom Menschen gestalteten bzw. bewirtschafteten Fichtenwaldes zum "Urwald von morgen" - unter dem Einfluss abiotischer (z. B. Sturmwurt) wie biotischer Störereignisse (z. B. Massenvermehrung baumschädigender Insekten, auch Wildverbiss), in Abhängigkeit zu Witterung, Samenbank (= Samen reservoir im Waldboden) und den Lebensstrategien der betroffenen Pflanzenarten (z. B. Produktion und Ausbreitung von Samen, Keimungschancen, zwischenartliche Konkurrenz und Lebenserwartung). In der Darstellung ihrer Erhebungen zur Entwicklung von Waldbeständen bzw. WaIdvegetation haben die Autoren unterschiedliche Betrachtungsebenen gewählt, die, in Abhängigkeit zur jeweiligen Korngröße der Datenaufnahme, deutlich verschiedene Interpretationsgenauigkeiten zulassen : Je gröber der Maßstab, desto augenfälliger zeichnen sich Verteilungs- und Ausbreitungsmuster auf Landschaftsebene ab, die klärende Hinweise zur Bedeutung von Orographie, Exposition, Seehöhe, Windrichtung oder Waldverteilung für Konstanz, Lückenbildung, Zusammenbruch oder Wiederbegründung von WaIdbeständen liefern können. Je feiner der Maßstab, desto besser lassen sich die Keimungs-, Wuchsund Konkurrenzbedingungen innerhalb der Pflanzendecke analysieren, denn letztlich werden die Chancen künftiger Baumgenerationen innerhalb der Moos- und Krautschicht - bereits im Sämlingsalter - entschieden! Je mehr Ebenen auf der Skala zwischen großräumiger Waldlandschaft und örtlichem Keimbett herangezogen werden, umso wirklichkeitsnäher gelingt die synoptische Verschneidung der Einzelbefunde. Da jede - noch so genaue - Erfassung des Vegetationszustandes nur eine Momentaufnahme aus einem permanenten Entwicklungsverlauf darstellen kann, sind Wiederholungsaufnahmen erforderlich, um Richtung, Tempo und Qualität des Naturgeschehens zielgerecht beschreiben zu können. Ein solches "Monitoring" stellt eine Schwerpunktaufgabe innerhalb der Nationalparkforschung dar, da Langzeitstudien zu natürlichen Prozessen nur in einem von Nutzungen dauerhaft freigestellten Schutzgebiet möglich sind. Allerdings sind künftige Ereignisse nicht vorhersehbar, 6


6 Windwurf Abteilung Hochseign 7998 weshalb die Auswahl der Methodik, der zu messenden Parameter und der Wiederholungsintervalle jeweils nur aus heutiger Sicht erfolgen kann. Die Diskussion der hier vorgelegten Ergebnisse aus Luftbildanalyse, Waldinventur und Vegetations-Monitoring kann auf vorausgegangene Berichte der Bayer. Landesanstalt für Wald und Forstwirtschaft (LWF), die Hefte der Wissenschaftlichen Schriftenreihe aus dem Nationalpark, das Sonderheft zum 25. bzw. die Festschrift zum 30. Gründungsjahr des Nationalparks zurückgreifen. Trotz des hohen fachlichen Niveauanspruchs haben sich die Autoren um eine allgemein verständliche Darstellung bemüht. Dabei zeigt sich das Problem, dass der Wortschatz unserer Alltagssprache der Vielseitigkeit der Naturphänomene nur unzureichend gerecht werden kann: Wenn z. B. die Einzelabschnitte natürlicher WaIdentwicklung als Zustand, Phase oder Strukturtyp beschrieben werden, so kann keiner dieser Begriffe der permanenten Dynamik des Naturgeschehens entsprechen, die weder Statik noch Stillstand kennt! Darüber hinaus wurzelt selbst das Fachvokabular zur Waldökologie in längst überholten Vorstellungen, sei es die anachronistisch anmutende Taxation bestimmter Baum- und Pflanzenassoziationen nach definierten Waldgeseilschaften, sei es die Erwartung langfristiger Stabilität ausgereifter Wälder in ihrer Alters- oder Schlussphase, der sogenannten "Klimax", entstammen diese Formulierungen ja der veralteten Erwartung zielgerichteter Entwicklungen der Vegetation zur Ausformung klar abgrenzbarer und von Natur aus vorgezeichneter Ökosysteme! Sehr viel realistischer - und wesentlich spannen der - sind aktuelle Einschätzungen zur Selbstorganisation von Ökosystemen, in denen Klima, Boden, Pilze, Pflanzen- und Tierarten in ununterbrochenem Wechselspiel z. B. den jeweiligen Waldbestand entwickeln. Nach diesem hochdynamischen Ansatz arrangiert sich die Vegetation zu jeder Zeit und an jedem Ort in individueller Ausprägung, weshalb Verallgemeinerungen spezieller Befunde - etwa aus dem Sukzessionsverlauf auf einer Beobachtungsfläche - nur bedingt möglich sind. Gleichzeitig wird klar, wie sehr die Nutzungsgeschichte der Vergangenheit zusammen mit den heutigen Umfeldbedingungen an der Waldentwicklung der Zukunft mitwirken, somit eine "Rückentwicklung" heutiger Wälder zu ihrer ursprünglichen Ausprägung unrealistisch ist. Die Interpretation naturgegebener Prozesse in den Wald-Lebensgemeinschaften muss aber noch ganz andere Schwierigkeiten überwinden, denn 7


7 Zerfal/sphase im Hochlagenwold, Hochfilzberg 7993 kaum ein Thema wurde derart mit emotionalen "Vorurteilen" überhäuft, wie die Natur des WaIdes : Ob schwärmerische Suche nach Vollkommenheit durch Harmonie und Gleichgewicht im Naturwald, ob ideal isierte Erwartung einer krisenfreien Waldnatur, die keine Katastrophen kennt, ob Wunschdenken nach dauerhaft nachhaltigem Waldwachstum oder illusorische Heilserwartung aus einer unberührten Natur, all diese populären Irrtümer sind als Diskussionsansatz für die Nationalpark- Forschung unbrauchbar. Vielmehr erscheint gerade eine unvoreingenommene, sachliche Deskription der Naturvorgänge als die beste Absicherung gegen Fehldeutungen und Öko-Dogmatik. Da entsprechende Forschungsergebnisse nicht nur Grundlage für ein zeitgemäßes Verständnis ökosystemarer Vorgänge sind, sie vielmehr auch eine Schlüsselrolle für das praktische Schutzgebiets-Management selbst einnehmen können, wurde dem Forschungsauftrag in Nationalparken innerhalb der Aufgabenliste nach IUCN 1994 eine hohe Priorität eingeräumt, vergleichbar dem Auftrag zu Umweltbildung und Besucherbetreuung. Das Heft 14 der wissenschaftlichen Schriftenreihe bildet einen Stein im Mosaik des Kenntnisgewinns, das den Einblick in das Naturgeschehen dieses Wald-Nationalparks fortlaufend verfeinern Es ist den Bewohnern des Bayerischen Waides, allen Freunden einer wilden Wald-Natur sowie all jenen Menschen gewidmet, denen der Erhalt unverfälschter Naturgüter ein wichtiges ku Itu relles An liegen ist. ~ i-~ Für das Redakt ions-team Dr. Wolfgang Scherzinger 8
8 Die Buchdruckermassenvermehrung im Nationalpark Bayerischer Wald von Marco Heurich, Arthur Reinelt und Lorenz Fahse 1. Einleitung Biologie des Buchdruckers Methoden Befliegung und Totholzkartierung Buchdruckerrelevante Klimakennwerte Ergebnisse Befallsentwicklung im Nationalpark Befallsentwicklung und Managementmaßnahmen Entwicklung buchdruckerrelevanter Klimakennwerte Potentielle Schwärm möglichkeiten Entwicklungsbedingungen für die Käfer Diskussion Habitatangebot Antagonisten Endogene Faktoren Konkurrenz...: Witterungsverlauf Synthese Zusammenfassung Summary Literaturverzeichnis
 verursachten Zugang an Totholzflächen sowie deren räumliche Verteilung zu dokumentieren.")
.")
9 Die Buchdruckermassenvermehrung im Nationalpark Bayerischer Wald Seit nunmehr 13 Jahren wird die WaIdentwicklung im Nationalpark Bayerischer Wald durch jährliche Luftbildserien dokumentiert. Ziel dieser Untersuchungen ist es vor allem, den durch den Buchdrucker Ups typographus) verursachten Zugang an Totholzflächen sowie deren räumliche Verteilung zu dokumentieren. Zusammen mit den Wald inventuren stellen die Luftbildserien die zentralen Komponenten für das Monitoring der natürlichen Waldentwicklung auf Landschaftsebene im Nationa lpark Bayerischer Wald dar. Weitere Monitoringprojekte, z. B. zur Ansprache des Belaubungszustandes von Laub- und Nadelbäumen greifen ebenfalls auf die Luftbilder zurück (BEU DERT und BREIT 2000). Darüber hinaus werden die bei der Totholzkartierung gewonnenen Informationen von Forschungsprojekten genutzt, die auf zeitnahe Informationen zur Waldentwicklung angewiesen sind. Neben der Bedeutung für die Forschung haben die Ergebnisse der Luftbildauswertung auch eine erhebliche Tragweite für politische Entscheidungen. Seitdem die Gewitterstürme 1983 und 1984 Waldbestände auf einer Fläche von 173 ha zu Boden warfen, ist der Buchdrucker und das durch ihn ausgelöste Absterben der alten Fichtenwälder das bestimmende Thema, mit dem sich die Nationalparkverwaltung auseinanderzusetzen hat. Obwohl nach einer Meinungsumfrage im Auftrag des WWF (WORLD WIDE FUND FOR NATURE) der Erhalt bestehender und die Gründung neuer Nationalparke von weiten Teilen der deutschen Bevölkerung unterstützt wird (95 % der Befragten finden Nationalparke wichtig, 70 % wollen, dass mehr Nationalparke ausgewiesen werden). sind die Mehrheitsverhältnisse vor Ort eher umgedreht. Vom Grundsatz her will man den Nationalpark zwar auch im Bayerischen Wald, über die Art und Weise, wie dort die Wälder behandelt werden sollen, gehen die Meinungen jedoch weit auseinander. Insbesondere die Politik der Nationalparkverwaltung, den Käfer in der Naturzone nicht zu bekämpfen, stieß auf scharfe Proteste in der örtlichen Bevölkerung. Mit dem Höhepunkt der Massenvermeh ru ng 1996 und 1997 erreichten auch die Auseinandersetzungen sowohl auf lokaler als auch nationaler Ebene ihren Höhepunkt. 10


10 Aber auch in Fachkreisen wurde lange über den richtigen Umgang mit dem Buchdrucker diskutiert, schließlich hatte man ja auch im weltbekannten Yellowstone Nationalpark eingegriffen, als sich dort 1987 riesige Brände ausbreiteten. Bis zu Feuerwehrleute waren damals mit schwerem Räumgerät eingesetzt, um die Brände zu bekämpfen. Im Nationalpark Bayerischer Wald entschieden sich die politisch Verantwortlichen - allen voran Ministerpräsident Dr. Edmund Stoiber - dafür, nicht einzugreifen und die Natur sich selbst zu überlassen. Diese Politik ist aber auch untrennbar mit dem Ziel verbunden, den konsequenten Schutz benachbarter Grundstücke vor Schäden aus dem Park sicherzustellen. Um die an den Nationalpark angrenzenden Privatwälder vor Buchdruckerbefall zu schützen, wurde die Randzone (= Bereich mit Waldmanagementl. entsprechend des örtlichen Gefahren- und Schadenspotentials und des zu erwartenden starken Befa IIsd rucks, neu abgegrenzt und eine aufwendige Befallskontrolle eingeleitet. Da es Anfang der 90er Jahre keinen größeren Windwurf oder Schneebruch gab, wird bis heute vor allem die Frage nach dem Auslöser für die Massenvermehrung diskutiert. Vereinfacht stehen sich zwei Lager gegenüber. Das eine Lager vertritt eine "WaIdschadenshypothese", die davon ausgeht, dass die Bäume durch Umweltverschmutzung so stark geschädigt waren, dass sie vom Buchdrucker befallen und zum Absterben gebracht werden konnten. Das andere Lager vertritt die Auffassung, dass es sich bei dem großflächigen Absterben der Altbestände um einen natürlichen Vorgang handelt, der im Naturraum Böhmerwald in gewissen Zeitabständen wiederkehrt. Ziel dieser Arbeit ist es, die aktuelle Entwicklung des Buchdruckers zu dokumentieren und den aktuellen Wissensstand über die Populationsdynamik des Buchdruckers im Nationalpark Bayerischer Wald zu diskutieren. Aufgrund der großen Bedeutung, die der Witterung in diesem Zusammenhang beigemessen wird (Zierl 1972, USDA FDREST SERVICE 2001), wurde der Witterungsverlauf der vergangenen 20 Jahre aus der "Sicht" des Buchdruckers analysiert :===========
 Durch die Froßgänge des Borkenkäfers wird der Nährstofftronsport im Boum unterbrochen.")
 befollenen Bäumen ous der Vogelperspektive. Abb.4(m. l.")
 Brutbild des Buchdruckers; senkrecht: Muttergänge woagrecht: Lorvengänge Abb. 6 (u. l.")
11 Abb. 2 (0. r.) Durch die Froßgänge des Borkenkäfers wird der Nährstofftronsport im Boum unterbrochen. Die Nadeln sterben ob und verfärben sich rot. Abb. 3 (0. 1.) Gruppe von frisch (rot) bzw. in den Vorjahren (grou) befollenen Bäumen ous der Vogelperspektive. Abb.4(m. l.) Einbohrlächer mit ousgeworfenem Bohrmehl om liegenden Stomm. Abb. 5 (m. r.) Brutbild des Buchdruckers; senkrecht: Muttergänge woagrecht: Lorvengänge Abb. 6 (u. l.) Am Stommfuß befollener Bäume sammelt sich dos chorkteristische broune Bohrmehl... Abb. 7 (u. r.)... und es bildet sich ein Teppich aus grünen Nodeln. 12


12 2. Die Biologie Des Buchdruckers Der Buchdrucker ist trotz seiner geringen Größe der zur Zeit auffälligste Akteur in den Wäldern des Nationalparks Bayerischer Wald. Es handelt sich um einen 4 bis 5,5 mm kleinen, walzenförmigen Käfer von rot- bis schwarzbrauner Farbe. Erkennungsmerkma l des Buchdruckers sind die zu ei nem Absturz abgeschrägten Flügeldecken, die an beiden Außenrändern mit jeweils vier Zähnen versehen sind. Deshalb heißt der Buchdrucker auch Großer Achtzähniger Fichtenborkenkäfer. Bevorzugter Wirtsbaum der Käfer ist die Fichte. Andere Baumarten wie Waldkiefer und Tanne werden nur in Ausnahmesituationen, z. B. während Massenvermehrungen befallen. Im Frühjahr, wenn die Lufttemperatur C überschreitet, meist im April oder Mai, verlassen die Käfer ihre Winterquartiere, fliegen zu geeigneten Fichten und bohren sich in deren Rinde ein. Zuerst schwärmen die Männchen. Nachdem sie sich eingebohrt haben, legen sie eine sogenannte Rammelkammer an und versuchen mit Hilfe von Duftstoffen (Pheromonen) Weibchen anzulocken. Nach der Befruchtung nagen 1 bis 3 Weibchen, ausgehend von der Rammelkammer, im Durchschnitt 7 bis 8 cm lange Muttergänge, an denen beidseitig 30 bis 60 weiße Eier in mehr oder weniger regelmäßigen Abständen in Einischen abgelegt werden. Das bei der Anlage von Rammelkam mer und Muttergang anfallende braune Bohrmehl wird aus dem Einbohrloch geworfen und ist ein wichtiges Kennzeichen für frisch vom Buchdrucker besiedelte Bäume. Nach ein bis zwei Wochen schlüpfen aus den Eiern weiße Larven, die sich vom Bast und Kambium, das sind die lebenden Gewebe unter der Baumrinde, ernähren. Die Larvengänge verlaufen leicht geschlängelt quer zum Muttergang und werden mit dem Wachstum der Larven zunehmend breiter. Das sich aus den Fraßgängen der Mutterkäfer und Larven ergebende Brutbild ähnelt einem aufgeschlagenen Buch und gab dem Käfer seinen Namen. Für die Bäume ist die Anlage der Brutsysteme jedoch lebensbedrohlich, da durch sie der Nährstofftransport (Kohlenhydrate) zu den Wurzeln unterbrochen wird, was dazu führt, dass die Nadeln nicht mehr ausreichend versorgt werden können, sich röten und letztendlich abfallen.je nach Witterung fressen die Larven drei bis vier Wochen unter der Rinde, bis ihre Entwicklung abgeschlossen ist und sie sich verpuppen. Nach ein bis zwei Wochen Puppenruhe schlüpfen die Jungkäfer. Sie sind auf Grund ihrer hellbraunen Färbung leicht von den Altkäfern zu unterscheiden. Die Entwicklung bis zum gesch lechtsreifen Tier dauert weitere zwei bis drei Wochen, so dass sich für die Entwicklung einer Generation eine Dauer von sieben bis elf Wochen ergibt. Nach einem Regenerationsfraß können die Altkäfer erneut zur Brut schreiten; man spricht dann von sog. Geschwisterbruten.ln einem Jahr können sich somit je nach Witterungsverlauf ein bis zwei (drei) Hauptgenerationen und zwei Geschwisterbruten entwickeln, was eine enorme Vermehrungsrate möglich macht. Unter günstigen Umweltbedingungen, wie relativ hohen Temperaturen (optimal 25 bis 30 C; COElN 1997) und ausreichend Brutraum in Form von Windwurf oder Schneebruch, können Käferpopulationen regelrecht "explodieren" (NIERHAUS-WUNDERWAlD 1992, SCHWERDTFEGER 1981). Eine Faustregel sagt, dass, ausgehend von einem mit Käfern besetzten Baum, zehn weitere Bäume befallen werden können. Abb.8 Ob wahl der Buchdrucker recht unscheinbar ist, fü hrt sein massenhaftes Auftreten zu tiefgreifenden Veränderungen im Ökasystem Wald. 13


13 3.1 Befliegung und Totholzkartierung Seit 1988 werden jährlich Luftbildserien erstellt, um die Entwicklung des Buchdruckers oder vielmehr die Fläche der durch ihn abgetöteten Bestände zu dokumentieren. Aufgrund der ungünstigen Witterung im Jun i und Juli 2000 musste die seit 1989 kontinuierliche Zeitreihe von Sommerbefliegungen unterbrochen und eine Befliegung im Herbst durchgeführt werden. Bei der Befliegung am wurden wie bereits in den Vorjahren überlappende CIR-Luftbilder im Maßstab 1 : des gesamten Nationalparkgebietes angefertigt. Die Kartierung der abgestorbenen und ausgeräumten Flächen beschränkte sich auf das Rachel-Lusen-Gebiet und erfolgte durch die Gesellschaft für angewandte Hydrologie und Kartographie mbh. Mit Hilfe eines Stereoskops wurden alle Totholzflächen, die seit der letzten Befliegung entstanden sind, identifiziert. Einzelbäume blieben unberücksichtigt, da die Erfassungsgrenze bei Gruppen von mindestens fünf abgestorbenen Bäumen liegt. Um den neuen Befa ll eindeutig von dem der vorausgegangenen Jahre unterscheiden zu können, wurden die bereits in den Vorjahren kartierten Flächen zusammen mit Orientierungshilfen wie Forstwege und Schneisen im Maßstab des Luftbildes auf die Folien gedruckt. Diese Folien wurden auf die Bilder gelegt und partiell eingepasst. Nach der Kartierung wurden die Folien gescannt, georeferenziert und anschließend am Bildschirm digitalisiert. Die weitere Verarbeitung erfolgte mit der GIS-Software ArcView. Da sich die im Frühsommer befallenen Bäume erst etwa ab Anfang August verfärben, wird bei Befliegungen im Jun i und Juli immer der Zugang an Totholzflächen des Vorjahres erfasst. Die bei der Käferbekämpfung entstandenen Kahlflächen können sofort im Luftbild erkannt werden, so dass bei diesen Flächen die aktuelle Entwicklung bis zum Zeitpunkt der Befliegung dokumentiert wird. Im Gegensatz dazu wird bei einer Herbstbefliegung weitgehend der aktuelle Stand der Totholzentwicklung dokumentiert. Dies hängt damit zusammen, dass die im Frühsommer befallenen Bäume im Herbst bereits rot verfärbt sind bzw. ihre Nadeln schon verloren haben und somit im Luftbild eindeutig identifiziert werden können. Ein Nachteil der Herbstbefliegung ist, dass sich die im Spätsommer befallenen Fichten zumeist erst im Winterhalbjahr verfärben und deshalb nicht erfasst werden können. Um die Größenordnung des Spätsommerbefalls quantifizieren zu können, müsste direkt nach einer Herbstbefliegung eine Sommerbefliegung durchgeführt werden. Das ist bisher noch nicht geschehen. Darüber hinaus ist auch zu erwarten, dass der Spätsommerbefall von Jahr zu Jahr witterungsbedingt stärker variiert und deshalb kaum abgeschätzt werden kann. Abb. 9 Nachdem sich die Borkenkäferanfangs vor allem in den fichtendominierten Hochlagenwäldern ausgebreitet hoben, werden in den letzten Jahren zunehmend auch die Wälder in den Hanglagen befallen. Um trotz der unterschiedlichen Befliegungstermine einen einheitlichen Bezug herzustellen, wurde bei der Darstellung der Ergebnisse nicht wie bisher das Befl iegungsjahr, sondern das Befallsjahr als Bezugszeitraum verwendet. Die dadurch verursachten Unschärfen bei der Zuordnung ausgeräumter Flächen werden in Kauf genommen. 14
 sowie seit 1997 (WEISSBACHER 1998, 1999, 2000) vorliegen, wurden biologisch bedeutsame")


14 3.2 Buchdruckerrelevante Klimakennwerte Ziel dieser Analyse war es, den Witterungsverlauf der vergangenen Jahre aus der "Sicht" des Buchdruckers zu untersuchen. Da Felddaten zur Entwicklung der Buchdrucker vornehmlich aus den Jahren 1989 bis 1992 (SCHOPF und KOEHLER 1995) sowie seit 1997 (WEISSBACHER 1998, 1999, 2000) vorliegen, wurden biologisch bedeutsame Kennwerte wie Schwärmbeginn, Schwärmtage und Entwicklungsgeschwindigkeit auf der Basis meteorologischer Daten rekonstruiert bzw. abgeschätzt. Weil sich die klimatischen Bedingungen im Rachel-Lusen-Gebiet schon auf Grund des großen Höhengradienten von mehr als 760 m erheblich unterscheiden, erfolgte die Analyse der relevanten Klimaparameter von drei Wetterstationen, welche die Höhenzonen des Nationalparks repräsentieren (ELLING et ai.1987). Die Wetterstation Arber befindet sich auf dem Gipfel des gleichnamigen Berges in 1457 m Meereshöhe. Sie repräsentiert die klimatisch ungünstigsten Verhältnisse der Gipfellagen in der Hochlagenzone. Die Wetterstation Waldhäuser befindet sich in einer Meereshöhe von 940 m im Übergangsbereich zwischen Oberer und Unterer Hanglage, deren Grenze entlang der 900 m Isolinie verläuft. Diese Wetterstation befindet sich auf einem nach Westen exponierten Hang mit 0-4 Grad Neigung. Die dort beobachteten Werte stehen stellvertretend für das Klima der Hanglagen. Das durch Kaltluftstau und eine hohe Strahlungsfrostgefahr geprägte Klima der Tallagen wird durch die Wetterstation Klingenbrunn-Bahnhof am Fuß des Rachelmassivs charakterisiert. Sie befindet sich auf einer Verebnung in einer Meereshöhe von 756 m. Eine Kurzübersicht über die Klimaverhältnisse in den vier Höhenstufen gibttabelle 1. Die folgenden Klimakennwerte besitzen eine große Bedeutung für die Entwicklung der Buchdrucker: Erster Tag mit einer Maximaltemperatur von über 20 oe Bei Temperaturen von mindestens 20 oe ist es dem Käfer möglich, zu schwärmen. Der erste Tag mit einer Maximaltemperatur über 20 oe stellt somit den frühestmöglichen Beginn für den Entwicklungszyklus des Buchdruckers dar. Die in der Fachliteratur angegebenen Temperaturschwellenwerte (gemessen im Freiland) für den Beginn Tab. 1 Kurzübersicht über die klimatischen Verhältnisse in den vier Hähenstufen des Natianalparks (nach ELLINGetal. 1987). Höhenstufe Mittlere Höhenbereich Jahresmittel der Jahressumme des Spätfrostgefahr Nebel- Schnee- Niederschlags m NN Lufttemperatur oe (Art der Fröste) niederschlag deckenzeit mm Monate Hochlage Untergrenze bei gering (Strahlung Advektion) ca bedeutend ca. 7 Obere Hanglage Obergrenze zwigering nach oben schen ca (Advektion) zunehmend Untergrenze 900 ca. 5-6 Obergrenze 900 gering Untere U nte rg re nze (Strahlung + Hanglage 900 Advektion) ca unbedeutend ca. 5 Flache Zone im unteren Bereich von m Tallage mit Mächtigkeit über groß nächtlichem der Talsohle. Oberin höheren Lagen (Strahlung) Ka Itl uftsta u grenze zwischen weniger 690 und 1120 m ca unbedeutend ca
15 des Schwärmfluges schwanken zwischen 15 und 20 oe (ADlUNG 1979, LOBINGER 1997, NIERHAUS WUNDERWALD 1992 zit. in NETHERER 2000), wobei sich die Mehrzahl der Autoren für den höheren Schwellenwert ausspricht (SCHWERDTFEGER 1981, HARDING und RAVEN, 1985 zit. in NETHERER 2000, NETHERER 2000). Auch die im Nationalpark ermittelten Schwellenwerte, die mit Temperatursensoren in unm ittelbarer Umgebung von Pheromonfallen auf Bekämpfungsflächen im Randbereich ermittelt wurden, deuten auf Schwellentemperaturen von ca. 18 bis 19 oe hin (NUESSlEIN et al. 1999). Addiert man zu diesem Wert die durchschnittliche Differenz der Maximaltemperaturen zwischen Waldbestand und Freifläche (Wetterstation), die nach Untersuchungen von JEHl (pers. Mitteilung) in den Hochlagen des Nationalparks ca. 1,8 oe beträgt und beachtet die Exposition der Stationen, ergibt sich eine Temperaturschwelle, die recht gut mit der in der Literatur genannten Freilandtemperaturvon 20 oe übereinstimmt. Es wird zwar immer Buchdrucker geben, die an klimatisch begünstigten Kleinstandorten schon früher schwärmen; zur Terminierung des allgemeinen Schwärmfluges dürfte sich die TemperaturschweIle von 20 oe jedoch gut eignen. Der aus den Daten der Wetterstationen abgeleitete potenzielle Schwärmbeginn ist als Schätzwert für den tatsächlichen Schwärmbeginn anzusehen. Letzter Tag mit Schneebedeckung Das Abschmelzen der Schneedecke ist die Voraussetzung dafür, dass der im Boden überwinternde Anteil der Käferpopulation zum Schwärmflug ansetzen kann. Teilweise kommt es nach dem ersten Abschmelzen der Schneedecke zu erneutem Schneefall: Wenn während des dazwischen liegenden Zeitraumes ohne Schneebedeckung Maximaltemperaturen von über 20 oe gemessen wurden, gilt der Tag nach dem ersten Abschmelzen des Schnees als erster Tag ohne Schneedecke, ansonsten der erste Tag nach dem letzen Abschmelzen des Schnees. Anzahl der Tage mit einer Maximaltemperatur von über 20 oe in den Monaten April und Mai Da die Käferpopulation meist mehrere Tage benötigt, bis alle Individuen ihren Schwärmflug durchgeführt haben, besitzt nicht nur der erste potentielle Schwärmtag, sondern auch die Anzahl dieser Tage im Frühling eine große biologische Bedeutung. Temperatursumme in Stundengraden "Temperatursummen in Stundengraden repräsentieren jene Energiemengen, welche für die Vollendung einzelner Entwicklungsabschnitte bzw. für die Gesamtentwicklung des Buchdruckers benötigt werden. Hierbei sind aber nur die effektiven Werte, d. h. jene, die Einfluss auf die Entwicklung nehmen, von Relevanz. Der Entwicklungsnullpunkt ist mit jener Temperatu r defin iert, unter der kein Entwicklungsfortschritt mehr zu beobachten ist" (eoeln 1997). Er wurde, wie auch die für die Entwicklung notwendigen Temperatursummen, experimentell bestimmt und liegt nach eoeln et al. (1996) bei 11,8 oe. Die Berechnung dertemperatursumme erfolgt nach folgender Formel (verändert nach eoeln 1997): Teff = I (Thv,i - ENP ) * 24 T", : effektive Temperatursumme T: Tagesmittel der Kambialtemperatur ENP: Entwicklungsnullpunkt (= 11,8 oe) Entsprechend dieser Formel wäre es für die Berechnung der effektiven Temperatursumme notwendig, die Kambialtemperatur lebender Fich- 16


16 Effektive Temperatur- Entwicklungsstadium summe (Stundengrade) Ei larve Puppe Jugendentwicklung tenstämme bei unterschiedlichem Beschirmungsgrad zu bestimmen. Erst durch eine Temperaturmessung in der Bast- oder Kambialschicht können die Entwicklungsbedingungen für die Buchdrucker optimal abgebildet werden. Da Messungen der Kambialtemperatur aus dem Untersuchungsgebiet erst seit 2000 vorliegen, mussten statt der Tagesmittel der Kambialtemperatur die Tagesmittel der Lufttemperatur in die Berechnungen eingehen. Bei dieser Vereinfachung ist allerdings zu beachten, dass die Kambialtemperatur in der Regel deutlich über der Lufttemperatur liegt, da sie durch direkte Einstrahlung beeinflusst wird. Je geringer der Bestandesschluss, desto größer ist die Differenz zwischen Kambial- und Lufttemperatur. Nur in dicht geschlossenen Beständen folgt die Kambialtemperatur dem Verlauf der Lufttemperatur, weil dort die Stammoberflächen nicht besonnt sind. Damit stellen die aus der Lufttemperatur berechneten Temperatursummen das ungünstigste Szenario (worst-case) für die Käferentwicklung dar. Die realen Entwicklungsbedingungen dürften günstiger gewesen sein, da sich die Entwicklung der Buchdrucker vor allem in der Umgebung von alten Totholznestern oder ausgeräumten Flächen, also in gut durchsonnten Bereichen mit höheren Kambialtemperaturen, abspielt. Tob. 2 zeigt die effektive Temperatursumme der einzelnen En twicklungsstodien vo n Ips typographus bei einem En twicklungsnullpunkt von II,BoC (COELN et , COELN 199B). 17



17 4.1 Befallsentwicklung im Nationalpark 'iä' 700 ;5 g' 600 '" c:n ~ c: '''' 'i':i r ~ Ausgeräumte Borkenkäferflächen im Randbereich 1- " <= ~ Abb.l0 Jährlicher Zugang an Totholzflächen im Rachel- Lusen-Gebiet {ohne Erweiterungsflächen}; die Erhebungen bis 1993 enthalten nur Totholzflächen ab 0, 1 ha Größe {Änderung des Aufnahmeverfahrens 1993}. Abb. ll Entwicklung der mittleren Flächengrößen als Boxplo t-darstellung. Befliegungsjahr 409 Seit der letzten Befliegung im Herbst 1999 (409 ha Befallsfläche) hat sich die Fläche der vom Buchdrucker befallenen Baumbestände im Nationalpark Bayerischer Wald um 605 ha erhöht. Diese Fläche umfasst neben den Befallsflächen in der Naturzone auch die in Folge der Käferbekämpfung ausgeräumten Flächen in der Randzone des Parks. Erstmals seit drei Jahren wurde damit wieder eine Zunahme der jährlich befallenen Fläche beobachtet. Nur 1996 wurde mit 827 ha eine noch größere Fläche neu befallen. Damit sind die durch Buchdruckerbefall abgestorbenen Fichtenbestände auf eine Fläche von mittlerweile ha angewachsen. Das sind 29 Ofo oder fast ein Drittel der gesamten Waldfläche des alten Nationa I parkgebietes. I ~ I'j I t I ü:i 7500 I ~ i c: 5000 I ~ ::: 2500 <LI!'B I ~ I OL- -- ~ ~ ~~ Befallsjahr Die jährliche Entwicklung des Medians der Größe der Befallsflächen und des Gesamtbefalls verlaufen in etwa parallel: Als der Käferbefall Ende der 80er und Anfang der 90er Jahre rückläufig war, sank auch der Median der Größe der Neubefallsflächen. Die Streuung (lnterquartilbereich) der Werte um den Median des Neubefalls wurden ebenfalls geringer. Mit dem Ansteigen der Neubefallsfläche bis 1996 wuchsen auch der Median der Flächen und die Streuung der Werte wieder an konnte die größte, innerhalb eines Jahres entstandene zusammenhängende Neubefallsfläche beobachtet werden. Sie war durch das Zusammenfließen von Befallsherden aus dem Jahr 1995 zwischen Lusen und Moorkopf entstanden und hatte eine Größe von 239 ha. Ein diese Fläche umschließendes Rechteck hat eine Ausdehnung von 2,8 mal 4,4 km. Als anschließend die Totholzflächenentwicklung rückläufig war, verringerten sich wiederum der Median der Flächengröße und die Streuung der Werte, allerdings wie beim Anstieg mit einer Zeitverzögerung von einem Jahr stand einer Zunahme der Neubefallsfläche ein Rückgang des Medians und der Streuung gegenüber. Auffällig war im Jahr 2000 besonders der starke Anstieg der Anzahl von Befallsherden von 884 auf Nur 1993 wurde mit eine noch größere Zahl von Befallsherden beobachtet. In den Hochlagen nahm die vom Buchdrucker befallene Fläche im Jahr 2000 um fast 220 ha zu und erreichte damit annähernd das Niveau der drei vorangegangenen Jahre. Inzwischen sind in dieser Höhenzone 85 Ofo der Waldbestände, das sind ha, abgestorben. Der Befall konzentriert sich auf die noch verbliebenen Bestände am östlichen und westlichen Ausläufer der Hochlagen in den Gebieten um den Siebensteinkopf und im Bereich westlich des Rachelgipfels. Ein Befallsschwerpunkt ist auch im Bereich nördlich der Schwarzbachklause zu finden. Die Befallsentwicklung in den Hochlagen ist insofern erstaun- 18



18 lieh, da der Neubefall trotz ständig zurückgehender Restbestockung mit unverminderter Geschwi ndigkeit fortschreitet. In der oberen Hanglage stieg die neu befallene Fläche von 137 auf 243 ha, also um 77 % an. Damit erreichte der Neubefall in dieser Höhenstufe seinen höchsten Wert und übertrifft auch den Neubefall in allen anderen Höhenlagen. Mittlerweile sind insgesamt ha Altbestände abgestorben, das entspricht rund 26 Ofo der gesamten Waldfläche dieser Höhenstufe. Der hier auftretende Käferbefall hat seinen Ursprung zum überwiegenden Teil in den Hochlagen. Starker Befall tritt in dieser Höhenzone vor allem im Rachelnordkar, in der Rachelseewand, südlich des Rachelsees, im Distrikt Fuchsenhöhl, im Distrikt Sagwasserhäng, im Distrikt Reschhäng, in den Bärnriegeln und nördlich destier-freigeländesauf. In der unteren Hanglage stieg der Neubefall von 43 auf 69 ha, also um 37 % an. Dies ist der dritthöchste Wert, der für diese Zone bisher ermittelt wurde. Damit sind in der unteren Hanglage bisher insgesamt 355 ha Fichtenaltbestände abgestorben, das sind ca. 10 Ofo der Waldfläche. Charakteristisch für den Befall in dieser Zone sind viele 80 ~ 70 't:i '" C :.,. 1;; 60 al '" ~ '" c 50 ~ '" 40 (;... VI 30 '" Cl ~.,. OE 20 c « HochJage -.-. Untere Hangjage relativ kleine Neubefallsflächen. Große Flächen mit frischem Befall sind in dieser Höhenzone nur durch das Zusammenlaufen von Totholzgebieten nordöstlich von Jägerfleck entstanden. In den Tallagen wuchs die Totholzfläche seit der letzten Befliegung um 75 ha. Das ist der höchste Wert seit Beginn der Käfergradation. Im Vorjahr lag die Neubefallsfläche in dieser Höhenzone nur bei 36 ha. Insgesamt sind dort inzwischen 309 ha abgestorben, das entspricht rund 11 Ofo der Waldfläche dieser Höhe~zone. Die Befallsschwerpunkte lagen im Talkessel nördlich von Guglöd Obere Hangjage - TaIJage i Abb.12 Jährlicher Zugang an Tathalzflächen im Rache/-Lusen-Gebiet (ohne Erweiterungsflächen) differenziert nach Hähenstufen.. ~,.,...s:: : -;:; 500.,. c Cl I ~ 400 ' '5'" I ~ 300..s::.~ -.::: 200..s:: ~ 100 Abb r ~ Entwicklung der Fläche 600 TalJage Hochjage abgestarbener Bestände im Rache/-Lusen-Gebiet (ohne Erweiterungsflächen), differenziert nach Hähenstufen. o ~ BefalJsjahr 19






19 4.2 Befallsentwicklung und Managementmaßnahmen Abb.14 Insbesondere bei der Schneebruchaufarbeitung und auf empfindlichen Böden werden häufig Rückepferde eingesetzt. In der Naturzone des Rachel-Lusen-Gebietes darf sich die Natur nach ihren eigenen Gesetzmäßigkeiten entwickeln. Hier wird, abgesehen von Verkehrssicherungsmaßnahmen und Wegeunterhalt, nicht lenkend in die natürlichen Abläufe der Waldentwicklung eingegriffen. Die Naturzone umfasst im Rachel-Lusen-Gebiet eine Fläche von etwa ha. Zum Schutz der an den Nationalpark angrenzenden Wirtschaftswälder wird der Buchdrucker allerdings in einer ca ha großen Randzone bekämpft. Je nach örtlichen Gegebenheiten hat diese Zone eine Tiefe von mindestens 500 bis 1500 m. 11 b muss die Buchdruckerpopulation in einem Übergangszeitraum bis zum Jahre 2017 kontrolliert werden. Im Falkenstein-Rachel-Gebiet wird der Buchdrucker bislang auf der überwiegenden Fläche kontrolliert. Die Naturzone hat dort bisher nur eine Größe von ha, soll aber in den nächsten Jahren schrittweise ausgeweitet werden. In den Hochlagen (Entwicklungszone 11 a) und in der zu deren Schutz ausgewiesenen Entwicklungszone DEnklaven Naturzone D EntlNickiungszone 11 a D EntlNickiungszone 11 b _ EntlNickiungszone 11 c D Randbereich _ Erholungszone Die Bekämpfung des Käfers erfolgt auf Basis eines integrierten, den örtlichen Gegebenheiten angepassten Konzeptes. Auf gefährdeten Böden wie Weich böden oder Felspartien, aber auch bei verstreutem Holzanfall in den Hochlagen werden die befallenen Bäume an Ort und Stelle entrindet und verbleiben im Wald. In schwierig bringbaren Lagen wird auf eine Erschließung mit Forststraßen verzichtet. Die befallenen Bäume werden unter Einsatz von Hubschraubern an zentrale Aufarbeitungsplätze geflogen. Auf den verbleibenden Flächen werden die befallenen Bäume motormanuell aufgearbeitet und mit Rückeschleppern sowie in schwächeren Beständen, insbesondere bei der Schneebruchaufarbeitung, wo möglich, mit Pferden an die Forststraßen transportiert. Von dort werden sie rasch mit Lkw 's abtransportiert oder, wenn dies nicht fristgerecht möglich ist, an Ort und Stelle maschinell entrindet. Karte 1 Managementzonen im Nationalpark Bayerischer Wald. Zwischen 1991 und 1996 erhöhte sich der Einschlag von "Käferholz" im Rachel-Lusen-Gebiet kontinuierlich von 525 auf fm wurde dort der Randbereich als Reaktion auf den steigenden Populationsdruck durch Befallsherde in der unmittelbar angrenzenden Naturzone vergrö- 20



20 ßert. Dadurch stieg der "Käferholzeinschlag" auf fast fm.1998 erreichte der Holzeinschlag mit knapp fm seinen höchsten Wert. Im Jahr 2000 wurden nur noch knapp fm Käferholz eingeschlagen. Allerdings traten in den letzten bei den Wintern starke Schneebruchereignisse auf, die dazu beitrugen, dass der Gesamteinschlag nur wenig zurückging konzentrierte sich der Schneebruch auf die Höhenlagen zwischen 900 und m. Insgesamt wurden fm eingeschlagen. Im Jahr 2000 mussten fast fm Schneebruch aufgearbeitet werden. Diesmal waren die unteren und mittleren Lagen zwischen 600 und m betroffen. Regulärer Einschlag spielte im Rachel-Lusen-Gebiet innerhalb des betrachteten Zeitraumes mit im Durchschnitt weniger als fm pro Jahr nur eine untergeordnete Rolle. Auch im Falkenstein-Rachel-Gebiet stieg der "Käferholzeinschlag" bis 1995 kontinuierlich an, verdoppelte sich 1996 auf fm, ging 1997 leicht zurück und erreichte ebenfalls 1998 sein Maximum mit fm. Anschließend war der Holzeinschlag wieder rückläufig und erreichte im Jahr 2000 mit fm den niedrigsten Wert seit Allerdings erhöhte sich der Gesamteinschlag seit 1997, dem Jahr der Nationalparkerweiterung, von Jahr zu Jahr. Zwar wurde die reguläre Nutzung, die 1996 noch knapp fm erreichte, auf unter fm vermindert; gleichzeitig stiegen aber die Zwangsanfälle durch Sturm und Schneebruch stark an. So mussten fm Sturmwurf und fm Schneebruch aufgearbeitet werden. Im Jahr 2000 betrugen die Zwangsnutzungen durch Windwurf nur fm, allerdings mussten fm Schneebruch aufgearbeitet werden. Interessant ist die Beobachtung von NUESSLEIN (1996), dass der "Käferholzeinschlag" im Falkenstein-Rachel-Gebiet synchron zu den Absterbe o Regulär o Schneebruch Sturm f- 2 Borken käfer ci E... ~ C"I :<: u VI c: '0; N "0 :I: Rachel-Lusen -Gebiet 0-'-"----'----' ,--., , , ~ E ~ C"I :<: ~ c: '0; N "0 :I: D Regulär o Schneebruch. Sturm Borken käfer = ---- I-~, :;;;-- I _ I ~ I r- I Falkenstein-Rachel-Gebiet I ~. ~ ~ -., prozessen im Rachel-Lusen-Gebiet erfolgt. Bisher gibt es nur zwei Ausnahmen von dieser Beobachtung 1998 und ging der Befall im Rachel-Lusen-Gebiet leicht zurück, während der Holzeinschlag im Falkenstein-Rachel-Gebiet anstieg, im Jahr 2000 war es umgekehrt. Deutliche Unterschiede zeigen sich auch im Ausmaß der Veränderung gegenüber dem Vorjahr. Beispielsweise stieg der "Käferholzeinschlag" im Falkenstein-Rachel-Gebiet nie um mehr als 200 % an, während der Zuwachs im Rachel-Lusen-Gebiet bis zu 843 % erreichte. Abb. 15 u. 16 Jährlicher Holzeinschlag im Rache/-Lusen-Gebiet (seit um Flächen des Forstamtes Regen erweitert) und im Falkenstein-Rachel-Gebiet (bis ehemaliges Forstamt Zwiesel), differenziert nach Aufarbeitungsursache. 21


21 Durch die Holzentnahme im Zuge der Käferbekämpfung entstanden nach und nach Kahlflächen. Mittlerweile haben diese geräumten Flächen im Rachel-Lusen-Gebiet eine Größe von insgesamt 170,2 ha erreicht. Die größte Fläche liegt in der Abteilung Geiermais im Reschbachtal (9,5 halo Der Zugang an neuen Kahlflächen betrug im Jahr 2000 etwa 23 ha und lag damit weit unter dem Maximalwertvon 85,4 ha im Jahre Die Bekämpfungsschwerpunkte lagen im Oberen Reschbachtal, entlang der Buchwaidstraße, im Distri kt Ohä ng westl ich der Ortschaft Wa Id häuser und im Distrikt Rachelhäng. Abbildung 17 zeigt die Entwicklung des Buchdruckerbefalls in Abhängigkeit von der Entfernung zur Naturzone. Während 1997, dem Jahr der Vergrößerung des Randbereiches, der Befall relativ gleichmäßig verteilt war, ja mit größerer Entfernung von der Naturzone sogar noch anstieg, entstand 1998 bereits ein leichtes Gefälle zwischen den unmittelbar an die Naturzone angrenzenden und etwas weiter entfernten Bereichen der Randzone zeigten sich dann die Erfolge der intensiven Befallskontrolle (ein Kontrolleur je 200 ha Holzbodenfläche) und des raschen Holzeinschlags: Das Befallsniveau in der Randzone ging zurück und in einer Entfernung von mehr als 600 m zur Naturzone trat nur noch geringer Neubefall auf. Auch die neuesten Zahlen (2000) belegen deutlich den Erfolg der Käferbekämpfung im Randbereich. Trotz des stärkeren Befallsdrucks aus der Naturzone heraus ist es gelungen, den Neubefall in der Randzone auf einen Bereich von 500 m zu begrenzen und damit den Schutz angrenzender Wi rtschaftswälder zu gewährleisten. 3,5 Abb.17 Flächenanteil ausgeräumter Borkenkäferflächen in der Randzone in Abhängigkeit von der Entfernung zur Naturzone im Rache/-Lusen-Gebiet. ~ 3,0 c:: 2,5 cu.c: <J "" 't ~ 2,0 ~ "" 0;... E :::J 1,5 :~ cu c:n '" :::J '" '0; 1,0... c:: <t 0,5 0, , Entfernung von der Naturzone (m) 22
")

22 23 (Abb. 18)


23 4.3 Entwicklung buchdruckerrelevanter Klimakennwerte Potenzielle Schwärmmöglichkeiten Diagramm 19 zeigt die Entwicklung des potenziellen Schwärmbeginns seit In der ersten Hälfte der betrachteten Periode wird der potenzielle Schwärmbeginn in den Hochlagen etwa einen Monat später als in den Hanglagen erreicht; auch zwischen Hang- und Tallagen bestehen deutliche Unterschiede. In den 90er Jahrenjedoch tritt der potenzielle Schwärmbeginn besonders in den Hochlagen immer früher ein, in den Hangund Tallagen beginnt das Schwärmen mehr oder weniger zeitgleich. Im Jahr 2000 kam es erstmals zu einem gemeinsamen potenziellen Schwärmbeginn in allen Höhenstufen. lichkeiten zwischen 1989 und Damals konnten die Buchdrucker erst Ende Juni ihre Winterquartiere verlassen. Die Tendenz zu einem früheren Schwärmbeginn ist schwach signifikant. In den Hanglagen wurde der früheste Tag mit Maximaltemperaturen über 20 oe am 21. April (1983 und 1996), der späteste am 22. Juni (1991) gemessen. Die Differenz zwischen diesen Werten beträgt 62 Tage. Auch hier findet sich eine Periode späterer Schwärmmöglichkeiten Ende der 80er und Anfang der 90er Jahre, die allerdings nicht so ausgeprägt ist, wie in den Hochlagen. Aus Abbildung 19 ist eine Tendenz zu einem früheren potenziellen Schwärmbeginn ersichtlich, sie konnte jedoch nicht statistisch abgesichert werden. In den Hochlagen wurde der früheste Tag mit Maximaltemperaturen über 20 oe am 23. April (2000) gemessen, der späteste Zeitpunkt war der 7. Juli (1991). Die Spannweite beträgt 75 Tage. Auffällig ist in dieser Höhenstufe vor allem die Periode sehr später potenzieller Schwärmmög- Die frühesten potenziellen Schwärmmöglichkeiten herrschten in den Tallagen. Im Jahr 1981 wurden bereitsam 1. April Temperaturen von über 20 oe erreicht. Wie bereits in den anderen Höhenstufen konnte der späteste Beginn des potenziellen Schwärmfluges 1991 beobachtet werden ~ 0 N E 31.5 ~ cu c:: '0;... 'E Cl ca Gi ~ 1.5 UJ y = -1,6471x R2 = 0,2369 p<0, Hochlage - Hanglage - Tallage Abb.19 Erster potentieller Schwärmtog (~ " ~ 20' Cj in den drei Hähenstufen des Nationalparks co <Xi co co co co co co co ~ ~ ~ ~ ~ ~ ~ ~ ~ N (") '<t U') CD... co '" co ~ 0 c;:; N (") '<t U') CD... co '" '" '" '" '" '" '" '" ~~ ~ N ~ ~ ~ ~ ~ ~ ~ ~ ~ '" 0 24



24 Damals wurde die Schwellentemperatur erst am 1. Juni erreicht. Die Spannweite des potenziellen Schwärmbeginns beträgt in den Tallagen 61 Tage. Im Gegensatz zu den anderen Höhenstufen scheinen die ersten potenziellen Schwärmtage zunehmend später aufzutreten. Diese Tendenz ist allerdings nicht signifikant. können die Beobachtungen aus Waldhäuser, nämlich wenige Schwärmtage Ende der 80er Jahre und die Zunahme der Schwärmtage in den 90er Jahren, auch für die Tallagen bestätigt werden. Die geringste Anzahl von Tagen größer 20 C konnte auch hier 1991 beobachtet werden. Der Maximalwert wurde 1993 mit 21 Tagen erreicht. Abb. 20, 21 u. 22 Anzahl der potenziellen Schwärmtage (~.. >20 C) in den Monaten April (blau) und Mai (rot) in den drei Höhenstufen des Nationalparks. Auch die Anzahl der Schwärmtage in den Monaten April und Mai ist eine wichtige Größe für die Entwicklung der Käferpopulation. Am ungünstigsten sind die Bedingungen in den Hochlagen; dort stehen in diesem Zeitraum im Durchschnitt nicht einmal vier Schwärmtage zur Verfügung. Im Bereich der Hanglagen sind es im Mittel 14 und in den Tallagen immerhin 20 Tage. Vor 1990 konnten Käfer in den Hochlagen, abgesehen von jeweils einem Maitag in den Jahren 1995 und 1996, erst im Juni schwärmen. Seit Mitte der 90er Jahre hat sich dort die Situation für die Buchdrucker deutlich verbessert: es treten häufiger Schwärmtage im Mai auf, im Jahr 2000 erstmals sogar im April. Die Zunahme der Schwärmtage in der Periode wa r sig n ifi ka nt. Bei der Analyse der Schwärmtage in den Hanglagen fallen zwei markante Muster auf: Während die 20 C-Schwelle zwischen 1987 und 1991 nur an zwei bis drei Tagen erreicht wurde, stieg die Anzahl der potenziellen Schwärmtage in den Folgejahren stark an und erreichte, abgesehen von 1994 mit nur einem potenziellen Schwärmtag, immer Werte über sieben. Im Jahr 2000 wurde mit 18 potenziellen Schwärmtagen das bisherige Maximum erreicht. Auch für die Hanglagen ergab sich eine signifikante Zunahme der Schwärmtage (Periode ). Für die Tallagen lässt sich aus den Werten der Wetterstation Klingenbrunn-Bahnhof keine gerichtete Entwicklung der Anzahl der potenziellen Schwärmtage erkennen. Nur andeutungsweise 25 - ~ 0 N 1\1 20 ~ E l- +"' 15 'E I:: QJ Cl cu I- I:: 0 > ::E cu 5 N I:: <C ' Y = O,1321x - 0,4771 R2 = 0,4424 p<o,01 ("') o:t Li) c.o r-. (Xl O'l 0 N ("') o:t (Xl (Xl (Xl (Xl (Xl (Xl (Xl O'l c; O'l O'l O'l ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ 25 Y = O,2734x + ',2094 ~ 0 R2 = 0,2633 p<o, ~ -_._~---" c;:: 20..._... ~ E l- +"' 15 'E I:: QJ $10 I:: 0 > ::E 5 cu N I:: <C 0 25 ~ 0 N 20 1\ ~ I-.<;: 15 E I:: QJ Cl cu I- 10 I:: 0 > ::E cu N 5 I:: <C 0 0 N o:t c.o (Xl 0 N (Xl (Xl (Xl (Xl (Xl O'l O'l ~ ~ ~ ~ ~ ~ ~ y = O,2377x + 7,4026 R2 = 0,0794 p>o,05 0 N o:t c.o (Xl 0 N (Xl (Xl (Xl (Xl (Xl O'l O'l ~ ~ ~ ~ ~ ~ ~ Hochlage Li) c.o r-. (Xl O'l 0 O'l O'l O'l O'l O'l 0 0 ~ ~ ~ ~ ~ N Hanglage _.,...,." '--.'._ -- I o:t c.o (Xl 0 O'l O'l O'l 0 ~ ~ ~ N o:t c.o (Xl 0 O'l O'l O'l 0 ~ ~ ~ N 25
25 Beim Vergleich des potenziellen Schwärmbeginns mit dem Zeitpunkt des letzten Tages mit Schneebedeckung wird deutlich, dass Temperaturen über 20 C, abgesehen von drei Ausnahmen, erst dann erreicht werden, wenn die Schneedecke bereits abgeschmolzen ist. Deshalb ist es nicht notwendig, den letzten Tag der Schneebedeckung als Kennwert für die Schwärmmöglichkeiten der im Boden überwinternden Buchdrucker zu verwenden. Der erste Tag mit Temperaturen über 20 C ist also in den allermeisten Fällen geeignet, den Schwärmbeginn sowohl der im Baum als auch der im Boden überwinternden Käfer zu charakterisieren. Zwischen potenziellen Schwärmtagen und dem Neuzugang an Totholzflächen scheint ein enger Zusammenhang zu bestehen. In Jahren, in denen der Buchdrucker erst spät schwärmt und die Anzahl der potenziellen Schwärmtage gering ist, wie dies Anfang der 80er Jahre der Fall war, ist auch der Buchdruckerbefall deutlich geringer. 1991, dem Jahr mit den ungünstigsten Schwärmbedingungen, wurde der erste potenzielle Schwärmtag in den Tallagen am 1. Juni, in den Hanglagen am 22. Juni und in den Hochlagen erst am 7. Juli registriert. Tatsächlich wurde im Jahr 1991 auch der geringste Zugang an Totholzflächen seit Beginn der Luftbildauswertung festgestellt. Seit 1991 erfolgte der Schwärmbeginn in den Hochlagen von Jahr zu Jahr früher, seit 1996 ergeben sich regelmäßig potenzielle Schwärmtage im Mai. Ein Befallszuwachs ist jedoch nur für die Jahre 1992, 1993, 1996 und 1997 festzustellen. Der markante Befallsrückgang in den Jahren 1994, 1997 und 1999 kann mit den potenziellen Schwärmbedingungen in den Hochlagen nicht erklärt werden. In den Tal- und Hanglagen geht der Rückgang des Neubefalls in diesen Jahren jedoch mit einem späteren potenziellen Schwärmbeginn und weniger potenziellen Schwärmtagen einher Entwicklungsbedingungen für die Käfer Die Entwicklung der Temperatursummen in Stundengraden verlief in allen Höhenstufen synchron. Beim Vergleich der absoluten Werte wird deutlich, wie unterschiedlich die Entwicklungsbedingungen für den Buchdrucker in den verschiedenen Höhenstufen des Nationalparks sind. In den Hochlagen wird im Mittel nur eine Temperatursumme von Stundengraden erreicht. In den Tallagen sind die Entwicklungsbedingungen für Buchdrucker mit Stundengraden wesentlich günstiger. In den Hanglagen erreichen sie mit Stundengraden mehr als das Doppelte der Hochlagenwerte. Die Temperatursumme von Stundengraden, die für die Jugendentwicklung des Buchdruckers notwendig ist, wurde in dichten Beständen auf Höhe des Arbergipfels selbst in günstigen Jahren nicht erreicht. Auch in dicht geschlossenen Tallagenbeständen wurden diese Temperatursummen in fast der Hälfte der Jahre nicht erreicht. Nur in den Hanglagenbeständen ist die Situation günstiger. Dort kann der Entwicklungszyklus selbst in dichten Beständen in fast jedem Jahr abgeschlossen werden. In allen Höhenzonen steigen die Temperatursummen seit Beginn der Wetteraufzeichnungen an. Allerdings kann diese Tendenz nur für die Hanglagen statistisch abgesichert werden. Auch zwischen Temperatursummen und Befallsentwicklung scheint es einen engen Zusammenhang zu geben, jedoch mit einer Zeitverzögerung von einem Jahr. Das heißt, dass hohe Temperatursummen im Jahr X erst einen Einfluss auf die Befallsentwicklung im Folgejahr haben. Die erreichte Temperatursumme bestimmt, ob die Käfer eine zweite Generation anlegen können und in 26



26 welchem Entwicklungsstadium sie überwintern. Kann sich die zweite Generation bis zum Jungkäfer entwickeln, ist die Wintersterblichkeit gering. Dann können im nächsten Frühling auch viele Tiere schwärmen und starken Befall verursachen. Müssen die Buchdrucker im Larvenstadium überwintern, ist die Wintermortalität wesentlich höher: die Anzahl der im nächsten Frühling schwärmenden Käfer ist geringer. Ebenso wie die Schwärmbedingungen waren auch die Entwicklungsbedingungen für Buchdrucker Ende der 80er und Anfang der 90er Jahre vergleichsweise schlecht. Darauf deuten die unterdurchschnittlichen Temperatursummen in diesem Zeitraum hin wurden in allen drei Höhenlagen sehr hohe Temperatursummen gemessen, denen ein starker Anstieg des Buchdruckerbefalls im Jahr 1993 folgte. Die unterdurchschnittliche Temperatursumme 1993 führte wiederum zu einem Rückgang des Totholzflächenzugangs im Jahr Im "Jahrhundertsommer" 1994 wurde in allen Höhenstufen die höchste Temperatursumme seit Beginn der Wetteraufzeichnungen im Nationalpark erreicht. Auf Grund der hohen Werte ist davon auszugehen, dass sich zumindest in den Hanglagen zwei Generationen ausbilden konnten. Folge war ein sprunghafter Anstieg des Befalls von 63 ha 1994 auf 367 ha Obwohl im Jahr 1995 die außergewöhnlichen Werte von 1994 nicht mehr erreicht wurden, lag die Temperatursumme weit über dem Durchschnitt der untersuchten Periode. Der Befall durch den Buchdrucker konnte sich noch einmal mehr als verdoppeln blieb die Temperatursumme wieder unter dem Durchschnitt; sie erreichte sogar nur ein Niveau wie Ende der 80er Jahre. Darauf folgte ein starker Befallsrückgang im Jahr Im Jahr 1998 bewegte sich der Neuzugang von Totholzflächen etwa in der Größenordnung von 1997, obwohl die Temperatursumme dieses Jahres gegenüber 1996 deutlich angestiegen war. Erwähnenswert ist, dass auf der Fläche des ehemaligen Forstamtes Zwiesel (überwiegender Teil des Falkenstein-Rachel-Gebietes), in dem sich der "Käferholzeinschlag" seit 1991 synchron zur Be "r ~ " ~ Hochlage - Hanglage - Tallage r:::: CIJ ~ ,---- C'l r:::: CIJ "C r:::: ~ "..,...f-----~~, I-~_,, --.At_~~-~--.~--I,.:: 2000 " 0 0 co N M.q- Ln (J:) r-. co (J) 0 ;;; co co co co co co co co co (J) ;!? ;!? ;!? ;!? ;!? ;!? ;!? ;!? ;!? ;!? ;!? ;!? N M.q- Ln (J:) r-. co (J) 0 (J) (J) (J) (J) (J) (J) (J) (J) ;!? ;!? ;!? ;!? ;!? ;!? ;!? ;!? 0 N Abb.23 Temperatursumme in Stundengraden bei einem Entwicklungsnullpunkt von 11,8 C (COELN 1997). 27



27 fallsfläche im Rachel-Lusen-Gebiet entwickelte, der Befall anstieg. Im Jahr 1999 ging der Buchdruckerbefall in beiden Teilgebieten zurück, obwohl die Temperatursumme des Jahres 1998 auf dem Niveau des Jahres 1997 geblieben war. Im Jahr 2000 entwickelte sich der Buchdruckerbefall in beiden Teilgebieten wieder auseinander: im Rachel-Lusen-Gebiet stieg er an, im Falkenstein Rachel-Gebiet ging er zurück. Auch in den be- nachbarten Forstämtern Bodenmais und Freyung ging der "Käferholzanfall" 2000 zurück. Aufgrund der geringen Temperatursumme im Jahr 1999 wäre ein leichter Rückgang zu erwarten gewesen. Über den Betrachtungszeitraum besteht ein signifikanter Zusammenhang zwischen der Temperatursumme des Vorjahres und derveränderungsrate der Totholzfläche gegenüber dem Vorjahr (R' =0,59, p<o,ol). 5. Diskussion Abb. 24 Jüngere Fichtenbestände werden selbst bei hohen Populationsdichten des Buchdruckers nur im Übergangsbereich zu Altbeständen befallen. Die Entwicklungsdynamik von Insektenpopulationen wird nach SCHWENKE (1981) im Wesentlichen durch folgende fünf Regulationsmechanismen bestimmt: 1. Habitatangebot 2. Antagonisten 3. endogene Faktoren 4. Konkurrenz 5. Witterungsverlauf In den nachfolgenden Kapiteln wird die Bedeutung der einzelnen Regulationsmechanismen und abschließend ihr Zusammenspiel während der Buchdruckermassenvermehrung im Nationalpark Bayerischer Wald diskutiert. 28

.")
28 5.1 Habitatangebot Der Buchdrucker ist in seinem gesamten Lebenszyklus eng an die Baumart Fichte gebunden. Abgesehen vom Schwärmflug vollzieht sich seine Entwicklung auf der Stammoberfläche und im Bast von älteren Fichten, denn nur dort können die Käfer erfolgreich brüten. Wesentlicher Habitatfaktor für den Buchdrucker ist somit die Verfügbarkeit von Fichten mit einem Brusthöhendurchmesser von mindestens 20 cm (PRELL 1948). Bevorzugt werden allerdings stärkere Fichten in einem Alter zwischen 80 und 100 Jahren befallen (ESCHERICH 1923). Aufgrund ihrer Erfahrungen in Baden-Wü rttemberg ischen Naturwa Idreservaten differenzieren BECKER und SCHROETER (2000) die Habitatqualität von Fichtenwälder genauer: "Durch den Buchdrucker wurden Bestände älter als 50 Jahre befallen. Bis zu 70 Jahre alte Bestände waren nur bei sehr hoher Populationsdichte und mangelnden Alternativen gefährdet. Bestände im Alter zwischen 70 und 90 Jahren wiesen bereits ein deutlich höheres Befallsrisiko auf. Ab einem Bestandesalter von 100 Jahren nahm das Befallsrisiko erheblich zu." Während Buchdruckermassenvermehrungen kommt es zwar vor, dass auch jüngere Bäume bis Daumenstärke angeflogen werden. Unter deren Rinde können allerdings keine Brutsysteme angelegt, sondern bestenfalls Regenerationsfraß durchgeführt werden (PRELL 1948, HEURICH 2001). Bestandes- Nadelbaumanteil (Ofo) alter bis 20 I J unter ungefährdet 30 Jahre über 30 I Bedingt I Potentiell Jahre gefährdet gefährdet 1999 führten NUESSLEIN et al. eine Gefährdungsanalyse für die Wälder des Nationalparks durch. Die zugrundeliegende Klassifizierung der Bestände ist in Tabelle 3 angegeben. Die Ergebnisse dieser Studie sind zwar keine Habitatkartierung im eigentlichen Sinn, da sie die unterschiedlichen Präferenzen der Käfer für verschiedene Altersklassen nicht berücksichtigen, sie geben aber einen guten Überblick über Umfang und Verteilung befallsgefährdeter Flächen. Gemäß der oben beschriebenen Kriterien wurden 62 % der Bestände im alten Nationalparkgebiet als gefährdet und 10 % als bedingt gefährdet eingestuft. Allerdings sind diese Flächen nicht gleichmäßig verteilt, die Unterschiede zwischen den Höhenstufen sind beträchtlich. Besonders in den Hoch- und Tallagen dominieren die gefährdeten bzw. bedingt gefährdeten Bestände mit 93 bzw. 90 %.In den Hanglagen ist ihr Anteil weitaus geringer. In der Unteren Hanglage sind 69 % und in der Oberen Hanglage 63 % der jeweiligen Fläche als gefährdet bzw. bedingt gefährdet eingestuft. Bei einem genaueren Vergleich der Wälder in den Hoch- und Tallagen, der auf ei~er Klassifikation nach Waldentwicklungsstadien (für eine Erläuterung der Entwicklungsstadien siehe HEURICH 2001) basiert, lässt sich allerdings feststellen, dass die Bestände in den Tallagen einen aus der Sicht des Buchdruckers wesentlich ungünstigeren Jugendstadium Wachstumsstadium Reifungsstadium D Verjüngungsstadium Plenterstadium Abb.25 Anteile der verschiedenen Waldentwicklungsstadien in den Hoch- und Tallagen (Stand 1992). Tab. 3 Kriterien zur Ausscheidung von Gefährdungsklassen (nach NUESSLEIN et al. 1999). 29




29 Abb. 26 u. 27 Kleinere Windwürfe (u.) und durch Blitzschlag geschädigte Bäume sind in den Wäldern des Nationalparks allgegenwärtig und bieten dem Buchdrucker günstigen Brutraum. Altersaufbau besitzen. Jugendstadien, Wachstumsstadien und sonstige Flächen, die nicht oder nur bei sehr hohen Populationsdichten vom Buchdrucker befallen werden können, erstrecken sich nach dieser Klassifikation auf über 50 % dertailagen. In den Hochlagen beträgt der Anteil der entsprechenden Flächen lediglich 17 Ofo. Dies ist ein wesentlicher Grund, warum die Befallsentwicklung in den Tallagen - trotz größerer Windwurfhäufigkeit - weit langsamer abläuft, als in den Hochlagen. Daneben spielen auch noch die weniger kompakte Form und der hohe Anteil des Randbereichs eine Rolle. Damit standen den Buchdruckern besonders in den Hochlagen große, zusammenhängende Flächen geeigneten Habitats zur Verfügung. In den Hang- und Tallagen waren die Verhältnisse etwas ungünstiger, allerdings gab und gibt es auch hier lokal größere Flächen mit geeigneten Fichtenaltbeständen. Während die hohen Fichtenanteile in den Tal- und Hochlagen vor allem durch natürliche Standortsfaktoren bedingt sind, haben sich die Habitatbedingungen für den Buchdrucker seit Beginn der geregelten Forstwirtschaft gegen Mitte des 19. Jahrhunderts in den Hanglagen merklich verbessert. Während die Bestände damals etwa zu je einem Drittel aus Fichte, Buche und Tanne zusammengesetzt waren (PLOCHMANN 1961). verdoppelte sich der Anteil der Fichte aufgrund der Bewirtschaftungsverfahren bis 1991, der Anteil der Tanne ging entsprechend zurück (RALL 1995). In der Latenzphase (Zustand zwischen den Massenvermehrungen) treten Buchdrucker nur in sehr geringen Dichten auf. Mit Hilfe effektiver Abwehrmechanismen, zu denen auch das Ausharzen sich einbohrender Käfer gehört, können die Bäume angreifende Käfer abwehren. Die Käfer sind dann nicht in der Lage, vitale Fichten zu besiedeln. In dieser Phase sind die Buchdrucker folglich auf die Nutzung geschwächter Fichten angewiesen. Beispiele hierfür sind einzelne Fichten mit Wipfelbrüchen, vom Blitz getroffene Bäume oder vom Wind geworfene Individuen. Umfangreicher Schneebruch, Windwurf oder ungewöhnliche Trockenheit führen zu einem größeren Angebot geschwächter Bäume, die nicht mehr in der Lage sind, die Käfer effektiv abzuwehren. Diese Bäume werden daher umgehend von den Käfern besiedelt. Aufgrund der günstigen Brutbedingungen kommt es in Kombination mit dem hohen Repro- 30
 während des anschließenden Schwärmfluges kann es den Käfern gelingen, auf einzelnen Bäumen sehr hohe Individuendichten zu erreichen.")

30 duktionspotential zu einem starken Anwachsen der Populationsdichte (Progradation). Durch den Einsatz von Lockstoffen (Aggregationspheromone) während des anschließenden Schwärmfluges kann es den Käfern gelingen, auf einzelnen Bäumen sehr hohe Individuendichten zu erreichen. Durch den dann möglichen massenhaften Angriff auf den Baum bricht dessen Abwehrssystem zusammen; selbst vitale Bäume können dann besiedelt werden. Über die Frage, welche Faktoren die Buchdruckermassenvermehrung seit 1993 ausgelöst haben, wird bis heute kontrovers diskutiert. Im Gegensatz dazu war die Situation 1983/84, als Gewitterstürme insgesamt 173 ha Wald zu Boden rissen, eindeutig zu erklären "Wiebke" fm fm fm fm fm zeinen Windwürfen vor allem auf Nassböden, die über das ganze Nationalparkgebiet verteilt waren. Diejenigen Stürme, die größere Baumbestände zu Boden warfen, sind in Tabelle 4 aufgeführt. Allerdings handelt es sich bei diesen Angaben nur um grobe Schätzwerte. Ein genauer Überblick über das Ausmaß der Windwürfe existiert nicht. Als Anhalt für den Umfang der Windwürfe im Rachel-Lusen-Gebiet können aufgrund der ähnlichen Topographie die Holzmengen aufgearbeiteter Windwürfe im ehemaligen Forstamt Zwiesel (große Teile des Falkenstein-Rachel-Gebietes) herangezogen werden. Zwischen 1990 und 1996 wurden dort fm Windwurf aufgearbeitet. Dies entspricht ungefähr dem Holzanfall bei den Windwürfen der Jahre 1983 und Da die Windwürfe in der Naturzone des Nationalparks nicht aufgearbeitet wurden, stand dem Buchdrucker somit wahrscheinlich ein großes Angebot an Brutraum zurverfügung. Tab. 4 Bedeutende Windwürfe im Rachel-Lusen -Gebiet zwischen 1990 und 1995 (nach JEHL 1997). Abb. 28 Größere Windwürfe, wie hier in der Abteilung Rachelkapelle, traten in den 90er Jahren nur selten au f. Windwürfe dürften allerdings auch als Initial der aktuellen Gradationswelle von großer Bedeutung gewesen sein. So berichtet JEHL (1997), dass in der Periode zwischen 1990 und 1996 in der Wetterstation Waldhäuser insgesamt 52. Stürme mit einer Windstärke von über 8 Beaufort gemessen wurden. Bei fast all diesen Stürmen kam es zu ein- Zusätzlich wurde die Vitalität der Fichten 1992 vermutlich durch eine Trockenperiode im Sommer 1992 gemindert. BEUDERT et al.(1997) konnten aufgrund von Bodenwasserbilanzen für einen Fichtenbestand in den Tallagen nachweisen, dass der Transpirationsbedarf im Zeitraum Mai bis August über dem Bestandesniederschlag lag und selbst die vollständige Nutzung des gebundenen nutzbaren Wassers die negative Wasserbilanz nicht ausgleichen konnte. Ob diese Trockenperiode in den Hochlagen aufgrund der dort flachgründigeren Böden eine bedeutendere Rolle gespielt hat, ist schwierig zu beurteilen. Dagegen spricht, dass die niedrigeren Temperaturen in höheren Lagen auch eine geringere Verdunstung zur Folge haben und die geringe Wasserkapazität der 31

 vermuten, kann nicht belegt werden.")

31 Abb.29 Auf Dauerbeobachtungsflächen wird der Eintrag von Schadstoffen in die Waldökosysteme des Nationalparks gemessen. Böden zumindest teilweise durch die größeren Baumabstände ausgeglichen wird. Allerdings sind die "Hochlagenfichten" an eine gute Wasserversorgung "gewöhnt" und reagieren auf Grund ihrer im Verhältnis zur großen transpirierenden Oberfläche geringen Wasserabsorptionskapazität der Wurzel empfindlicher auf Wassermangel (KAUF MANN zit. in KAHLE 1994). Ob es zusätzlich bereits Anfang der 90er Jahre aufgrund schneearmer Winter zu Engpässen in der Wasserversorgung kam, wie SCHOPF et al. (1993) vermuten, kann nicht belegt werden. Darüber hinaus traten 1992 und 1995 Samenjahre bei der Fichte auf. Folge war, dass Reservestoffe und Assimilate der Bäume für die Bildung von Samen und Zapfen verbraucht wurden, was eine große Belastung für die Bäume bedeutet. Anschaulich wird diese Anstrengung, wenn man bedenkt, dass im Jahr 1992 für die Anlage der Zapfen und Samen in etwa so viel Energie aufgewendet wurde, wie für den jährlichen Holzzuwachs. Auch die deutliche Reduktion der mittleren Länge neu gebildeter Seitentriebe auf 30 0/0 (1992) bzw. 60 % (1996) zeigt deutlich, wie stark die Bäume durch eine Vollmast in Anspruch genommen werden (BEuDERTet al. 1997). Eine andere Argumentationskette stellt die Bedeutung "neuartiger Waldschäden" für die Entstehung der Massenvermehrung heraus. Es wird vermutet, dass die durch den Menschen verursachte Schadstoffbelastung zur Vitalitätsminderung der Fichten führt und die geschwächten Bäume leichter vom Buchdrucker befallen werden können. Von besonderer Bedeutung ist in diesem Zusammenhang der Eintrag von Schwefel- und Stickstoffverbindungen, der zu einer Versauerung und damit einem Anstieg von Protonen- und Aluminiumkonzentrationen im Boden führt, die wiederum schädigend auf die Feinwurzeln der Bäume wirken können. Darüber hinaus hat die Versauerung auch die Auswaschung bedeutsamer Pflanzennährstoffe wie Kalzium, Magnesium und Kalium zur Folge, die den Bäumen anschließend nicht mehr zurverfügung stehen. Hohe Stickstoffeinträge wirken wachstumsfördernd und bewirken dadurch einen höheren Verbrauch von Magnesium und Kalium; der besonders auf den sauren Böden nicht mehr gedeckt werden kann. Darüber hinaus erhöhen die Stickstoffeinträge die Anfälligkeit gegenüber Trockenheit und Frost und führen zu Wuchsungleichgewichten zwischen Spross und Wurzel. Auch die in den letzten Jahren gemessenen hohen Ozon konzentrationen könnten eine Rolle bei den Absterbeprozessen gespielt haben, da durch sie die Funktion der Spaltöffnungen beeinträchtigt und die Frostresistenz herabgesetzt wird. Die im Nationalparkgebiet gemessene Ozon belastung liegt in fast allen Jahren über dem definierten Belastungsindex (AOT). Allerdings ist die ökologische Relevanz dieses Indexes noch unklar, zumal bei Begasungsversuchen im Freiland (Freeair) selbst bei dreifacher Überschreitung dieses 32


32 Damit liegen die Werte im Durchschnitt der bayerischen Waldklimastationen (BAYERISCHE LANDESANSTALT FUER WALD UND FORSTWIRTSCHAFT 2000, BEUDERT et al. 1997, GIETL 1990). Abb.30 Jahrringanalysen geben wichtige Hinweise über die Veränderung der Vitalität von Waldbäumen. Die Einträge von Stickstoffverbindungen verminderten sich seit Mitte der 80er nur in geringem Maße. Unter Buche gingen sie von etwa 11 auf 8 bis 9 kg ha a und im Freiland von 13 bis 14 auf 9 bis 10 kg ha a zurück. Damit liegt die Belastung durch Stickstoffeinträge erheblich unter dem Durchschnitt aller Bayerischen Waldklimastationen (BAYERISCHE LANDESANSTALT FUER WALD UND FORSTWIRTSCHAFT 2000, BEUDERT et al. 1997, GIETL 1990). Wertes keine Schadenssymptome hervorgerufen werden konnten (BEUDERT et al. 1997, SKEFFINGTON und Mc LEOD 1996). Die Summe der Einträge an Ammonium (NH.J, Nitrat (NO) und Sulfat (S0.J, die ein gutes Maß für die Gesamtbelastung der Wälder mit versauernden Stoffen ist, ging im Nationalpark seit 1991 um 24 (Freiland) bis 49 % (Buche) zurück. Sie lag 1999 in einem Bereich zwischen 0,9 (Buche) und 1,0 (Freiland) kmolc ha ' a ' (BEUDERT et al. 1997), was im Vergleich mit den bayerischen Waldklimastationen einen der günstigsten Werte darstellt (BAYERISCHE LANDESANSTALT FUER WALD UND FORSTWIRTSCHAFT 2000). Allerdings wird durch diese Säureeinträge das Puffervermögen der Böden, das in einer Größenordnung von 1, in den Hochlagen eher bei 0,5 kmolc ha ' a ' liegt, noch überschritten. Die Einträge von Schwefelverbindungen sind seit Mitte der 80er Jahre, als die ersten Messungen durchgeführt wurden, stark zurückgegangen. Unter Buche verminderten sich die Einträge von 17 bis 21 auf etwa 5 bis 7 kg ha ' a ', im Freiland von 10 bis 13 auf heute ebenfalls etwa 5 bis 7 kg ha ' a '. Bei den Depositionen besteht jedoch ein ausgeprägter Höhengradient, die Belastung in den Hochlagen liegt erheblich über den aufgeführten Werten (BEUDERT et al. 1997). Die Belastungen des Waldökosystems im Nationalpark haben sich damit seit den 80er Jahren, vor allem durch die Halbierung bzw. Drittelung der Einträge von Schwefelverbindungen, aber auch durch einen Rückgang der Stickstoffeinträge deutlich verringert und zu einer Entspannung der Situation beigetragen (GIETL 1990, BEUDERT und BREIT 2001). Dass mittlerweile auch die Bäume auf die positive Entwicklung reagieren, zeigt die Entwicklung verschiedenervitalitätsindikatoren: So nimmt die Breite der Jahrringe bei der Fichte wieder zu. PICARD et al. (1999) konnten auf Grund von Jahrringanalysen im Nationalpark nachweisen, dass sich die Fichten nach einer Wuchsdepression in den 70er Jahren wieder erholten. Allerdings war die Regeneration nicht bei allen untersuchten Individuen gleichermaßen ausgeprägt. Manche Probebäume konnten sich nicht erholen, bei ihnen verharrte der Jahrringzuwachs auf niedrigem Niveau. 33


33 Abb.37 Vergilbungserscheinungen infolge von Nährstoffmangel, wie hier bei diesen Fichten, sind seit 7985 erheblich zurückgegangen. Blattschäden in der Lichtkrone von Buchen beobachten. Diese waren entgegen der Vermutung in den höher gelegenen Beständen weniger ausgeprägt als in den tiefer gelegenen. An Fichten konnten bisher keine Schäden nachgewiesen werden. Auch die auf Magnesiummangel zurückgehenden Vergilbungserscheinungen bei der Fichte haben sich seit 1985 deutlich vermindert. Während 1985 noch % der Fichten im Wuchsgebiet Bayerischer Wald Nadelvergilbungen aufwiesen, verringerte sich ihr Anteil bis 1997 auf /0 (BAYERISCHE LANDESANSTALT FÜR WALD UND FORSTWIRTSCHAFT 2000). Die Ergebnisse der Kronenzustandserhebung aus CIR-Luftbildern im Nationalpark liefern ein etwas abweichendes Bild: Sowohl der mittlere Nadelverlust als auch der Anteil von Individuen mit deutlichen Schäden stieg in den 90er Jahren langsam an bzw. bewegte sich auf einem hohen Niveau. Auch der Waldschadensbericht 2000 bestätigt diese Entwicklung. Bei der Differenzierung nach Höhenlagen zeigt sich, dass sowohl der Anteil von Individuen mit deutlichen Schäden als auch der mittlere Nadelverlust in den Hochlagen Anfang der 90er Jahre deutlich geringer war als in den Hanglagen (BEUDERT und BREIT 2000, BAYERI SCHE LANDESANSTALT FÜR WALD UND FORSTWIRT SCHAFT 2000). Welche Bedeutung den neuartigen Waldschäden bei der Auslösung des Befallsgeschehens im Nationalpark Bayerischer Wald zukommt, ist umstritten. Allerdings scheint es unwahrscheinlich, dass neuartige Waldschäden gerade dann starken Buchdruckerbefall ausgelöst haben sollen, als die Schadstoffbelastung des Ökosystems bereits kräftig zurückgegangen war und die Bäume deutliche Erholungstendenzen zeigten. Viel plausibler wäre es gewesen, wenn der Käferbefall bereits in den 80er Jahren - während der höchsten Schadstoffbelastung - aufgetreten wäre. Obwohl die thermischen Bedingungen z. B. in den Jahren 1983 und 1984 relativ günstig waren, wurde damals keine größere "spontane" Käferentwicklung beobachtet. Die Befallsentwicklung nach 1984 konnte auf die Windwurfereignisse 1983 und 1984 zurückgeführt werden. Auch die von PICARD et al. (1999) aufgestellte Vermutung, dass die Altbäume, die sich erwiesenermaßen nicht von den starken Schädigungen in den 70er Jahren erholen konnten, bevorzugt vom Buchdrucker befallen wurden, ist eher unwahrscheinlich. Zum einen ist der Stichprobenumfang, auf den sich PICARD et al. (1999) stützen, mit nur vier Probeflächen und jeweils 20 beprobten Bäumen sehr gering, zum anderen zeigen Untersuchungen von SCHOPF und KOEHLER (1995), dass Fichten mit hohen Nadelverlusten nicht bevorzugt von Buchdruckern befallen wurden. Blattschäden in Folge einer hohen Ozonbelastung wurden im Nationalpark intensiv an der im Vergleich zur Fichte empfindlicher reagierenden Buche untersucht konnte BAUMGARTEN (1999) Damit kann der Verlauf des Buchdruckerbefalls die "Waldschadenshypothese" nicht stützen, da die Käfer dafür die "falschen" Bäume zur "falschen" Zeit befallen haben. 34
 nennt 55 Arten. Zu den wichtigsten Buchdruckerantagonisten zählen: Spechte (Picidae spec.), Waldameisen (Formica spec.), Laufkäfer (Carabidae spec.), Raubfliegen (Asilidae spec.")
, Kurzflügler (Staphylinidae spec.), Glanzkäfer (Nitidulidae spec.), Fadenwürmer (Nemathelminthesspec.), Pilze, Bakterien u. a. (SCHOPF und KOEHLER 1995, SCHWENKE 1974, NIERHAUS-WUNDERWALD 1993 zit.")
34 5.2 Antagonisten Es gibt eine ganze Reihe von Gegenspielern (Antagonisten) des Buchdruckers. So listen SAMUELSSON et al. (1994, zit. in SCHERZlNGER 1996) 140 Insektenarten auf, die sich u. a.. von Borkenkäfern ernähren. Auch OKOLOW (1991) nennt 55 Arten. Zu den wichtigsten Buchdruckerantagonisten zählen: Spechte (Picidae spec.), Waldameisen (Formica spec.), Laufkäfer (Carabidae spec.), Raubfliegen (Asilidae spec.), Kamelhalsfliegen (Raphidia spec.), Buntkäfer (C/eridae spec.), Schlupfwespen (Apocrita spec.), Brackwespen (Braconidae spec.), Erzwespen (Chalcidiodea spec.), Rindenkäfer (Rhizophagidae spec.), Kurzflügler (Staphylinidae spec.), Glanzkäfer (Nitidulidae spec.), Fadenwürmer (Nemathelminthesspec.), Pilze, Bakterien u. a. (SCHOPF und KOEHLER 1995, SCHWENKE 1974, NIERHAUS-WUNDERWALD 1993 zit. in SCHERZlNGER 1996). nicht im Stande gewesen waren, der Kalamität bald Einhalt zu tun" (BUETTNER 1927). Allerdings konnte HERRLEIN (1878) damals beobachten, wie parallel zum Anstieg der Population des Käfers auch dessen Feinde zunahmen. Ab 1875 trat verstärkt eine "Pilzbildung an den Borkenkäferlarven" auf. "Mehr als die Hälfte der Borkenkäferlarven" war "schwarz verfärbt und tot". Ob die Antagonisten damals eine Bedeutung für den Rückgang des Käferbefalles hatten, lässt sich aus den Aufzeichnungen von HERRLEIN jedoch nicht ableiten. Abb.32 Schlupfwespen legen ihre Eier mit einem Legebohrer in die Larven der Buchdrucker, in denen sie anschließend parasitisch leben. Auf dem Bild sieht man die Kokons von verpuppten Schlupfwespenlarven. Trotz dieser hohen Artenanzahl scheinen die Buchdruckergegenspieler nur einen geringen Einfluss auf den Massenwechsel der Käfer zu haben. Wenn sich der r-stratege Buchdrucker auf Grund günstiger Umweltbedingungen massenhaft vermehren ~ann, ist seine Population in der Lage, auch sehr hohe Verluste durch Prädation und Parasitierung auszugleichen (SCHERZlNGER 1996). Auch SKATULLA vertritt die Meinung, dass natürliche Feinde i. d. R. die Populationsdynamik des Buchdruckers nicht beeinflussen. Ihm ist kein Beispiel bekannt, in dem eine Buchdruckerkalamität auf Grund natürlicher Feinde zusammengebrochen ist. WERMELINGER vermutet, dass natürliche Feinde lediglich einzelne "Käfernester" auslöschen könnten (beide in: BAYERISCHE LANDESAN STALT FUER WALD UND FORSTWIRTSCHAFT, 1998). Auch die Käferkalamität im Anschluss an die Windwürfe 1868/70 wurde wahrscheinlich nicht durch natürliche Feinde gestoppt. Damals kam man während einer gemeinsamen Tagung bayerischer, österreichischer und böhmischer Forstleute zum Schluss, dass "die Feinde des Borkenkäfers Für den Rückgang der Befallsentwicklung im Anschluss an die Windwurfereignisse 1983/84 konnte ebenfalls ke in Zusammenhang mit der Ent wicklung der Antagonisten festgestellt werden, "nur loka l könnte Coeloides bostrychorum (eine Erzwespenart) von Bedeutung gewesen sein" (SCHOPF und KOEHLER 1995). In den letzten Jahren gab es zwar Hinweise auf eine z. T. starke Verpilzung der Larven und eine Zunahme der Parasiten (WEISSBACHER pers. Mitteilung), ein bedeutender Einfluss auf die Dichte der Käferpopu lation konnte bislang jedoch nicht festgestellt werden. Allerdings ist zu bedenken, dass es in Mitteleuropa bisher nicht möglich war, den ungestörten Ablauf einer Massenvermehrung des Buchdruckers und die Entwicklung seiner Antagonisten auf großer Fläche zu beobachten. Auf Grund dieser Wissenslücken ist es trotz methodischer Schwierig keiten wichtig, den Einfluss der Antagonisten auf die Populat ionsentwicklung und ihre tatsächliche Bedeutung im Massenwechsel weiterhin zu verfolgen. 35
35 5.3 Endogene Faktoren Eine wesentliche Voraussetzung, Fichtenbestände großflächig zum Absterben bringen zu können, ist das bereits beschriebene enorme Vermehrungspotential des Buchdruckers. Die Frage, ob und in welchem Umfang dichteabhängige Faktoren die Populationsdynamik des Buchdruckers beeinflussen und welche Mechanismen dabei zum Tragen kommen könnten, ist bisher nur unzureichend erforscht. Nach WEISSBACHER (1999) ändern sich im Verlauf des Massenwechsels die Populationskennwerte GeschlechteNerhältnis und Eizahl je Muttergang: Während das GeschlechteNerhältnis in der Latenzphase (Zustand zwischen den Massenvermehrungen) ausgeglichen ist, verschiebt es sich in der Progradation (starkes Anwachsen der Population) und Gradation (Höhepunkt der Massenvermehrung) zugunsten der Weibchen und fällt während der Retrogradation (Rückgang der Populationsdichte) wieder ab. Die durchschnittliche Eizahl je Muttergang liegt in der Latenz bei 40 bis 50 und steigt während Progradation und Gradation auf 60 bis 80 an. In der Retrogradation liegen die Eizahlen bei 30 bis 40 oder sogar noch darunter. HERRLEIN (1878) beobachtete während der Buchdruckermassenvermehrung 1868 bis 1876 im Böhme ~wald ebenfalls die Entwicklung der Eizahlen je Weibchen. -Während 1872/73 jedes Weibchen zwischen 60 und 100 Eiern ablegte, waren es 1874 nur noch 40 bis 70 Eier reduzierte sich die Zahl nochmals auf 25 bis 30. Leider ist unbekannt, auf welcher Datenbasis diese Aussagen beruhen. Bei den beobachteten Veränderungen der Populationsparameter ist unklar ob sie innerhalb der Populationsdynamik des Buchdruckers Ursache oder Wirkung sind. Während der aktuellen Buchdruckermassenvermehrung konnten weder während der Feldstudien zwischen 1989 bis 1992 noch in den Feldstudien nach 1997 Indizien für die Bedeutung endogener Faktoren für die Populationsentwicklung gefunden werden. Beispielsweise blieb das Geschlechterverhältnis selbst während der Phase des Befallsrückganges nach 1989 zu Gunsten der Weibchen verschoben, auch die Eizahlen streuten stark, ohne dass eine Tendenz erkennbar war. (SCHOPF et al. 1993, WEISSBACHER 1998, 1999, 2000). Hätte es sich damals um eine klassische Retrogradation gehandelt, wäre ein ausgeglichenes GeschlechteNerhältnis und sinkende Eizahlen zu erwarten gewesen. Allerdings ist zu beachten, dass die Zahl der untersuchten Proben auf Grund des hohen Bearbeitungsaufwandes jeweils relativ gering war und deshalb Aussagen über die Veränderung von Populationsparametern sowohl auf lokaler als auch auf Ebene des Nationalparkgebiets (soweit dies überhaupt Sinn macht) nur von eingeschränkter Aussagekraft sind. Zusammenfassend kann man feststellen, dass endogene Faktoren nach dem aktuellen Kenntnisstand im bisherigen Verlauf der Massenvermehrung keine auffällige Rolle gespielt haben.



36 5.4 Konkurrenz Ein weiterer Faktor, dem ein Einfluss auf die Populationsentwicklung zugeschrieben wird, ist die Konkurrenz der Käfer um den knappen Brutraum. Wenn sich sehr viele Altkäfer in einen Stamm einbohren, liegen die Muttergänge sehr dicht zusammen. Den einzelnen Larven steht dann nur ein begrenztes Nahrungsangebot zur Verfügung: Sie müssen um die knappen Ressourcen konkurrieren bzw. fressen sich gegenseitig die Nahrung weg. In der Folge können sich die Larven nicht ausreichend entwickeln, erreichen nur ein geringes Gewicht oder sterben ganz ab. Schließlich schlüpfen aus den verbliebenen Puppen relativ kleine Käfer, die nur über geringe Nährstoffreserven verfügen. Somit sinkt der Reproduktionserfolg eines Weibchens mit zunehmender Besiedlungsdichte. Dieser Zusammenhang konnte in verschiedenen Studien (ANDERBRAND et al. 1985, BERRYMAN 1974, BEAVER 1974) u. a. auch im Nationalpark Bayerischer Wald bestätigt werden (SCHOPF und KOEHLER 1995). Abb.33 Wenn sehr viele Käfer ihre Brutsysteme in einem Baum anlegen, können sich die Larven aufgrund der sehr eng zusammenliegenden Muttergänge (senkrecht) nicht optimal entwickeln. Nach SCHWERDTFEGER (1974) soll diese Konkurrenz zwischen den Larven "oft ausschlaggebend am raschen Zusammenbruch von Käferkalamitäten beteiligt" sein. Weil die Buchdru-eker in ihrem begrenzten, zweidimensionalen Lebensraum unter der Fichtenrinde einem starken Konkurrenzdruck ausgesetzt sind, besteht ein starker Selektionsdruck zugunsten von Individuen, die Konku rrenz vermeiden. Die Annahme, dass der innerartlichen Konkurrenz eine große Bedeutung in der Regulation der Populationsgröße (-dichte) zukommt, steht somit im Gegensatz zu den "Interessen" der Käferindividuen und zu den ausgeklügelten, auf Konkurrenzvermeidung ausgelegten Kommunikationssystemen des Buchdruckers. Beispiele hierfür sind die Produktion von verschiedenen Ablenkpheromonen (lpsenol, Ipsdinol, Verbenon), die anfliegenden Käfern signalisieren, dass Bäume oder Rindenabschnitte bereits dicht besiedelt sind und die Anlage von Brutsystemen keinen Erfolg verspricht. Andere Verhaltensmechanismen tragen dazu bei, dass Mindestdistanzen zu benachbarten Einbohrlöchern eingehalten werden und dass sich sowohl die Muttergänge, als auch die Larvengänge nicht überschneiden (BYERS 1993). Auch die Beobachtungen im Nationalpark bestätigen diese Überlegungen: Obwohl nach dem Absterben großer Fichtenkomplexe starke Nahrungskonkurrenz beobachtet wurde - die Buchdrucker bohrten sich selbst in daumenstarke Fichten ein - hatte diese große innerartliche Konkurrenz keinen wesentlichen Einfluss auf den Fortgang der Massenvermehrung. 37

 Organismen, deren Aktivität und Entwicklungsdynamik maßgeblich von der Umgebungstemperatur beeinflusst wird.")
37 5.5 Witterungsverlauf Abb. 34 Zur Überwachung des Sch wärmfluges der Käfer setzt die Bayerische Landesanstalt für Wald und Forstwirtschaft im Nationalpark Mess Stationen ein, mit denen neben dem Käferanflug auch die Lufttemperatur und Luftfeuchtigkeit kontinuierlich aufgezeichnet werden. Buchdrucker sind wechselwarme (poikilotherme) Organismen, deren Aktivität und Entwicklungsdynamik maßgeblich von der Umgebungstemperatur beeinflusst wird. Insbesondere Schwärmbeginn, Entwicklungsgeschwindigkeit der Brut und damit die Anzahl der innerhalb eines Jahres auftretenden Generationen sind stark vom saisonalen Witterungsgeschehen abhängig. Die Rekonstruktion buchdru'ckerrelevanter Klima parameter für die Periode 1980 bis 2000 bestätigt die Bedeutung des Witterungsverlaufes für die Populationsdynamik des Buchdruckers. Fast alle Schwankungen im Befallsgeschehen können mit Hilfe der verwendeten Parameter erklärt werden. Insbesondere zwischen der Veränderung des Befalls gegenüber dem Vorjahr und der Temperatursumme des Vorjahres besteht ein signifikanter Zusammenhang. Durch Messung der Kambialtemperaturen seit 2000 wird die Prognose für die Käferentwicklung, aber auch die Erklärung der zurückliegenden Entwicklung weiter verbessert werden. Die Schwärmbedingungen haben vor allem dann eine große Bedeutung, wenn die Käfer erst sehr spät schwä-rmen können und nur wenige Schwärmtage zur Verfügung stehen, wie dies am Beispiel des Jahres 1991 deutlich wird. Außerdem verlängert sich durch den früheren Schwärmbeginn der Zeitraum, in dem sich die Brut entwickeln und eventuell eine weitere Generation angelegt werden kann. Bemerkenswert ist hier vor allem, dass der Schwärmbeginn in den Hochlagen im Vergleich zum Zeitraum Ende der 80er Jahre fast einen Monat früher erfolgt. Somit kann der Rückgang des Buchdruckerbefalls bis 1992 vor allem durch den ungünstigen Witterungsverlauf Ende der 80er und Anfang der 90er Jahre erklärt werden. Als sich die Witterungsbedingungen ab 1992 verbesserten, war der Weg frei für einen raschen Anstieg der Buchdruckerpopulation, der nur durch die thermisch ungünstigen Jahre 1993 und 1996 leicht abgebremst wurde. Schwierigkeiten bereitet allerdings die Interpretation der Befallsentwicklung der Jahre 1999 und Aufgrund der Temperatursummen in den Jahren 1998 und 1999 wäre im Jahr 1999 ein unveränderter und im Jahr 2000 ein leicht abnehmender Befall zu erwarten gewesen. Tatsächlich ging er im Jahr 1999 zurück und stieg im Jahr 2000 stark an. Grund dafür könnten die relativ schlechten Schwärmbedingungen im Jahr 1999 und die sehr guten Schwärmbedingungen im Jahr 2000 gewesen sein. Aber auch die Umstellung auf eine Herbstbefliegung kann hierzu einen Erklärungsbeitrag liefern. Die im Spätsommer 1999 befallenen Bäume wurden erst mit der Befliegung im Jahr 2000 erfasst und damit dem Jahr 2000 zugerechnet. Leider lässt sich der Anteil dieser Bäume nicht quantifizieren, so dass unklar bleibt, um we lchen Wert die Befallsfläche des Jahres 1999 erhöht und die Befallsfläche des Jahres 2000 verringert werden müsste. 38


38 5.6 Synthese Die Auslöser der Gradationswelle seit 1993 sind bis heute umstritten. Einfacher war die Situation in den 80er Jahren, als 1983 und 1984 zwei Gewitterstürme insgesamt 173ha Waldbestände zu Boden rissen. Damals wurde die Entscheidung getroffen, die Windwürfe in der Naturzone (etwa 83 ha) nicht aufzuarbeiten. In den folgenden Jahren kam es zu einem geradezu klassischen Verlauf einer Käfergradation. Zunächst besiedelten die Buchdrucker die vom Wind geworfenen Stämme und brachten, nachdem die Populationsdichte angestiegen war, seit 1986 sogar lebende Fichten in der Umgebung von Windwurfflächen zum Absterben. Bereits 1988 wurde der Höhepunkt des Borkenkäferbefalls überschritten und der Neubefall ging bis 1991 kontinuierlich zurück. Damit gelang es der Buchdruckerpopulation aufgrund der ungünstigen thermischen Situation - trotz günstiger Startbedingungen durch zahlreiche Windwürfe und einem riesigen Habitatangebot - nicht, ihre Massenvermehrung fortzusetzen. Zu stark sind die Käfer als wechselwarme Organismen in ihrem Verhalten und ihrer Entwicklung von physikalischen Umweltbedingungen abhängig. Bei ungünstiger Frühjahrs-Witterung können sie nicht oder erst verspätet schwärmen. Außerdem verlängert sich ihr Entwicklungszyklus vom Ei bis zum ausgereiften Käfer bei niedrigen Temperaturen erheblich bzw. ist bei weniger als 11,8 C ga nz gestoppt. SCHOPF und KOEHLER (1995). die damals die Entwicklung mit einer Feldstudie begleiteten, konnten neben der Witterung keine anderen Ursachen für den Rückgang der Buchdruckerpopulation feststellen. Allerdings überdauerten viele Befallsherde auf niedrigem Niveau. Auch wenn in den 90er Jahren - anders als in den 80er Jahren - großflächige Windwürfe als Auslöser für die Befallsentwicklung fehlten, reichen die vorliegenden Informationen aus, die Entstehung der zweiten Gradationswelle zu erklären (NUESS LEIN 1998). Zum einen waren selbst im Jahr 1991, als der Neubefall sein Minimum erreichte, immer noch mehr als 150 Befallsherde im Gebiet vorhanden; zum anderen führten zahlreiche kleinere Windwürfe, die Trockenperiode 1992 sowie die "kräftezehrenden" Vollmasten 1992 und 1995 zu einem ausreichenden Angebot befallstauglicher Fichten. Die Hypothese, dass verstreuter, kleinflächiger Windwurf und Buchdruckerbefall zu einer Gradation führen können, wird auch durch die Ergebnisse einer Computersimulation unterstützt. Mit Hilfe eines Dispersionsmodelles konnte gezeigt werden, dass sich schon bei einem Anteil von weniger als 3 Ofo Prozent geschwächter Bäume die Wahrscheinlichkeit für den Ausbruch einer Massenvermehrung um ein Vielfaches erhöht (FAHSE und HEURICH in Vorbereitung). Diese Auslöser - zusammen mit den günstigen thermischen Bedingungen im warmen Sommer 1992, im "Jahrhundertsommer" 1994 und den folgenden Jahren - ermöglichten es den Käfern, sich das optimale Habitatangebot der Hochlagen zu erschließen, was vorher auf Grund der rauen Witterungsbedingungen nicht möglich war. Es kam zu einer "explosionsartigen" Vermehrung des Buchdruckers, die in ihrem Ausmaß viele Fachleute überraschte. Solange sich die Buchdrucker Abb. 35 "Schrotschussartiger" Borkenkäferbefall in den ausgedehnten Fichtenwäldern nördlich des Farrenbergs. Im Hintergrund sieht man Kahlflächen, die im Zuge der Borkenkäferbekämpfung im Nationalpark Sumava entstanden sind. 39



39 in den großflächigen Fichtenaltbeständen radial ausbreiten konnten, nahm die Totholzfläche exponentiell zu. Nachdem die befallstauglichen Fichten in den zentralen Bereichen der Hochlagen um den Lusen abgestorben waren, konnten sich die Buchdrucker im wesentlichen nur noch linear in zwei Richtungen (Rachel, Siebensteinfelsen) ungebremst ausbreiten, da auf tschechischem Staatsgebiet befallene Bäume entfernt wurden und auf bayerischer Seite aufgrund ausgedehnter Laubholzpartien und relativ junger Bestände eine Dispersion in' die Oberen Hanglagen größtenteils nur überfichtenreiche Talungen möglich war. Bei der Massenvermehrung des Buchdruckers im Nationalpark Bayerischer Wald kommt den von SCHWENKE (1981) genannten Regulationsmechanismenjeweils eine ganz unterschiedliche Bedeutung zu. So konnten Antagonisten, innerartliche Konkurrenz und endogene Faktoren bisher keinen wesentlichen Einfluss auf die Populationsdynamik des Buchdruckers nehmen. Allein das Zusammenspiel zwischen Habitatangebot und Witterungsverlauf scheint bislang die maßgebliche Steuergröße der Populationsentwicklung des Buchdruckers zu sein. Inwieweit "neuartige Waldschäden" im weitesten Sinn z. B. durch kumulative Effekte zu einervitalitätsminderung der Bäume beigetragen haben und ob dieser Prozess die Toleranz gegenüber Stressfaktoren wie Trockenheit und Mast jahre vermindert haben, so dass die Bäume anfälliger gegenüber Buchdruckerbefall wurden, lässt sich nicht eindeutig nachweisen. Die These, dass immissionsbelastete Fichten bevorzugt vom Buchdrucker befallen werden, ist wissenschaftlich umstritten. Einen Überblick über die aktuelle Diskussion gibt WEISSBACHER (1997). Da sich die Befallsentwicklung im Nationalpark Bayerischer Wald jedoch auch ohne "neuartige Waldschäden" hinreichend erklären lässt und einige Argumente gegen eine "Waldschadenshypothese" angeführt werden können, wird ihr im Rahmen dieser Arbeit keine große Bedeutung zugemessen. Abb.36 Aufgrund ausgedehnter Laubholzpartien in den Hanglagen erfolgte dort die Ausbreitung der Käfer vor allem über fichtenreiche Talungen. l 40
40 Auch die Erfahrungen aus anderen Gebieten unterstreichen die Bedeutung von Habitat- und Witterungsbedingungen für die Entwicklung von Borkenkäferpopulationen. So wird für den Zusammenbruch der Borkenkäfermassenvermehrung nach den Stürmen 1868/70 als wesentlicher Grund die nasskalte Witterung genannt und nicht die (damals noch unzureichende) Bekämpfung (ZIERL 1972). Auch WHISTLING (1950) bringt den Zusammenbruch der Borkenkäferkalamität in den Jahren von 1944 bis 1949, die auftschechoslowakischem Staatsgebiet nicht bekämpft wurde, mit der kühlen und feuchten Witterung in der Schwärmzeit des Borkenkäfers in Verbindung. Der Rückgang der aktuellen Borkenkäfermassenvermehrung in Alaska wird ebenfalls durch kühlfeuchte Sommer und den Rückgang befallstauglicher Fichte erklärt (USDA FOREST SERVICE 2001). Ein weiterer Faktor, der eine Massenvermehrung stoppen kann, sind sehr niedrige Wintertemperaturen (OKOLOW 1991). Die dazu notwendigen Temperaturen, -30 C bis -40 C, werden in Mitteleuropa jedoch nur in Ausnahmejahren erreicht. Die alten Fichtenbestände der Hochlagen sind durch Buchdruckerbefall mittlerweile bis auf Jungbestände und einzelne Altbäume (HEURICH 2001) fast vollständig abgestorben. Gleichzeitig wird in den angrenzenden Hanglagen die Ausbreitung des Käfers durch einen hohen Anteil nicht gefährdeter Bestände gehemmt. Damit hat sich das Habitatangebot für die Käfer seit 1991 wesentlich verschlechtert. Möglicherweise ist in den nächsten Jahren mit einem Rückgang des Befalls zu rechnen, insbesondere dann, wenn über einen Zeitraum von mehreren Jahren ungünstige thermische Bedingungen die Vitalität und die Vermehrungsrate des Buchdruckers beeinträchtigen. In den dann vermutlich kleineren Befallsherden könnten auch Antagonisten, Larvenkonkurrenz und stochastische Prozesse lokal zu einem Ende des ein oder anderen Käferherdes beitragen. Über die ganze Naturzone hinweg gesehen wird es aufgrund jederzeit auftretender Windwürfe immer wieder zu neuem Befall kommen. Günstige Witterung dürfte dazu führen, dass die Massenvermehrung weiter anhält, bzw. der Befall auch nach einem Populationsrückgang jederzeit wieder rasch ansteigen kann, wie dies 1992 der Fall war. 41
41 Die Buchdruckermassenvermehrung im Nationalpark Bayerischer Wald Okanartige Stürme in den Jahren 1983 und 1984 warfen im Rachel-Lusen-Gebiet unzählige Bäume auf einer Gesamtfläche von 173 ha zu Boden. Betroffen waren vor allem fichtenreiche Bestände auf Weich böden in den Tal- und Hanglagen des Grenzkammes, lediglich 14,3 ha der Sturmwürfe ereigneten sich in den Hochlagen. Entsprechend der Zielsetzung des Nationalparks wurden die Sturmwürfe innerhalb der Naturzone - etwa die Hälfte der Windwurfgesamtfläche - nicht aufgearbeitet und der Borkenkäfer dort in der Folgezeit nicht bekämpft. Aufgrund des großen Brutraumangebotes kam es zu einem starken Anstieg der Käferpopulation, der schließlich auch zum Befall gesunder Fichten führte. Nach einem Rückgang der Buchdruckeraktivität Anfang der 90er Jahre stieg die Fläche der jährlich abgestorbenen Altfichten seit 1992 zunächst langsam, danach sprunghaft an und blieb anschließend auf hohem Niveau. Um die Auswirkungen dieser Massenvermehrung zu dokumentieren, werden seit 1988 jährlich Befliegungen durchgeführt. Anhand der erstellten Luftbilder lassen sich die vom Buchdrucker abgetöteten Altfichten genau kartieren. Die bisher letzte Befliegung fand im Herbst 2000 statt. Die Auswertung zeigte, dass sich die Buchdruckermassenvermehrung unvermindert fortgesetzt hat. So stieg die Totholzfläche im Jahr 2000 um 605 ha an. Dies ist nach 1996 (827 ha) der zweithöchste Wert seit Beginn der Beobachtung. Damit hat sich die Fläche der abgestorbenen Fichtenbestände auf insgesamt ha - entsprechend 29 % des Rachel-Lusen Gebietes - erhöht. Am stärksten sind die Hochlagen betroffen, wo bis heute 85 % der Waldbestände abgestorben sind. Auch in den anderen Höhenstufen sind mittlerweile größere Totholzkomplexe entstanden, der Anteil abgestorbener Bestände ist hier jedoch weit geringer. Er beträgt in der oberen Hanglage 26 0 /0, in der unteren Hanglage 10 0 /0 und in den Tallagen 11 0/0. Zum Schutz angrenzender Wirtschaftswälder wird der Borkenkäfer im Nationalpark in einer mindestens 500 m breiten Randzone bekämpft. Im Falkenstein-Rachel-Gebiet wird darüber hinaus der Borkenkäfer mit Ausnahme der ha großen Naturzone auf der ganzen Fläche bekämpft. Der höchste "Käferholzeinschlag" wurde 1998 mit über Efm erreicht, seitdem ging dieser stetig zurück. Ein weiteres Indiz für die Effektivität der Borkenkäferbekämpfung ist auch die Verteilung der durch die Käferbekämpfung entstandenen Kahlflächen im Randbereich. Während diese 1997 noch mehr oder weniger gleichmäßig verteilt waren, konzentrierte sich der Holzeinschlag im Jahr 2000 auf die unmittelbare Nachbarschaft zur Naturzone. Mit zunehmender Entfernung zur Naturzone ging der Borkenkäferbefa II sta rk zu rück. Um die Bedeutung der Witterungsbedingungen für die Populationsdynamik des Buchdruckers zu untersuchen, wurden Klimakennwerte der letzten 20Jahre von den Wetterstationen Arber (Hochlagel, Waldhäuser (Hang lage) und Klingenbrunn Bahnhof (Tallage) analysiert. Es zeigte sich, dass der erste Tag mit einer Maximaltemperatur über 20 oe (potenzieller Schwärmtag) sowie die Summe dieser potenziellen Schwärmtage in den Monaten April und Mai einen Erklärungsbeitrag für den Rückgang des Buchdruckerbefalls Anfang der 90er Jahre leisten können. Noch deutlicher war der Zusammenhang zwischen dem Temperaturverlauf in der Vegetationsperiode und der Veränderungsrate der Totholzflächen gegenüber dem jeweiligen Vorjahr. Die für die Populationsdynamik von Insekten maßgeblichen Regulationsmechanismen Habitatangebot, Antagonisten, endogene Faktoren, Konkurrenz und Witterungsverlauf wurden auf der Basis aktueller Forschungsergebnisse diskutiert. Es zeichnet sich ab, dass das Zusammenspiel zwischen Witterungsverlauf und Habitatangebot die wichtigste Steuergröße für die Populationsentwicklung des Buchdruckers im Nationalpark Bayerischer Wald ist. 42
42 Bark beetle outbreak in the Bavarian Forest National Park In 1983 and 1984 hurricane like storms uprooted countless trees in the Rachel-Lusen area of the Bavarian Forest National Park, especially spruce forest stands on swampy an weak soil in the lowland spruce forests and to a lesser extent, the mountainmixed forests of the park. The total area of disturbed forests was 173 ha. Only 14.3 ha of the disturbance occured in the mountain spruce forest. Over all, halfthe affected areas were in the Park's no management zone and, as such, these areas were left alone. Due to the huge amount of breeding material, the bark beetle was able to reproduce quickly and populations became large enough to attack and kill healthy trees. By the beginning of the 1990's bark beetle activity had declined and the total area of spruce stands being attacked began to decrease slowly. However, in 1992 beetle activity sudden Iy i ncreased aga i n a nd has since remained on a high level. Aerial photographs have been taken annually since 1988 and allow for the precise mapping of old dead forest stands and for the effect of bark beetle outbreaks to be documented. The most recent aerial photos were taken on the 24th September Analysis of these photographs indicates that bark beetle populations remain high. protect private forests along the borders of the Park. Active forest managment is also carried out in all areas of the Falkenstein-Rachel area, with exception to hectares of core zone. The highest amount of so-called "bark beetle timber cut" was m' in Since then "bark beetle cut" has decreased to ha If of th is a mou nt. The distribution of clear cuts caused by forest management in the margin zone, provide further proof of the effectiveness of bark beetle management. In 1997 most of the clear cuts were distributed around the margin zone. However since the year 2000, management activities have been mainly undertaken close to the core zone. Since then, evidence of bark beetle damage has decreased as the distance from the core zone increases. It was assumed that the weather plays an important role in the population dynamics of the bark beetle and as such, weather data from the last 20 years was collected and analysed. The data relating to mountainous spruce stands was taken from the Arber weather station, whilst, data relating to the lowland spruce forests was taken from the Klingenbrunn-Bahnhof weather station. In the year 2000, the total stand area killed by the beetle increased by 605 ha. This is the second highest peak since the start of monitoring activities, the highest being in 1996 with 827 ha of forest stands killed, the total area has now increased to ha, equivalent to 29 % of the Rachel-Lusen area. The mountainous spruce forests are the main affected areas within the Park with 85 % ofthese forests hav,ing been killed. Large complexes of dead trees can also be found at other elevations, however the proportion of killed trees present is lower than in the mountainous spruce forests. Up to 36 % of mountain mixed forests stands and 11 % of lowland spruce forest stands have also been killed by beetle activity. A 500 m strip of forest in the National Park is subject to active forest management in order to This data showed that the decline of the bark beetle in the early nineties correlates to a reduction in potential swarm days during this period (potential swarm days occur when the temperature reaches 20 ' Cl. The relationship between the average temperature in the vegetation period and the rate of change in the area of the dead stand, when compared to the previous year was even more significant. The dynamics of insect populations in respect to its major influencing factors (i.e. habitat, natural enemies, competition and weather patterns) were analysed on the basis of these latest research resu Its. It is clear that weather patterns and habitats "are the most important regulatory factors on the dynamics ofthe bark beetle populations. 43
43 Literaturverzeichnis ANDERBRANT, 0., SCHlYTER, F. und G. BIRGSERSSON. (1985): I ntraspecific com petition affecti ng parents and offspring in the bark beetle Ips typographus. Oikos 45; S BAUMGARTEN, M. (1999): Charakterisierung des physiologischen Zustands von Altbuchen in zwei Höhenlagen des Bayerischen Waldes unter Berücksichtigung der Standortfaktoren und der Ozonbelastung. Dissertation an der Forstwissenschaftlichen Fakultät der Ludwig-Maximilians-Universität München; 235 S. BAYERISCHE LANDESANSTALT FUER WALD UND FORST WIRTSCHAFT (1998): Borkenkäferproblematik im Nationalpark Bayerischer Wald - Ergebnis des internationalen Expertengremiums; 47 S. BAYERISCHE LANDESANSTALT FUER WALD UND FORST WIRTSCHAFT (2000) Waldzustandsbericht 2000; 58 S. BEAVER, R. A. (1974): Inraspecific competition among bark beetle larvae. J. Anim. Ecol. 43; S BECKER, T. und H. SCHROETER (2000): Ausbreitung von rindenbrütenden Borkenkäfern nach Sturmschäden. AFZ/Der Wald, 6/2000; S BERRYMAN, A. A. (1974): Dynamics of bark beetle populations: towards a general productivity model. Environ. Entomol. 3; S BEUDERT, B., BREIT, W., DIEPOLDER, U. und M. KAISER (1997): Integrated Monitoring im Nationalpark Bayerischer Wald. Bericht im Auftrag des Umweltbundesamtes; 265 S. BEUDERT, B. und W. BREIT (2001): Integrated Monitoring Programm an der Meßstelle Forellenbach im Nationalpark Bayerischer Wald. Bericht im Auftrag des Umweltbundesamtes; 92 S. BUETINER (1927): Forstamtsgeschichte Zwiesel Ost. Archiv NPV; 59 S. BYERS, J. A. (1989): Behavioral mechanisms involved in reducing competition in bark beetles. Holarctic Ecology, 12; S BYERS, J. A. (1993): Avoidance of competition by spruce bark beetles Ips typographus and Pityogenes chalcographus. Experientia, 49; S COElN, M., YANZHANG, N. und E. FUEHRER (1996): Entwicklung von Fichtenborkenkäfern in. Abhängigkeit von thermischen Bedingungen verschiedener montaner Waldstufen (Coleoptera: Scolytidae). Entomol. Gener., 21; S COElN, M. (1997): Grundlagen für ein Thermoenergetisches Modell zur Fernüberwachung der Borkenkäferentwicklung. Dissertation, Institut für Forstentomologie, Forstpathologie und Forstschutz BOKU Wien. ElLlNG, W., BAUER, E., KLEMM, G. und H. KOCH (1987): Klima und Böden - Waldstandorte. Wissenschaftliche Schriftenreihe. Bayerisches Staatsministerium für Ernährung, Landwirtschaft u. Forsten, München; 255 S. ESCHERICH (1923): Die Forstinsekten Mitteleuropas. Bd. 2., Verlag Paul Parey Berlin. FUEHRER, E. und M. COElN (1998): Thermoenergetisches Modell zur Fernüberwachung der Borkenkäferentwicklung. AFZ GIETl, J. (1990): Untersuchungen zur Bulk Deposition im Einzugsgebiet der Großen Ohe. Landwirtschaftliches Jahrbuch, Sonderheft 1990, 67; S GRAD, M., GREUNE, A. und R. MOESSMER (1992): Totholz im Nationalpark Bayerischer Wald. Entwicklung des Anteils abgestorbener Fichten nach den Windwurfereignissen 1983/84. Auswertung von Farbinfrarot Luftbildern Unveröff. Bericht Bayer. Forstliche Versuchs- und Forschungsanstalt München; 15 S. + Anh. 44
44 GRAD, M., GREUNE, A. und R. MOESSMER (1993): Totholz im Nationalpark Bayerischer Wald. Entwicklung des Anteils abgestorbener Fichten nach den Windwurfereignissen 1983/84. Auswertung von Farbinfrarot-Luftbildern Unveröff. Bericht Bayer. Forstliche Versuchs- und Forschungsanstalt München; 14 S. HERRLEIN (1878): Der Fichten-Barken-Käfer. Dessen Auftreten, Vermehrung und Verbreitung im Bayerischen- und im Böhmer Walde. F.W. Keppler'sche Buchdruckerei. Passau. HEURICH, M. (2001): Waldentwicklung im montanen Fichtenwald nach großflächigem Buchdruckerbefall im Nationalpark Bayerischer Wald. In: Waldentwicklung im Bergwald nach Windwurf und Borkenkäferbefall. S JEHL, H. (1997): Das Wetter aus dem Blickwinkel der gegenwärtigen Borkenkäfergradation. Unveröffentlichter Bericht der Nationalparkverwaltung ; 7 S. KAHLE, H.-P.(1994): Modeliierung der Zusammenhänge zwischen der Variation von klimatischen Elementen des Wasserhaushaltes und dem Radialzuwachs von Fichten aus Hochlagen des Schwarzwaldes; 161 S. LOBINGER, G. (1994): Die Lufttemperatur als limitierender Faktor für die Schwärmaktivität zweier rindenbrütender Fichtenborkenkäferarten. Anz. Schädlingskde., Pflanzenschutz, Umweltschutz 67; NETHERER, S. (2000): Aspekte einer umfassenden Risikoanalyse bezüglich Ips Typographus L. Einschätzung und räumliche Darstellung standorts- und bestandesbezogener Gefährdungspotentiale unter Anwendung kurzund langfristiger Bewertungsinstrumente. Unveröffentlichter Bericht des Instituts für Forstentomologie, Forstpahtologie und Forstschutz. Boku. Wien. NIEHAUS-WUNDERWALD, D. (1992): Biologie der Buchdruckerarten. Wald und Holz 6/92. NUESSLEIN, 5., GREUNE, A., ADLER, H., TROYCKE, A., FAISST, G., REIMEIER, S. und R. DIETRICH (1997): Totholzflächen und Waldstrukturdaten im Nationalpark Bayerischer Wald 1996/1997. Unveröff. Bericht Bayer. Landesanstalt für Wald und Forstwirtschaft; 18 S. + Anh. NUESSLEIN, S. (1997): Wie wirksam ist die Borkenkäferbekämpfung? Erkenntnisse und Erfahrungen aus dem Bayerischen Wald. Forstinfo 5/97, Bayer. Staatsforstverwaltung. NUESSLEIN, S. und G. FAISST (1998): Waldentwicklung im Nationalpark Bayerischer Wald Totholzflächen und Waldverjüngung. Bayerische Landesanstalt für Wald und Forstwirtschaft; 24 S. NUESSLEIN, S. FAISST, G. WEISSBACHER, A. MORITZ, K. ZIMMERMANN, L. BITIERSOHL, J. KENNEL, M. TROYCKE, A. und H. ADLER (1999): Zur Waldentwicklung im Nationalpark Bayerischer Wald Bayerische Landesanstalt für Wald und Forstwirtschaft; 47 S. OKOLOW, C. (1991): Der Einfluss der Forstwirtschaft auf die natürlichen Begrenzungsfaktoren des Buchdruckers. Seevögel 12/Sonderheft; S PICARD, B., ARBEITER, c., MEDERER, M. und M. TILL (1999): Jahrringbau von Fichten im Bereich der Borkenkäfer-Massenvermehrung in den Hochlagen des Nationalparks Bayerischer Wald. Unveröffentlichte Diplomarbeit. Fachhochschule Weihenstephan. PLOCHMANN, R. (1961): 150 Jahre Waldbau im Staatswaldgebiet zwischen Osser und Dreisessel. Forstwissenschaftliche Forschungen. Beihefte zum Forstwissenschaftlichen Zentralblatt, Heft 13. PRELL, H. (1948): Kampf dem Borkenkäfer. Neumann Verlag. Radebeul und Berlin; 31 S. 45
45 RALL, H. (1995): Die Wälder im Nationalpark Bayerischer Wald: Von forstwirtschaftlicher Prägung zur natürlichen Entwicklung. In: Nationalparkverwaltung Bayerischer Wald (Hrsg.): Nationalpark Bayerischer Wald - 25 Jahre auf dem Weg zum Naturwald, Grafenau; S SCHERZlNGER, W. (1996): Naturschutz im Wald: Qualitätsziele einer dynamischen Waldentwicklung. Ulmer, Stuttgart. 447 S. SCHOPF, R., KOEHLER, U., GRAD, M., TIMANS, U., GREUNE, A. und B. KRAUSSE-OPATZ (1993): Untersuchungen zur Populationsdynamik der Borkenkäfer im Nationalpark Bayerischer Wald. Unveröffentlichter Forschungsbericht; 169 S. SCHOPF, R. und U. KOEHLER (1995): Untersuchungen zur Populationsdynamik der Fichtenborkenkäfer im Nationalpark Bayerischer Wald. In: Nationalparkverwaltung Bayerischer Wald (Hrsg.): Nationalpark Bayerischer Wald - 25 Jahre auf dem Weg zum Naturwald, Grafenau; S SCHWENKE, W. (Hrsg.) (1974): Die Forstschädlinge Europas. Bd. 11. Käfer. Verlag Paul Parey, Hamburg, Berlin; 500 S. SCHWENKE, W. (1981): Leitfaden der Forstzoologie und des Forstschutzes gegen Tiere. Parey, Hamburg, Berlin;. 188 S. SCHWERDTFEGER, F. (1974): Die Waldkrankheiten. Ein Lehrbuch der Forstpathologie und des Forstschutzes. 4. Auflage. Hamburg und Berlin; 486 S. SKEFFINGTON, R. A. und A. R. Mc LEOD (1996): Critical levels for ozone of tree species of forest trees as judged by the Liphook Forest Fumigation Experiment. In: Kärenlampi, L. und Skärbi, L (eds.): Critical Levels for Ozone in Europe: Testing and Finalizing the Concepts. UN-ECE Workshop Report. University of Kuopio. Finnland; S STRUNZ, H. (1994a): Totholz im Nationalpark Bayerischer Wald. Entwicklung des Anteils abgestorbener Fichten. Auswertung von Farbinfrarot-Luftbildern Nationalparkverwaltung Bayerischer Wald; 13 S. + Anh. STRUNZ, H. (1994b): Sägen und bekämpfen oder einfach zusehen? Wie man mit Borkenkäferbekämpfung den Böhmerwald ruiniert. Nationalpark, Grafenau; Nr. 82; S STRUNZ, H. (1995): Entwicklung von Totholzflächen im Nationalpark Bayerischer Wald - Luftbildauswertungen und Folgerungen. In: Nationalparkverwaltung Bayerischer Wald (Hrsg.): Nationalpark Bayerischer Wald - 25 Jahre auf dem Weg zum Naturwald, Grafenau; S STRUNZ, H., REINELT, A., WAGNER, S. und M. WOTSCHIKOWSKY (1995): Totholz im Nationalpark Bayerischer Wald. Entwicklung des Anteils abgestorbener Fichten. Auswertung von Farbinfrarot-Luftbildern Nationalparkverwaltung Bayerischer Wald; 24 S. + Anh. STRUNZ, H. und A. REINELT (1996): Totholz im Nationalpark Bayerischer Wald. Entwicklung des Anteils abgestorbener Fichten. Auswertung von Farbinfrarot-Luftbildern Nationalparkverwaltung Bayerischer Wald; 36 S. + Anh. TRAENKNER, H. und E. KENNEL (1989): CIR-Luftbildauswertung 1988/89: Borkenkäfer Nationalpark Bayerischer Wald. Unveröff. Bericht Bayer. Forstliche Versuchs- und Forschungsanstalt München; 7 S. + Anh. TRAENKNER, H., TROYCKE, A. und R. MOESSMER (1989): Waldschäden im Luftbild - Borkenkäferschäden im Nationalpark Bayerischer Wald. Unveröff. Bericht Bayer. Forstliche Versuchs- und Forschungsanstalt München; 10 S. 46
46 TRAENKNER, H., TROYCKE, A., ADLER, H., GRAD, M., GREUNE, A. und R. MOESSMER (1990): Waldschäden im Luftbild: Totholzkartierung im Nationalpark Bayerischer Wald 1981,1988, 1989 und Unveröff. Bericht Bayer. Forstliche Versuchs- und Forschungsanstalt München; 30 S. + Anh. TRAENKNER, H., GRAD, M., GREUNE, A. und R. MOESSMER (1991): Totholz im Nationalpark Bayerischer Wald. Entwicklung des Anteils abgestorbener Fichten nach den Windwurfereignissen 1983/84. Auswertung von Farbinfrarot-Luftbildern Unveröff. Bericht Bayer. Forstliche Versuchs- und Forschungsanstalt München; 40 S. USDA FOREST SERVICE (Hrsg.) (2001): Forest Insect and Disease Conditions in Alaska General Technical report R10-TP-86; 67 S. WEISSBACHER, A. (1998): Begleitende Untersuchungen zur Massenvermehrung des Buchdruckers im Nationalpark Bayerischer Wald Unveröffentlichter Bericht der Bayerischen Landesanstalt für Wald und Forstwirtschaft; 41 S. WEISSBACHER, A. (1999): Begleitende Untersuchungen zum Buchdruckermassenbefall im Nationalpark Bayerischer Wald Unveröffentlichter Bericht der Bayerischen Landesanstalt für Wald und Forstwirtschaft; 22 S. WEISSBACHER, A. (2000): Untersuchungen zum Buchdruckerbefall im Nationalpark Bayerischer Wald 1999/2000. Unveröffentlichter Bericht der Bayerischen Landesanstalt für Wald und Forstwirtschaft; 22 S. WEISSBACHER, A. (2001): Untersuchungen zum Buchdruckerbefall im Nationalpark Bayerischer Wald 1999/2000. Unveröffentlichter Bericht der Bayerischen Landesanstalt für Wald und Forstwirtschaft; 33 S. WHISTLING (ca. 1950): Ein Vergleich drängt sich auf. Gedanken zur Borkenkäferkalamität im bayerischen Wald. AFZ Nr. 24; S ZIERL, H. (1972): Der Hochwald. Forstwissenschaftliche Forschungen. Beihefte zum Forstwissenschaftlichen Centralblatt. Heft 33; 80 S. 47
47 Abbildungsverzeichnis Zur Einführung Scherzinger: S. 7 u. 8 Panorama Rall: S. 10 u. 11 Archiv Nationalparkverwaltung: 30, 32, 33 Eder: 8 Heurich: 4, 6, 7, 24,31, 34 Pöhlmann: 14 Scherzinger: 2, 26, 27 Strunz:3,9, 18,28,29,35,36 Weißbacher: 5 48
48 Die Waldentwicklung nach Windwurf in den Hochlagen des Nationalparks Bayerischer Wald von Hans Jehl 1. Einleitung Das Untersuchungsgebiet Naturräumliche Gegebenheiten Waldaufbau Methoden Flächenauswahl und -design Datenerhebungen Vegetationsaufnahme Strukturaufnahme...; Temperatur und Luftfeuchte Berech n u ng agg reg ierter Besta ndesken nwerte und I nd izes Statistische Auswertu ng und verwendete Softwa re Ergebnisse Aufbau des Baumholzbestandes Kleinstrukturen Kraut- und Moosschicht Naturverjüngung Dichte Keimungssubstrat (Kleinstandorte) Höhenentwicklung Einfluss von Reh und Rothirsch Diskussion Störungen als Motor der Waldentwicklung Veränderungen nach dem Sturm Entwicklung einer neuen Waldgeneration Schlussbemerkungen Zusammenfassung Summary... ; Literaturverzeichnis
, für das Gebiet des Nationalparks Bayerischer Wald")

, die ihren vermeintlichen Höhepunkt mit den Stürmen \"VlVIAN\"")
49 Die Waldentwicklung nach Windwurf Ein Blick in die Archive zeigt jedoch, dass verheerende Stürme die Forstwirtschaft von Anbeginn an begleiten. So sind z. B. für die zweite Hälfte des 19. Jahrhunderts mehrere Stürme mit extrem hohen Schadholzanfällen in Deutschland belegt (Dezember 1868, Oktober 1870, Februar 1894; KARIUS 1990), für das Gebiet des Nationalparks Bayerischer Wald konnten ELLING et al. (1987) seit Mitte des 18. Jahrhunderts ca. 90 größere Sturmwurfereignisse anhand der Aufzeichnungen von Forstleuten nachweisen. Abb. 1 Orkanartige Stürme im August 1983 und November 1984 rissen im Nationalpark Bayerischer Wald unzählige Bäume zu Boden. Abb. 2 In den Folgejahren entwickelte sich, ausgehend von den Windwurfflächen, eine Borkenkäfergradation. In den letzten Jahrzehnten wird eine deutliche Zunahme heftiger Stürme mit verheerenden Schäden an Waldbeständen beobachtet (KARluS 1990), die ihren vermeintlichen Höhepunkt mit den Stürmen "VlVIAN" (26./ ) und "WIEBKE" ( bis ) erreichte: In West- und Mitteleuropa fielen in deren Folge ca. 120 Mio. Festmeter (fm) Sturmwurfholz an, alleine in Bayern betrug die Schadholzmenge etwa 23 Mio. fm (LWF 1995). Aber bereits wenige Jahre später wurde das Schadensausmaß dieser als "Jahrhundertereignisse" eingestuften Orkane weit übertroffen: Am 26. Dezember 1999 überquerte das Orkantief "LOTHAR" mit Windgeschwindigkeiten bis 226 km/h (gemessen am Wendelstein, DWD 1999) West- und Mitteleuropa und hinterließ erneut eine Spur der Verwüstung. Mehr als sechzig Tote, unzählige abgedeckte Häuser und geschätzte 200 Mio. fm Windwurfholz in Europa waren die erschreckende Bilanz dieses Ereignisses. Wenngleich die Forstwirtschaft in den Folgejahren alles daran setzen wird, die Windwürfe zu räumen und wieder aufzuforsten, die Spuren dieser Orkane werden in den Wäldern über Jahrzehnte sichtbar sein. Für die Forstwirtschaft stellen derartige Stürme eine außerordentliche Belastung dar: Die planmäßige Arbeit von Generationen von Forstleuten ist im Handstreich zunichte gemacht, die Aufarbeitung des geworfenen und zerborstenen Holzes 50

.")
.")
 - langfristig angelegte Projekte existieren aber nur wenige.")
50 ist sehr schwierig und gefährlich. Der Holzmarkt ist rasch übersättigt und die schrumpfenden Erlöse für das Windwurfholz decken kaum die Kosten. Die Wiederbewaldung der riesigen Kahlflächen ist erschwert und nimmt viel Zeit in Anspruch. In Naturwäldern sind solche Störungen jedoch ein wichtiger Bestandteil der natürlichen WaIdentwicklung ("disturbance-regime", LORIMER und FRELICH 1994): Sie geben den Anstoß zur Erneuerung von Waldbeständen, verändern über Jahrzehnte oder Jahrhunderte gewachsene Strukturen und tragen so wesentlich zurvielfältigkeitvon Urwäldern bei (FALINSKY 1976). Von Bedeutung ist aber darüber hinaus, dass Wälder nicht nur zahlreiche Strategien entwickelt haben, um sich nach solchen Störungen wieder zu regenerieren, sondern dass eine Vielzahl von Pflanzen-, Pilz- und Tierarten zur Gänze auf die Wiederkehr bestimmter Störungen angewiesen ist (SCHERZlNGER 1996). Zur Wiederbewaldung von Windwurfflächen liegen mittlerweile zahlreiche Einzelerkenntnisse vor - überwiegend jedoch von Untersuchungen außerhalb Europas (z. B. SPURR 1956; PUTZ et al. 1983; SCHAETZL et al. 1988, WHITMORE 1988) - langfristig angelegte Projekte existieren aber nur wenige. Hier bieten Naturwaldreservate und vor allem Wald-Nationalparke, die deutlich größer sind, die einmalige Chance, die Waldentwicklung nach Sturmwurf über lange Zeiträume wissenschaftlich zu beobachten. Aus diesem Grund wurde 1986 damit begonnen, auf großen Windwurfflächen der Jahre 1983 und 1984 im Nationalpark Bayerischer Wald Dauerbeobachtungsflächen einzurichten, dauerhaft zu vermarken und detaillierte Vegetations- und Strukturaufnahmen durchzuführen (HOMANN und ENGELS 1988, FISCHER etal. 1990;JEHL 1995). Abb.3 Die Vogelperspektive macht deutlich, dass die Gewalt der Stürme nicht nur zerstörerisch wirkt, sondern auch eine neue Strukturvielfalt schafft. Abb. 4 Es entstanden unterschiedlich große Lücken in Wald beständen und über Jahrzehnte oder Jahrhunderte gewachsene Strukturen wurden schlagartig zerstört. 51

 ergänzen einander: Erlauben die Aufnahmen auf permanenten Stichprobenflächen, die regelmäßig über die")


51 Abb. 5 Für alte Bäume bedeu ten Windwurf und Borkenkäfer den Tod. Ziel dieser Untersuchungen ist es, die Wiederbewaldung in völlig naturbelassenen sowie in geräumten, aber nicht aufgeforsteten Windwurfund Borkenkäfer-Totholzflächen in den wichtigsten Waldgesellschaften des Nationalparks Bayerischer Wald - Aufichtenwald, Bergmischwald und Bergfichtenwald - sowohl in zeitlicher als auch räumlicher Dimension zu dokumentieren. Dauerbeobachtungsflächen und Waldinventur (siehe HEURICH 2001 b in diesem Band) ergänzen einander: Erlauben die Aufnahmen auf permanenten Stichprobenflächen, die regelmäßig über die Nationalparkfläche verteilt sind, Aussagen auf Landschaftsebene, so vermitteln die wesentlich detaillierteren Untersuchungen auf repräsentativen Dauerbeobachtungsflächen ein tiefergehendes Verständnis natürlicher Abläufe und tragen dazu bei, Ursachen und ihre Wirkungen aufzudecken. In diesem Beitrag werden die Ergebnisse der ersten beiden Aufnahmen auf einer belassenen Windwurffläche in den Hochlagen des Nationalparks Bayerischer Wald dargestellt, zumal die Sorge um die Zukunft der Bergfichtenwälder wegen der seit einigen Jahren zu beobachtenden großflächigen Borkenkäfer-Gradation in der Öffentlichkeit heftige Diskussionen ausgelöst hat. Abb. 6 Andere Pflanzen, wie z. B. das Schmalblättrige Weidenröschen, profitieren jedoch von derartigen Störungen. Wesentliche Fragen dieses vegetationskundlichen Monitorings sind: Wie verändert sich die Vegetation (Moos-, Kraut-, Strauch- und Baumschicht) in den Totholzflächen im Vergleich zu den intakt gebliebenen Altholzbeständen? Welche Bedeutung hat das vielfältige Mosaik unterschiedlichster Kleinstandorte (Wurzelteller, Baumstümpfe, liegendes und stehendes Totholz) für die Wiederbewaldung und die zukünftige Waldstruktur? Darüber hinaus ergeben sich aus diesen Dauerbeobachtungsprojekten noch eine Vielzahl von Einzelfragen, wie z. B. die Bedeutung derartiger Störungen für die floristische und faunistische Artenvielfalt (Biodiversität), die im Rahmen von Detailuntersuchungen erforscht werden. 52







52 2.1 Naturräurnliche Gegebenheiten Die Untersuchungsfläche liegt im Nordostteil des Nationalparks Bayerischer Wald in der WaIdabteilung 33/2 Gfeichtethöh in einer Höhe von 1140 bis 1180 m ü. NN an einem schwach nach Süden geneigten Hang (durchschnittliche Hangneigung ca. r; Abbildung 7). Das Klima in dieser Höhenstufe ist sehr rau und niederschlagsreich (siehe auch HEURICH 2001 b); einen Überblick geben ELLING et al. (1987). Das geologische Ausgangsmaterial bilden Glimmergneise im Wechsel mit grobkörnigem Finsterauer Kristallgranit. Der Talboden und die Einhänge der "Fleischhüttenseige" sind geprägt von verfestigtem, eiszeitlichen Schutt ("Firneisgrundschutt"). Entsprechend dem Ausgangsmaterial entwickelten sich saure bis stark saure Braunerden, z. T. mit Podsolierungserscheinungen (Bodenform "Tiefgründiger Lehm über verfestigtem Schutt" in Anlehnung an die Standortskartierung im Nationalpark Bayerischer Wald; ELLING et al. 1976). Einen bedeutenden Flächenanteil nehmen darüber hinaus noch mineralische und organische Nassböden ein (Abbildung 8). ZWIESEL. FREYUNG Karte 1 Ausschnitt aus der Waldkarte des Nationalparks Bayerischer Wald (Stand ). Maßstab 1:

, Wald-Hainsimse (Luzu/a sy/vatica) und Drahtschmiele (Deschampsia f/exuosa).")
53 Wegen der extremen Standortsbedingungen haben sich in dieser Höhenstufe natürliche Fichtenwälder entwickelt, in die nur vereinzelt Vogelbeeren und, im Übergangsbereich zu den Bergmischwäldern, auch Bergahorne und schwachwüchsige Buchen eingesprengt sind. Auch die Kraut- und Moosschicht wird nur von wenigen Arten geprägt, die aber meist herdenbildend auftreten: Berg Frauenfarn (Athyrium distentifo/ium), Berg-Reitgras (Ca/amagrostis vil/osa), Heidelb~ere (Vaccinium myrtil/us), Wald-Hainsimse (Luzu/a sy/vatica) und Drahtschmiele (Deschampsia f/exuosa). Von Pflanzensoziologen wird diese WaIdgesellschaft als Berg-Reitgras-Fichtenwald (Ca/amagrostio-vil/osae-Piceetum) bezeichnet und zwar in der Subassoziation "-barbi/ophozietosum" (Trennarten sind die Lebermoose Bärlapp- und Flörke-Spitzenbartmoos (Barbi/ophozia /ycopodioides und B. f/oerkeo, zur Differenzierung gegenüber den Aufichtenwäldern der Kaltluft-beeinflussten, nassen Niederungen (Subassoziation "-bazzanietosum'1, in denen v. a. Torfmoose (Sphagnum div. spec.) und das Dreilappige Peitschenmoos (Bazzania tri/obata) charakteristisch sind. Karte 2 Ausschnitt aus der Standortskarte des Nationalparks Bayerischer Wald {7976}. Maßstab 1: Lehm tiefgründiger Lehm über verfestigtem Schutt mittelgründiger Lehm über verfestigtem Schutt gebleichter, mittelgründiger Lehm über verfestigtem Schutt mineralischer Nassboden flaches Niedermoor mittleres und tiefes Niedermoor hochgelegenes Quellmoor Übergangsmoor V anstehender Fels b. schwache Blocküberlagerung «1 0 Ofo der Fläche) extrem ho her Skelettgehalt überdurchschnittlich humusreicher Oberboden. sandig-grusiger Unterboden in cm Tiefe * Vorkommen von verfestigtem Schutt in weniger als 1 m Tiefe Gleye mit deutlichem Go-Horizont und Braunerde-Gleye 11 schlecht zersetzte organische Auflage (bei Nassböden) organische Auflage bei mineralischen Nassböden cm f Mächtigkeit der organischen Auflage bei Mooren cm t Mächtigkeit der organischen Auflage bei Mooren> 100 cm quellige Stellen 54
: 45,3 I-- 99,9 Fichte Oberschicht 0,1 Vogelbeere Baumarten: Unterschicht 100 Vogelbeere Bestocku ngsgrad: 1,07 Beschirmungsgrad: - 0,50 Vorrat")

 entstanden sind (ELLING et al. 1987).")

.")
54 2.2 Waldaufbau Waldbestand 2a 1 ( ~ von... bis Alter Jahre: Mittel 135 Fläche (ha): 45,3 I-- 99,9 Fichte Oberschicht 0,1 Vogelbeere Baumarten: Unterschicht 100 Vogelbeere Bestocku ngsgrad: 1,07 Beschirmungsgrad: - 0,50 Vorrat (fm/ha): Einzelne Vogelbeeren. Sonstige Bemerkungen: Bestand licht geschlossen bis lückig. Unter- und Zwischenstand nicht ausreichend. Stellenweise sehr starkes Altholzgerüst. Einen Eindruck vom Waldaufbau vor dem Windwurfereignis vermitteln die Ergebnisse der Waldinventur Die Untersuchungsfläche liegt in einem Zentrum derjenigen Windwürfe, welche durch die verheerenden Stürme im 19. Jahrhundert (7. und 8. Dezember 1868, 26. und 27. Oktober 1870; Abbildung 7) entstanden sind (ELLING et al. 1987). Da man damals mit der Holzverwertung und Bekämpfung des Borkenkäfers über Jahre hinaus ausgelastet war - neben ortsansässigen WaIdarbeitern waren zahllose Arbeitskräfte aus dem Ausland zur Unterstützung nötig - überließ man in Erwartung natürlicher Ansamung von Fichtenjungwuchs die geräumten Sturmwurfflächen meist sich selbst. Vereinzelt stellte sich Fichten Naturverjüngung auch ein; nach mehreren Jahren jedoch lag der Großteil der Flächen "teils verödet, teils ungemein vergrast und verunkrautet, auch teilweise mit einzelnen älteren lebhaft wachsenden Jungwüchsen durchstellt" da (zitiert nach ZIERL 1972). Deshalb wurde in den 80er Jahren das Hauptaugenmerk auf die Wiederbestockung dieser Flächen - in der Regel durch Pflanzung - gelegt. Ob im Bereich des Untersuchungsgebietes aufgeforstet wurde, ist nicht bekannt. 2a 2 2b 1 2b ,2 31,8 8,5 100 Fichte 99,6 Fichte 0,4 Vogelbeere 50 Rchte, 50 Vogelbeere 100 Fichte keine Angaben 1,02 keine Angaben 0,70 0,90 0,80 keine Angaben 514 keine Angaben... entlang des Baches etwas jünger - stufig aufgebaut. In der Waldabteilung Gfeichtethöh waren die forstlichen Eingriffe in den folgenden Jahrzehnten sicherlich sehr extensiv. Einerseits sah man keine Notwendigkeit zu Durchforstungen in den auf Windwurfflächen herangewachsenen gleichaltrigen Hochlagenbeständen bzw. beschränkte sich auf die Entnahme vom Schnee gebrochener Bäume aus Sorge vor dem Borkenkäfer (ZIERL 1972). Andererseits fehlte es an den notwendigen Erschließungsseinrichtungen: lediglich eine Schlittenziehbahn führte von der Schwarzbachklause (errichtet 1823) in diesen Teil der Hochlagenwälder. Tab. 7 Auszug aus den Ergebnissen der Waldinventur 7982 für die Waldabteilung 33/2 Gfeich tethöh. Abb. 7 Schwere Stürme in der zweiten Hälfte des 79. Jahrhunderts verwüsteten weite Teile des Bayerischen Waldes und Böhmerwaldes. 55



 befallen und abgetötet. Bis 1989 kam der Borkenkäferbefall jedoch fast vollständig zum Erliegen (Abbildung 9).")
55 als sogenannte Reservate ausgeschieden, in denen keine direkte menschliche Einflussnahme mehr erfolgen soll. Auch in der Abteilung Gfeichtethöh fanden seit der Schneebruchaufarbeitung im Jahr 1969 keine forstlichen Eingriffe mehr statt. Abb. 8 Luftaufnahme von der Waldabteilung Gfeichtethöh, erstellt im November In Bildmitte die untersuchte Windwurfflöche. Nachdem es im Winter 1966/67 zu massiven Schneebruchschäden von den Tal- bis zu den Hochlagen kam, wurde 1969 diese Schlittenziehbahn zu einem Schlepper-befahrbaren Rückeweg ausgebaut, das "LusenstraßI" fertiggestellt und die Schneebruchschäden in diesem Bereich aufgearbeitet. Erste tiefgreifende Veränderungen in den Bergfichtenwäldern des Nationalparks lösten mehrere Herbststürme im November 1984 aus: In der Waldabteilung Gfeichtethöh entstanden zwei größere Windwurfflächen von jeweils etwa einem Hektar Größe (Abbildung 8) und einige kleinere Nesterwürfe. In den Folgejahren wurden die Ränder dieser Sturmwürfe vom Borkenkäfer (v. a. Buchdrucker, Ips typographus) befallen und abgetötet. Bis 1989 kam der Borkenkäferbefall jedoch fast vollständig zum Erliegen (Abbildung 9). Erst seit 1996 wird in dieser und den benachbarten Waldabteilungen eine neue Borkenkäfer-Gradation beobachtet, die dort bis heute /0 derverbliebenen Altholzbestände abgetötet hat. Bereits zwei Jahre nach der Gründung des Nationalparks Bayerischer Wald am 7. Oktober 1970 wurden die Wälder der abgelegenen Hochlagen Abb.9 Die Untersuchungsflöche im Frühsommer Zur Dokumentation des Witterungsverlaufs wurde eine Wetterhütte (rechter Bildrand) aufgebaut. 56



56 3.1 Flächenauswahl und -design Zum Verständnis der Eigendynamik von WaIdbeständen sind langfristig angelegte, interdisziplinäre Untersuchungen eine Grundvoraussetzung. In der Naturwaldforschung werden bisher unterschiedlichste Verfahren - Stichproben, Repräsentationsflächen, Transekte - angewendet. Einen Überblick hierüber geben z. B. ALBRECHT (1990) und MEYER et al. (2001). Zur wissenschaftlichen Beobachtung der Waldentwicklung nach großflächigem Sturmwurf im Nationalpark Bayerischer Wald wurde die Transektmethode gewählt. Dieses Konzept hat in diesem Zusammenhang folgende Vorteile: Das Einmessen und Vermarken der Dauerbeobachtungsflächen ist unter den schwierigen Verhältnissen eines Windwurfverhaus einfacher und weniger aufwendig als bei Stichprobenverfahren. Darüber hinaus können die Flächen auch bei Verlust einzelner Markierungen später leicht wieder aufgefunden bzw. rekonstruiert werden. Transekte geben einen anschaulichen Bestandsquerschnitt wieder. Randeffekte spielen beim zusammenhängenden Transekt eine geringere Rolle als bei einzelnen Probekreisen. Im Frühjahr 1992 wurde analog zu den Windwurf-Dauerbeobachtungsflächen in den Tal- und Hanglagen (FISCHER et al. 1990, JEHL 1995) ein 390 m langer Transekt in einer naturbelassenen Sturmwurffläche in der Waldabteilung Gfeichtethöh eingemessen und dauerhaft vermarkt (Abbildung 10). Wesentliches Kriterium für die Auswahl dieser Fläche war neben der Größe - sie ist eine der bedeutendsten Sturmwurfflächen in den Hochlagen des Nationalparks - die Zugänglichkeit: In weniger als dreißig Minuten ist die Untersuchungsfläche zu Fuß von der Schwarzbachklause, zu der eine Forststraße führt, erreichbar. Abb.10 Luftaufnahme der Untersuchungsfläche mit der (ungefähren) Lage des Transekts (November 1991). 57
.")

. Maßstab 1: 10.")
57 Der Transekt wurde quer durch die Windwurffläche gelegt und hat Anschluss an die angrenzenden, von den Stürmen unversehrt gebliebenen Altholzbestände. Dadurch wurden auch die durch Borkenkäferbefall entstandenen Totholzsäume um den Windwurf erfasst (Karte 3). Die Transekte sind 10m breit und wurden in jeweils 100 m 2 große Aufnahmeeinheiten ("Probequadrate") unterteilt (siehe auch JEHL 1995). Für die Anfangsphase der Waldentwicklung nach Windwurf und Borkenkäferbefall, in der rasche Veränderungen zu erwarten waren, wurde ein Aufnah~eturnus von fünf Jahren geplant, dem sich in der weiteren Entwicklung Wiederholungsaufnahmen im zehnjährigen Rhythmus anschließen werden. Karte 3 Ausschnitt aus der Totholzkarte des Nationalparks Bayerischer Wald (Stand ). Maßstab 1: Lebende Bestände Transekt _ Windwurfflächen
 einen Eindruck von den standörtlichen")
 erfasst: Nach Erstellung einer vollständigen Artenliste, getrennt nach den verschiedenen")
58 3.2 Datenerhebungen Vegetationsaufnahme Über die Erfassung der krautigen Vegetation ist die synsystematische Zuordnung zu Pflanzengesellschaften möglich. Außerdem vermittelt die Analyse der Zeigerwerte (Skalierung des ökologischen Verhaltens von Pflanzen bezüglich verschiedener Standortsfaktoren, ELLENBERG 1992) einen Eindruck von den standörtlichen Gegebenheiten eines Untersuchungsgebietes. Zudem stellt die krautige Vegetation als Konkurrenzfaktor eine wesentliche Einflussgröße für die Etablierung der Baumverjüngung dar. Die Vegetation wurde auf den Probequadraten durch pflanzensoziologische Aufnahmen nach BRAUN-BLANQUET (1964) erfasst: Nach Erstellung einer vollständigen Artenliste, getrennt nach den verschiedenen Vegetationsschichten, wurde die Artmächtigkeit über Abundanz (Anzahl) bzw. Dominanz (Deckung) der jeweiligen Art nach einer neunstufigen Skala geschätzt (WILMANNS 1993, in Anlehnung an BRAUN-BLANQUET 1964; Tabelle 2). Der Bewuchs von Felsen und Totholz - es sind dies vor allem Moose und Flechten - wurde nicht berücksichtigt. Die Nomenklatur der Gefäßpflanzen richtet sich nach ROTHMALER (1990), die der Moose nach FRAHM und FREY(1983). Die Sortierung der Arten in dervegetationstabelle erfolgte ~icht streng nach rein pflanzensoziologischen Kriterien, vielmehr sollten auch ökologische Aspekte berücksichtigt werden (Ausscheidung sog. soziologisch-ökologischer Artengruppen). Hat eine Art jedoch diagnostische Bedeutung als sog. Charakterart einer synsystematischen Einheit (Pflanzengesellschaft, vgl. hierzu OBERDORFER 1978, 1992). so ist dies in der Vegetationstabelle durch gebräuchliche Kürzel vor dem Artnamen vermerkt: "0": Charakterart einer Ordnung, hier v. a. boreal-alpine Nadelwälder (Pieeetalia abietis), aber auch mesophytische, buchenwaldartige Laubwälder Mitteleuropas (Fagetalia sylvatieae) und mitteleuropäische Schlag- und Vorwald- Gesellschaften (Atropetalia). "V":Verbandscharakterart; hier v. a. Fichten Tannen- und Fichtenwälder (Pieeion abietis), aber auch Buchen- und Tannen-Buchenwälder (Fagion sylvatieae). bodensaure Schlagfluren (Epilobion angustifolitl und VorwaId Gesellschaften (Sambueo-Salicion). Klammern um die Kürzel bedeuten in diesem Zusammenhang "schwache" Charakterart. Die Berechnung der mittleren Zeigerwerte je Aufnahmefläche erfolgte quantitativ; d. h. dass nicht nur das Vorhandensein einer Art, sondern auch ihre Artmächtigkeit (Abundanz bzw. Dominanz) berücksichtigt wurde. Dazu wurden die Schätzwerte der Artmächtigkeit nach WILMANNS (1993) entsprechend einem Vorschlag von VAN DER MAAREL (1979) in Multiplikatoren (Tabelle 2) umgesetzt. (Das Rechnen mit Zeigerwerten, die streng genommen weder Kardinal- noch Ordinalzahlen, sondern "Platzhalter" für eine Beschreibung des ökologischen Verhaltens von Pflanzen darstellen, wurde u. a. von ROST-SIEBERT (1988) problematisiert. Zahlreiche Untersuchungen jedoch, die sich mit dieser Fragestellung beschäftigten, zeigen BERG 1991). + 2m 2a 2b Abundanz bzw. Dominanz (bezog~n auf di~ Aufnahmdläch~) ein Individuum, Deckung< 5% 2-5 Individu~n, D~ckung <5~ 6-50 Individuen, Deckung< 5% >50 Individu~n, Deckung < 5~ Individuenzahl beliebig, Deckung >5-15% Individuenzahl ~Ii~big, Deckung> 15-2SOtb Individuenzahl beliebig, Deckung >25-500/0 Individuenzahl ~Ii~big, D~ckung >50-75~ Individuenzahl beliebig, Deckung >75-100% Tab. 2 Transformation der Schätzwerte nach WILMANNS (1993) in Multiplikatoren nach VAN DER MAAREL (1979). eine gute Übereinstimmung der mittleren Zeigerwerte mit gemessenen Standortsfaktoren; ELLEN Artmäch tigk~it Mittlere Multipli- D~ckung kator (~) 0,1 1 0,5 2 2,5 3 5,0 4 10,0 5 20,0 6 37,5 7 62,S 8 87,5 9 59
.")
, darüber ein SUUNTO Höhen- und Neigungsmesser. Distanzen wurden mit einem Ultraschall-Entfernungsmessgerät erfasst.")
59 3.2.2 Strukturaufnahme Die Waldstrukturuntersuchungen umfassen die lebenden Bäume und Sträucher, deren Verjüngung und das Totholz (PROJEKTGRUPPE NATURWALD RESERVATE 1993). Darüber hinaus werden Kleinstandorte wie z. B. Wurzelteller, größere Felsen und Baumstümpfe erfasst (Tabelle 3). Die Durchmesser wurden ab Derbholzstärke (7 cm) mit einem Umfangmaßband, darunter mit einer Messkluppe ermittelt. Zur Baumhöhenmessung bis 8 m Höhe diente eine Teleskop Messlatte (Typ MESSFIX), darüber ein SUUNTO Höhen- und Neigungsmesser. Distanzen wurden mit einem Ultraschall-Entfernungsmessgerät erfasst. Bei der Erstaufnahme wurden darüber hinaus in den geschlossenen Altholzbeständen jeweils acht Kronenablotungen je Baum mittels Kronenspiegei zur Ermittlung der Kronenschirmflächen durchgeführt. Wegen der Unsicherheit der Entstehungsgeschichte des untersuchten Waidbestandes (Kapitel 4.1) wurden Bohrspäne zur AItersbestimmung an abgestorbenen (1992) und an frisch vom Borkenkäfer befallenen Fichten (1997) in 1.30 m Höhe entnommen. Besondere Geländestrukturen, wie Fließgewässer, Quellsümpfe oder alte Bringungssysteme wurden bei der Erstaufnahme ebenfalls eingemessen und das Geländeprofil entlang des Transekts durch ein Nivellement erfasst. Die Vermessungsarbeiten erfolgten mit einem WILD Theodolith. Tab. 3 Aufnahmeparameter der Waldstrukturaufnahme. : ist' bei einer Höhe unter ',30 m Aufnahmeparamder Derbholzbestand (d ~ 7cm) lebend stehend tot X!y -Koordinaten Baumart BHD 11! bzw. Durchmesser Kronenansatz Gesamthöhe Breite Anfa ngs-/endd u rch messer Schaftlänge Verbiss Schä 1-/Fegeschaden Vermoderungsgrad Kleinstandort (z. B. Moderholz) Wuchsbesonderheiten (z. B. Zwiesel) liegend Verjüngung (d < 7cm) Baum- Wur- Höhe stümpfe zelteller lebend tot ~O,2m <O,2m _ N 1- Q): :::T.(1) 1- C :::I lo lo n -~ :::I n C :::I ~ :::I r-t- <D:::I <DQ) 1'.)(") '-' :::T tp Q) C 3 ;:+ n :::I -.~ Q) Während auf den Transekten der Tal- und Hanglagen lediglich bei der Verjüngung ab 1.0 m Höhe die Standpunkte genau eingemessen wurden, erfolgte die Einmessung in den Hochlagen bereits bei Bäumen ab 20 cm Höhe. Dies vor allem deshalb, weil für die Etablierung der Verjüngung in den Bergfichtenwäldern offenbar besondere Kleinstandorte Voraussetzung sind, die jedoch bei älteren Pflanzen häufig nicht mehr erkennbar sind, wie z. B. Moderholz als Keimbett (zur Aufnahmemethodik siehe auch JEHL 1995) Temperatur und Luftfeuchte Zur Dokumentation des Witterungsverlaufs während der Vegetationsperiode wird ein Thermohygrograph in 2 m Höhe in einer kleinen Wetterhütte auf der belassenen Windwurffläche betrieben. Mit diesem Messgerät werden sowohl Lufttemperatur als auch relative Luftfeuchtigkeit kontinuierlich registriert. 60
. Die Charakterisierung der Durchmesserverteilung in den geschlossenen Altholzbeständen erfolgte nach v.")
 und eins (die Baumdurchmesservariieren sehr stark) möglich.")

 in Abhängigkeit vom BHD ein Polynom zweiten Grades berechnet (Abbildung 12).")
60 ~ 3.3 Berechnung aggregierter Bestandeskennwerte und Indizes Zur Beschreibung der Durchmesserverteilung in den geschlossenen Altholzbeständen wurden die Bäume entsprechend ihrem Brusthöhendurchmesser in Durchmesserklassen zusammengefasst (Tabelle 4). Die Charakterisierung der Durchmesserverteilung in den geschlossenen Altholzbeständen erfolgte nach v. GADOW und FUELDNER (1992): dabei wird jeweils der Brusthöhendurchmesser eines Baumes in Re lation zu dem seines nächsten (bzw. zweit- oder drittnächsten) Nachbarn gesetzt. Es sind Werte zwischen null (alle Bäume sind gleich dick) und eins (die Baumdurchmesservariieren sehr stark) möglich. Durchmesser in cm Durchmesserklasse 7-14 '- 1 r ' r ' r ~ 65 7 Der mittlere Kronendurchmesser wurde errechnet als geometrisches Mittel aus acht Kronenablotungen je Baum. Während sich die mittleren Kronendurchmesser in Abhängigkeit vom BHD durch eine lineare Regression recht gut beschreiben ließen (Abbildung 11), variierten die Kronenlängen sehr stark: Hier wurde deshalb zur Schätzung der Kronenansatzhöhen (H) in Abhängigkeit vom BHD ein Polynom zweiten Grades berechnet (Abbildung 12). Die Ausgleichsfunktionen waren erforderlich, um später sowohl die mittleren Kronendurchmesser und mittleren Kronenlängen als auch den sog. Plumpheitsgrad (Verhältnis Kronendurchmesser/Kronenlänge) und den relativen Kronenanteil für den Grundflächenmittel- und den Oberhöhenstamm berechnen zu (Kapitel 4. 1). können Das Volumen der stehenden lebenden und toten, jedoch noch nicht abgebrochenen Bäume wurde sowohl mit Hilfe der "Massentafeln" in GANG- HOFERS Praktischem Holzrechner (MANTEL 1950) als auch über waldwachstumskundliche Standardverfahren (FRANZ und KENNEL 1973, KENNEL 1973) berechnet. Durch den Vergleich der Ergebnisse sollten Fehler bei dervorratsermittlung ausgeschlossen werden. Zur Volumenberechnung der abgebrochenen Bäume wurde zunächst über die Bestandeshöhenkurve (Abbildung 13) eine Schätzung der erwarteten Baumhöhe vorgenommen. Verwendet wurde die Schätzgleichung nach PETTERSON (1955). 10 ]: 8 - e< VI cu E.s:::. ~ ::I 'l:i c: cu c: 0 ::z o Brusthöhendurehmesser (ern) Tab. 4 Abgrenzung der Durchmesserklassen. Abb. 11 Kronendurchmesser bei der Fichte in Abhängigkeit vom BHD. Abb. 12 Kronenansatzhöhen bei der Fichte in Abhängigkeit vom BHD. y = O,0775x + 0,671 R 2 = 0, gemessener Kronendurchmesser --Näherungsgerade o y = - O,0065x 2 + O,6397x - 4,0231 R 2 = 0,2983.* Brusthöhendurehmesser (em) Gemessene Kronenansatzhöhe --Näherungskurve 80 61

.")
61 Zur Berücksichtigung der Bruchhöhe wurde eine von KRAMER und AKCA (1995) angegebene Faustregel angewendet, die besagt, dass in einer relativen Baumhöhe von 0,125 vom Stammfuß her gesehen bereits 25 % einer relativen Höhe von 0,3 50 % halben Baumhöhe etwa 75 % des Stammvolumens enthalten sind. des Stammvolumens, in und bei der 40 r ~ 30 :[ ~ 20 :0 ~ 10 o o Abb.13 Gemessene Baumhöhen in Abhängigkeit vom BHD bei der Fichte und errechnete Bestandeshöhenkurve (nach PWERSON 1955). Brusthöhendurehmesser (em) Gemessene Baumhöhe --Bestandeshöhenkurve Die Berechnung des Volumens der liegenden Stämme erfolgte nach SMALIAN (1837, in KRAMER und AKCA 1995). Zur Beschreibung der durchschnittlichen Baumdimensionen in den intakten Altholzbeständen wurden Grundflächenmittel- und Oberhöhenstamm verwendet. Diese sind die in Praxis und waldwachstumskundlicher Forschung wichtigsten Bestandesmittelstämme zur Charakterisierung von Wäldern. Der GrundflächenmitteIstamm ergibt sich aus dem arithmetischen Mittel der Brusthöhenquerschnittsflächen aller Bäume eines Bestandes, aus dem dann der mittlere BHD berechnet werden kann; die dem Grundflächenmitteistamm entsprechende Baumhöhe wird aus der Bestandeshöhenkurve ermittelt. Die Dimensionen des Oberhöhenstammes werden durch das arithmetische Mittel von BHD und Höhe der 100 stärksten Bäume je Hektar oder im Falle des Transekts (0,25 ha Aufnahmefläche) der 25 stärksten Bäume errechnet. Die Analyse der horizontalen Bestandesstruktur (Verteilungsmuster der Bäume einer Art) erfolgte über den Aggregationsindex nach CLARK und EVANS (1954). Dieser beschreibt das Verhältnis vom beobachteten zum erwarteten mittleren Abstand benachbarter Bäume. Er kann Werte zwischen Null (stärkste Klumpung) und 2,1491 (regelmäßige Verteilung nach einem Hexagonalmuster) annehmen; eine zufällige Verteilung ergibt den Wert 1 (PRETZSCH 1993). Der berechnete Aggregationsindex für den Baumholzbestand (BHD ~Jcm) ergab jedoch Werte, die nicht dem optischen Eindruck (Stammfußkarte!) entsprechen. Eine mögliche Ursache besteht in der Plausibilitätskontrolle von SILVA 2.2, die, vereinfacht ausgedrückt, sämtliche Bäume aus der Berechnung ausklammert, deren Distanz zum Rand der Untersuchungsfläche geringer als die Distanz zum nächsten Nachbarn ist. Dadurch wurde der Stichprobenumfang, bedingt durch die Form der Untersuchungsfläche (großer Umfang im Verhältnis zur Fläche), wesentlich reduziert. Eine Möglichkeit zur Beseitigung dieses Mangels besteht darin, Bäume, die in unmittelbarer Nachbarschaft zum Transekt stehen, zukünftig für die Strukturberechnungen mit zu erfassen. Für die Baumverjüngung ließ sich allerdings wegen der großen Individuenzahl ein aussagekräftiger Aggregationsindex berechnen. Die Verteilung einzelner Baumarten, also die Frage, ob eine intensive Durchmischung oder eine klare räumliche Trennung zu beobachten ist, wurde mit dem Segregationsindex nach PIELOU (1977) beschrieben. Für dessen Berechnung wurde ein "Such lauf' für alle Bäume einer Untersuchungsfläche durchgeführt und die Artzuge- 62
 und 1 (räumliche Trennung von Arten) annehmen; Null entspricht einer unabhängigen Verteilung. Die vertikale Bestandesstruktur, d. h.")
: 100-80 Ofo der maximalen Baumhöhe im Bestand; Zone 2 (Mittelschicht): 80-50 Ofo der maximalen Baumhöhe im Bestand; Zone 3")
62 hörigkeit des jeweils nächsten Nachbarn bestimmt (diesen führt das Programm SILVA 2.2 automatisch durch!). Dieser Index kann Werte zwischen -1 (enge Assoziation von Arten) und 1 (räumliche Trennung von Arten) annehmen; Null entspricht einer unabhängigen Verteilung. Die vertikale Bestandesstruktur, d. h. die Verteilung der Baumarten auf drei Höhenschichten, wird durch den Artprofilindex nach PRETZSCH (1996) charakterisiert. Dabei werden folgende "Höhenzonen" ausgeschieden: Zone 1 (Oberschicht): Ofo der maximalen Baumhöhe im Bestand; Zone 2 (Mittelschicht): Ofo der maximalen Baumhöhe im Bestand; Zone 3 (Unterschicht): < 50 Ofo der maximalen Baumhöhe im Bestand. Der maximale Artprofilindex A... ist abhängig von der Anzahl der beteiligten Baumarten. Bei nur einer Baumart beträgt er 1,099. Da sich die Altholzbestände am Anfang und Ende des Transekts bereits optisch unterscheiden, wurden diese Indizes getrennt für beide Abschnitte (Probequadrate 1 bis 9 und 24 bis 39) berechnet. Die Baumstümpfe wurden, analog den stehenden Bäumen (BHD), entsprechend der Durchmesser an Schnitt- bzw. Bruchfläche nach Durchmesserklassen zusammengefasst (Tabelle 4.) Zur Kalkulation der flächenmäßigen Bedeutung dieser sog. Kleinstandorte wurde bei den Baumstümpfen neben der reinen Schnitt- bzw. Bruchfläche auch diejenige Fläche berechnet, welche durch deren dreifachen Durchmesser bestimmt ist ("Wirkungsfläche"). Die Beobachtung zeigte nämlich, dass von diesen Kleinstrukturen eine nicht zu unterschätzende positive Wirkung für die Ansamung und Entwicklung von Naturverjüngung in der unmittelbaren Umgebung ausgeht. Analog wurde bei stehendem Totholz verfahren, allerdings mit dem Unterschied, dass dessen (gedachte) Stammquerschnittsfläche von der berechneten Wirkungsfläche abgezogen wurde. Bei liegenden Stämmen wurde einerseits die projizierte Fläche über Anfangs- und Enddurchmesser sowie deren Länge als auch die doppelte Fläche als Wirkungsfläche berechnet. Die Ergebnisse der Verjüngungsinventur in den Hochlagen weisen auch hier auf einen gewissen positiven Effekt des liegenden Totholzes für die Naturverjüngung hin (HEURICH 2001 b). Der durch die herausgehebelten Wurzeln umgestürzter Bäume freigelegte Mineralboden scheint vorrangig von Pionieren besiedelt zu werden (JEHL 1995, FISCHER und JEHL 1999). Deshalb wurde die Fläche eines Wurzeltellers und somit auch des darunter freigelegten Bodens näherungsweise als halbe Ellipse berechnet. Die Berechnung erfolgte über die gemessene Breite und Höhe des Wurzeltellers. Zur Darstellung der Höhenentwicklung der Baumverjüngung wurden die Bäume in Höhenklassen eingeteilt: Höhe in an Höhenklasse '--- - < ,20-0,49 1 '-- r--- '--,--- r- 0,50-0,99 2 1,00-1,49 3 1,50-1,99 4 2,00-2,49 5 2,50-2,99 6 ~3,00 7 An dieser Stelle möchte ich Herrn Dr. Peter Biber vom Lehrstuhl für Waldwachstumskunde der TU München danken, der mir durch seinen fachkundigen Rat die umfangreichen ertragskundlichen Berechnungen wesentlich erleichtert hat. Tab. 5 Einteilung der Baumverjüngung in Höhenklassen. 63
 auf Signifikanz")
 diente das Programm SILVA 2.")
 wurden mit dem geografischen Informationssystem ArcView 3.2 und Corel Draw 8.0 erstellt.")
63 3.4 Statistische Auswertung und verwendete Software Zur Überprüfung der Regression der Bestandeshöhenkurve (Schätzgleichung nach PmERsoN 1955) wurde ein F-Test durchgeführt. Die Veränderungen in den mittleren Zeigerwerten in den verschiedenen Entwicklungsstadien wurden mit Hilfe eines verteilungsfreien Mediantests (Vierfelder-Chi L Test nach LOZAN 1992) auf Signifikanz überprüft. Da dieser im Vergleich zum T -Test nur eine Trennschärfe von 64 0J0 aufweist, werden nur signifikante Veränderungen (p S. 0,01) thematisiert. Strukturindizes zur Charakterisierung des Bestandsaufbaus (CLARK und EVANS 1954, PIELOU 1977, v. GADOW und FUELDNER 1992, PRmSCH 1996) diente das Programm SILVA 2.2, das vom Lehrstuhl für Waldwachstumskunde der Technischen Universität München entwickelt wurde. Die kartografischen Darstellungen (Stammfußund Kronenkarten, Bestandsaufrisse) wurden mit dem geografischen Informationssystem ArcView 3.2 und Corel Draw 8.0 erstellt. Alle erhobenen Daten sind in einer MS-Access 97- Datenbank gespeichert. Die Berechnungen und die statistische Datenauswertung erfolgten mit MS-Excel 97 und SPSS 10.0; zur Ermittlung der Abb. 14 Geschlossenes Altholz am Beginn des Transekts. Bei dieser Baumgruppe bildete ganz offensichtlich das verrottende Holz eines umgestürzten Baumes das Keimbett ("Ranne'J 64
 im \"geschlossenen Altholz\" ab-, die der Probequadrate im \"stehenden Totholz\" zugenommen (Tabelle 6).")
. Bei den Strukturaufnahmen im Sommer und Herbst 1992 wurden auf dem Transekt insgesamt 2.")
 gezählt. Einen optischen Eindruck vom Bestandsaufbau acht Jahre nach dem Sturm vermitteln Stammfuß- und Kronenkarten und die Bestandsaufrisse in der Anlage. Bestandssituation 1992 1997 PO Nr.")

64 4.1 Aufbau des Baumholzbestandes Der Transekt in der Abteilung Gfeichtethöh umfasst die Bestandssituationen "geschl ossenes Altholz", "stehendes Totholz" und "belassener Windwurf'. Durch erneuten Borkenkäferbefall seit Mitte der 90er Jahre hat die Zahl der Probequadrate (PO) im "geschlossenen Altholz" ab-, die der Probequadrate im "stehenden Totholz" zugenommen (Tabelle 6). Die Fläche des gesamten Windwurfs beträgt 0,82 ha, der Umfang 480 m (ermittelt mit Hilfe von ArcView auf der Grundlage von Color-Infrarot-Luftbildern, die seit 1988 jährlich von einem Flugzeug aus aufgenommen werden). Bei den Strukturaufnahmen im Sommer und Herbst 1992 wurden auf dem Transekt insgesamt Bäume ab 20 cm Höhe (lebend und tot, stehend und liegend) und 199 Kleinstandorte (Wurzelteller, Baumstümpfe) erfasst und lagegetreu eingemessen sowie 934 Keimlinge und Sämlinge (Pflanzen kleiner 20 cm) gezählt. Einen optischen Eindruck vom Bestandsaufbau acht Jahre nach dem Sturm vermitteln Stammfuß- und Kronenkarten und die Bestandsaufrisse in der Anlage. Bestandssituation PO Nr. N pa Nr. "geschlossenes Altholz": Von den Stürmen und nachfolgendem 1 bis bis 8 Borkenkäferbefall unversehrt gebliebene 24 bis bis 39 Altholzbestände..stehendes T otholz N : Durch Borkenkäferbefall abgestorbene Altholzbestände. "belassene Windwurffläche": Naturbelassene Windwurffläche ohne direkte menschliche Einflussnahme, entstanden im November Bäume) gleicht, hat die Durchmesserverteilung der abgestorbenen, aber noch stehenden Bäume dagegen einen konkaven (exponentiellen) Verlauf (Abbildung 15). Dies ist naheliegend, da nach einer Phase hoher Mortalität (beginnend bei den Samen, dann bei den Keimlingen und Sämlingen Lebende Bäume 40 r ~ bis bis bis bis bis bis 20 Tab. 6 Bestandssituation in den verschiedenen Transektabschn itten. N Neben der kartografischen Aufbereitung der umfangreichen Messungen auf dem Transekt vermitteln die berechneten Bestandskennwerte einen Eindruck des Waldaufbaus (Dichte, Dimensionen, horizontale und vertikale Struktur). Es wurden insgesamt 109 lebende, 63 tote und 182 liegende Bäume ab Derbholzstärke (7 cm Durchmesser) aufgenommen. Die Bäume im geschlossenen Altholz sind über alle Durchmessserklassen verteilt. Während bei den lebenden Bäumen die Durchmesserverteilung in etwa einer Gauß'schen Glockenkurve (Normalverteilung) mit einem Maximum in den Klassen 3 und 4 (zwischen 25 und 44 cm BHD; 60 % der o Durchmesserklassen (BHD) Abgestorbene Bäume 40 r ~ 30 :c ~ 20 «10 o Durchmesserklassen (BHD) Abb.15 Durchmesserverteilung des lebenden und toten Derbholzbestandes. 65
 durch Borkenkäferbefall in den späten 80er Jahren abgetötet wurden.")

65 ~ usw.) die Überlebenden eine hohe Lebenserwartung haben: so dürften die schwachen Individuen vor allem auf Grund von intraspezifischer, d. h. innerartlicher Konkurrenz um Licht, Wasser und Nährstoffe abgestorben sein, während die stärkeren (und alten) durch Borkenkäferbefall in den späten 80er Jahren abgetötet wurden. Abb. 16 Bäume mit langen, schlanken Kronen sind hervorragend an die rauen Lebensbedingungen in den Hochlagen des Bayerischen Waldes angepasst. Tab. 7 Charakterisierung des geschlossenen Altholzes anhand gebräuchlicher Bestandesmittelstämme. Brusthöhendurchmesser [ern] Gesamthöhe [m] Die Berechnung der Durchmesserdifferenzierung TD nach v. GADOW und FUELDNER (1992) ergab Werte von 0,22 (Probequadrate 1 bis 9) und 0,33 (Probequadrate 24 bis 39). Dies sind lediglich 22 bzw. 33 % des maximal möglichen Wertes, d. h. die Brusthöhendurchmesser der Bäume variieren nur in einem sehr engen Rahmen. Eindrucksvoll sind die gemessenen Baumhöhen. Ab einem Brusthöhendurchmesser von 35 cm erreichen die meisten Bäume 25 bis 30 m Höhe. Die maximale Höhe von 36 m wurde von einem außerordentlich imposanten, 1992 bereits abgestorbenen Baum in der Nähe der Fleischhüttenseige auf Probequadrat 35 erreicht, dessen BHD 90 cm betrug! Die überwiegende Anzahl der Bäume weist lange und schlanke Kronen auf. Besonders auffällig ist dies in den Probequadraten 24 bis 39 (siehe auch Bestandsaufrisse in der Anlage!); dort reicht die Krone einzelner Bäume bis nahe an den Boden herab (Abbildung 16). Ein Phänomen, das sich nur sehr aufwendig fassen und darstellen lässt, aber für die Bergfichtenwälder auf Grund der Baumverteilung typisch ist, sind die teilweise extrem asymmetrischen horizontalen und vertikalen Kronenausformungen. Grundflächen- Oberhöhenstamm T mittelstamm ~ 39,2 55,S I 25,7 29,8 Sehlankheitsgrad (Höhe/BHD; "H/D-Verhältnis"): I I' Kronendurchmesser [m] 3,56 4,81 Kronenansatzhöhe [m] 10,8 11,7 Kronenanteil [%] ~ Plumpheitsgrad der Krone (KD / KL) 0,24 - I~ 0,26 Zur Beschreibung der durchschnittlichen Baumdimensionen wurden Grundflächenmittel- und Oberhöhenstamm verwendet (Tabelle 7). Die berechneten Mittelwerte machen deutlich, dass die vom Sturm und dem nachfolgenden Borkenkäferbefall unversehrt gebliebenen Bäume im Untersuchungsgebiet bis 1992 beträchtliche Dimensionen (BHD und Höhe) erreicht hatten. Sie besaßen lange, schlanke Kronen (Verhältnis Kronendurchmesser zu -länge etwa 1 : 4) und waren auf Grund ihres geringen Schlankheitsgrades (H/D-Verhältnis < 70) widerstandsfähig gegenüber abiotischen Belastungen wie v. a. Schnee und Wind (ROTIMANN 1986). Die ermittelten Stammzahlen, Holzvorräte und der durchschnittliche Vermoderungsgrad des Totholzes auf dem Transekt im Jahr 1992 sind in Tabelle 8 zusammengefasst. Die 1992 noch geschlossenen Altholzbestände waren stammzahlreich; die Bäume waren aber überwiegend in z. T. recht dichten Gruppen ("Rotten") zusammengedrängt; es wechselten also dichte mit sehr lockeren Bestandesteilen (vgl. An- 66
.")

![Einheit A B Stammzahl (lebend): N/ha 428 0 Stammzahl (stehend tot): N/ha 148 193 liegende Stämme und Stammteile: N/ha 288 786 Grundfläche (lebend): G/ha [m 2 ] 52,3 0, Vorrat (lebend): V/ha [VfmD]](/docs-images/61/46190100/images/66-2.png "632 0 Vorrat (stehend, tot): V/ha [VfmD] 116 293 Vorrat (liegend, tot): V/ha [VfmD] 68 414 Vorrat gesamt: V/ha [VfmD] 816 707 I Anteil der Fichte: NOfo = GOfo = VOfo 100 100 Totholzanteil: V(tot)!")
66 lagen 3a und 3b). Das berechnete Holzvolumen von 816 Vorratsfestmetern Derbholz je Hektar (VfmD/ha). ist für diese Höhenlage überraschend hoch (im Rahmen der Waldinventur 1996 wurden z. B. für die gesamten Hochlagen des Nationalparks Bayerischer Wald im Durchschnitt 406 VfmD/ha ermittelt; NUESSLEIN 1996). Es hatte sich seit dem letzten forstwirtschaftlichen Eingriff im Jahr 1969 bereits eine beträchtliche Menge an liegendem und stehendem Totholz - mit 184 VfmD/ha knapp ein Viertel des gesamten Vo lumens - angesammelt. Einheit A B Stammzahl (lebend): N/ha Stammzahl (stehend tot): N/ha liegende Stämme und Stammteile: N/ha Grundfläche (lebend): G/ha [m 2 ] 52,3 0, Vorrat (lebend): V/ha [VfmD] Vorrat (stehend, tot): V/ha [VfmD] Vorrat (liegend, tot): V/ha [VfmD] Vorrat gesamt: V/ha [VfmD] I Anteil der Fichte: NOfo = GOfo = VOfo Totholzanteil: V(tot)!V(ges) [Ofo] I Mittlerer Zersetzungsgrad (stehend) : 1,1 1,0 Mittlerer Zersetzungsgrad (liegend): 2,1 1,1 Die Stammzahlen im Totholzsaum und auf der belassenen Windwurffläche waren etwas höher als im angrenzenden Altholz, was aber v. a. methodische Ursachen hat: waren liegende Stämme in mehrere Teile zerbrochen, so wurden die einzelnen Stammstücke erfasst. Außerdem wurden herabgestürzte Gipfelstücke von stehenden toten Bäumen aufgenommen, sobald sie mindestens 7 cm stark waren. Auch auf der Windwurffläche waren die Bäume vor dem Sturm in Rotten angeordnet, worauf die Verteilung der liegenden Stämme, aber auch die sehr unterschiedliche Anzahl aufgeklappter Wurzelteller in den einzelnen Probequadraten deutlich hinweist (Anhang 3a und 3b; Kapitel 4.2). Während in einigen Probequadraten nur wenige umgestürzte Bäume liegen, sind es z. B. im PO 15 sogar 19 Stämme bzw. Stammteile. Der überwiegende Teil der geworfenen Bäume liegt mit den Kronen in östlicher bis nordöstlicher Richtun g. Dies ist verständlich, da die Herbststürme im November 1984 aus westlicher Richtung kommend den Nationalpark überquerten. einzelnen Probequadraten zu beobachten sind - (Abbildung 18), ist der Unterschied von ca. 100 VfmD/ha (1 3 Ofo) v. a. durch die lange und schmale Form der Untersuchungsfläche zu erklären. Das liegende Totholz auf der Windwurffläche war zum überwiegenden Teil noch kaum ve rmodert, im Gegensatz zum liegenden Toth olz im intakten Altholzbestand. Anscheinend sind dort die Bäume schon früher umgestürzt. Zur Analyse der vertikalen Bestandesstruktur (Schichtung) wurde der Artprofilindex na ch PRUZSCH (1996) verwendet. Für die Probequadrate 1 bis 9 weist der Wert von 0,68 auf ei ne relativ geringe Vertikalstruktur hin, wohin gegen der Art- Tab. 8 Waldwachstumskundliche Bestandskennwerte. A: geschlossenes Altholz; B: Totholzsaum und belassene Windwurffläche. Abb.77 Aufgrund des Verteilungsmusters der liegenden Stämme kann auf die Rottenstruktur des Bestandes vor dem Sturm geschlossen werden. Die Holzvorräte auf der Windwurffläche und dem umgebenden Totholzsaum entsprachen etwa jenen der angrenzenden Altholzteile. Da das Holz nicht gleichmäßig über den Transekt verteilt ist, sondern teilweise große Schwankungen zwischen 67
 E ::s (5 10 ~ (5 :I: 5 30 r--------------------------------------------------------------, o 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33 35 37 39 Probequadrate")
.")
67 Abb.18 Holzvorrat auf den einzelnen Probequadraten. 25 '4J "0 c: Ci: 20 E E ~ 15 c: CI) E ::s (5 10 ~ (5 :I: 5 30 r , o Probequadrate lebende Bäume L...-_... 1stehendes Totholz liegendes Totholz Mittelwert profilindex für den letzten Transektabschnitt (0,99) eine sehr hohe vertikale Strukturierung kennzeichnet! Dieses Ergebnis wird auch durch die Bestandsaufrisse sehr anschaulich wiedergegeben (Anlagen 3 a und 3 b). Bei den Wiederholungsaufnahmen im Sommer 1997 wurden bei den alten Bäumen keine BHDoder Höhenmessungen vorgenommen, da die Veränderungen in fünf Jahren vermutlich gering sind. Waren jedoch Bäume zwischen den beiden Aufnahmejahren abgestorben, abgebrochen oder umgestürzt, so wurde dies festgehalten. Ebenso wurde aktueller Borkenkäferbefall bei der Wiederholungsaufnahme registriert. Auf den Probequadraten 1 bis 9 sind zwischen Erst- und Wiederholungsaufnahme acht Bäume (22 Ofo) abgestorben, 13 (35 0/0) waren aktuell vom Buchdrucker befallen. Drei bereits 1992 tote Bäume mit einem BHD zwischen 48 und 61 cm (50 0/0) sind in Höhen zwischen 1,8 und 3,2 m abgebrochen. Außerdem wurden elf neue Stamm- bzw. Gipfelstücke erfasst, die von außerhalb in die Aufnahmeflächen gestürzt sind. Im Altholzbestand in der zweiten Hälfte des Transekts sind acht Bäume (11 Ofo) in der Zwischenzeit abgestorben, drei (4 0/0) waren aktuell von Borkenkäfern befallen und vier (13 0/0) in Höhen zwischen 2 und 3 m abgebrochen. Es lagen acht neue Stammstücke auf den Probequadraten. Im Totholzsaum, entsprechend der Abgrenzung von 1992, waren zwei Bäume (8 0/0) in einer Höhe von ca. 2 m abgebrochen und 16 neue Stammund Gipfelstücke von außerhalb hereingestürzt. In der belassenen Windwurffläche traten dagegen keine auffälligen Veränderungen am Derbholzbestand ein. Die Vermoderung des Totholzes ist in fünf Jahren kaum fortgeschritten (soweit dies durch eine visuelle Ansprache beurteilt werden kann). Anhand der stichprobenartigen Entnahme von 47 Bohrkernen in 1,30 m Höhe von Bäumen auf dem gesamten Transekt bzw. in dessen unmittelbarer Nachbarschaft konnten die Baumalter durch Auszählen der Jahrringe näherungsweise ermittelt werden. Nicht berücksichtigt wurden Jahrring- 68
.")
 > 200 BHD in cm ermittelte Alter Anzahl von... bis von.")

 sehr unterschiedlich. Tab. 9 Ermittelte Altersspannen in den verschiedenen Durchmesserklassen.")
68 ausfälle, wie sie nach KAATZ (1987) bei Fichten in den Hochlagen des Nationalparks Bayerischer Wald in den letzten Jahrzehnten gehäuft beobachtet wurden, so dass die aus der Zählung ermittelten Baumalter als untere Schätzwerte der tatsächlichen angesehen werden müssen. Es wurden zwischen 80 und 252 Jahrringe gezählt (Abbildung 19). Berücksichtigt man, dass in dieser Höhenlage Jahrzehnte verstreichen, bis die heranwachsenden Bäume die Messhöhe von 1,30 m erreicht haben, so sind die ermittelten Alter sehr beachtlich! Alter (Jahre) > 200 BHD in cm ermittelte Alter Anzahl von... bis von... bis Bäume , , ,- - ~ 65 (- 90) bereits einen BHD von 66 cm erreicht hatte, bei Baum Nr. 2 in der Nähe von PO 3 mit einem BHD von 39 cm wurden dagegen 192 Jahrringe gezählt! Im Gegensatz zur geringen Durchmesserdifferenzierung ist die Altersstruktur auch innerhalb einzelner Baumkollektive ("Rotten": Beispiel P031, Nr. 2: 198Jahre; Nr. 3: 178Jahre und Nr. 4: 136 Jahre) sehr unterschiedlich. Tab. 9 Ermittelte Altersspannen in den verschiedenen Durchmesserklassen < 100 o Anzahl Bäume Es wird darüber hinaus deutlich, dass eine Altersschätzung über die Dimension eines Baumes in den Bergfichtenwäldern sehr fragwürdig ist (Tabelle 9): So wurden z. B. bei Baum Nr. 4 auf PO 31 lediglich 136 Jahrringe gezählt, obwohl er Anhand der Bohrspäne einzelner Individuen wurde deren Wachstumsverlauf rekonstruiert (Abbildung 20). Dabei erlaubte der Bohrspan Nr. 12 von einer außerordentlich eindrucksvollen Fichte etwas außerhalb von PO 35 einen Rückblick etwa bis ins Jahr 1760! Auffallend ist die deutliche Zunahme der Jahrringbreiten (10-Jahres-Zuwachs) um 1880, in einer Zeit also, die geprägt war vom "Großen Sturm" (1868/70) und der nachfolgenden Borkenkäfer-Gradation. Abb.19 Altersverteilung bei den 47 gemessenen Bäumen. 40 -~ ~ Abb.20 Wachstumsverlauf ausgewählter Fichten im Untersuchungsgebiet. E 30-5 VI ~ C.J cu ~ ~ 20 - ~ ~ cu 7 c: ~ ~ BSPll (BHD = 66) -- BSP12 (BHD = 72) --BSP36 (BHD = 90) --BSP37 (BHD = 60) --BSP44 (BHD = 45) -- BSP45 (BHD = 53) - Mittlerer Zuwachs (berechnet aus dem Wachstumsverlauf von 47 Bäumen) 69
 Das verrottende Holz von umgestürzten")


.")
69 Abb. 21 (0. I.) Das verrottende Holz von umgestürzten Bäumen... Abb. 22 (u. I.)... oder von Baumstümpfen bildet ein günstiges Keimbett für die Naturverjüngung in Bergfichtenwäldern. Abb. 23 (0. r.) Durch Windwurf... Abb. 24 (u. r.)... und Borkenkäferbefall entstehen neue verjüngungsfreundliche Kleinstandorte. 70



. d. h.")
70 4.2 Kleinstrukturen 15 -r ~ ::c cu N c: « Probequadrate o frisch tot D beginnende Zersetzung Alte Baumstümpfe, liegendes Moderholz oder Wurzelteller sind wichtige Elemente im Bergfichtenwaid. Sie tragen ganz wesentlich zur Strukturvielfalt in den scheinbar sehr monotonen Wäldern bei und bilden offensichtlich wichtige Trittsteine für die Etablierung der Naturverjüngung (Abbildungen 21-24). Darüber hinaus kann über eine Erfassung der alten Baumstümpfe die horizontale Struktur der Waldbestände in der Vergangenheit analysiert werden. Einen Überblick über Anzahl, Durchmesserverteilung und Zersetzungsgrad (Ansprache in vier Stufen in Anlehnung an ALBRECHT 1990) der Baumstümpfe vermitteln die Abbildungen 25 und 26. fortgesch rittene Zersetzu ng stark vermodert Im Durchschnitt wurden vier Baumstümpfe je Probequadrat gefunden. Beim überwiegenden Teil - etwa zwei Drittel- handelt es sich um schwache Baumstümpfe der Durchmesserklassen eins bis drei, während nur noch fünf starke Baumstümpfe mit einem Durchmesser von über 54 cm gefunden wurden. Der überwiegende Teil ist bereits stark bis sehr stark vermodert (durchschnittlicher Zersetzungsgrad 2,9). d. h. dass die letzten forstwirtschaftlichen Eingriffe tatsächlich schon viele Jahre zurückliegen. Die wenigen Exemplare der Stufe eins rühren daher, dass in den letzten Jahren immer wieder abgestorbene Bäume umgebrochen sind. Abb. 25 Anzahl und Zersetzungsgrad der Baumstümpfe in den einzelnen Probequadraten. 80 -, , 80 -~ ~ 60 ::c ~ 40 «20 ::c 60 ~ 40 c: «20 o Durchmesserklassen Zersetzungsgrad Abb.26 Durchmesserverteilung und Zersetzungsgrad der Baumstümpfe M


.")
71 Abb.27 Flächenmäßige Bedeutung verschiedener Kleinstrukturen Baumstümpfe + Umgebung 3+4 Liegendes Totholz + Umgebung 5 Stammfußbereich toter Bäume 6 Durch herausgerissene Wurzeln freigelegter Boden 7 Fläche ohne Kleinstrukturen Die Anzahl aufgeklappter Wurzelteller auf dem belassenen Windwurf schwankt zwischen einem und sechs Stück je Probequadrat, im Durchschnitt waren es drei. Da die Krautschicht in den Bergfichtenwäldern meist sehr dicht und üppig entwickelt ist (Kap. 4.3 und HEURICH 2001 b) und Baumstümpfe, liegendes Totholz und der durch die herausgehebelten Wurzeln umgestürzter Bäume freigelegte Mineralbo den häufig die einzigen, von krautiger Konkurrenzvegetation freien oder nur schütter bewachsenen Bereiche sind, wurde deren flächenmäßige Bedeutung auf dem Transekt berechnet (Abbildung 27). Im geschlossenen Altholz finden sich zwar bereits alle ausgeschiedenen Kleinstrukturen, deren flächenmäßige Bedeutung ist aber mit durchschnittlich 11 % Gesamtdeckung noch sehr gering. Die Schnitt- bzw. Bruchflächen der Baumstümpfe nehmen nur 0,3 % der Fläche ein. Berücksichtigt man jedoch deren unmittelbare Umgebung bis zum Abstand des jeweiligen Stockdurchmessers ("Wirkungsfläche"; Kap ), so erhöht sich der Flächenanteil auf etwa 2 bis 3 0/0. Das liegende Totholz bedeckt im Durchschnitt ca. 3 0 /0, einzelne Probequadrate weisen bereits bis zu 12 % Totholzbedeckung auf. Berücksichtigt man auch hier die Umgebung, so verdoppelt sich die Wirkungsfläche. Der überwiegende Teil der Baumstümpfe und der liegenden Stämme ist bereits so weit vermodert, dass er ein geeignetes Keimsubstrat für Baum- verjüngung darstellt. Einzelne Stämme sind schon dicht mit jungen Fichten bewachsen (siehe Stammfußkarte, PO 35 und 36!). In den durch Borkenkäferbefall abgestorbenen Bestandesteilen beträgt der Anteil der Kleinstrukturen im Mittel bereits 30 % an der Gesamtfläche. Neben den innerhalb weniger Jahre ab- bzw. umgebrochenen toten Bäume sind es vor allem die Stammfüße der abgestorbenen Bäume, um die sich herabfallende Nadeln, Feinreisig und Rinde sammeln und die so zu einer Veränderung im Vegetationsmosaik führen. In der Windwurffläche hat sich die Situation gravierend verändert, nur mehr 26 % der Fläche sind ohne besondere Kleinstrukturen. Vor allem das liegende Totholz und die aufgeklappten Wurzelteller tragen zu einer enormen Strukturvielfalt bei. In einzelnen Probequadraten wurde eine Totholzbedeckung bis zu 50% berechnet. Jedoch haben dort viele geworfene Stämme bis heute noch keinen Bodenkontakt, sondern hängen, ineinander verkeilt, bis zu 2 m hoch über dem Erdboden. Die Holzzersetzung und damit verbunden die Sackung wird noch einige Jahre in Anspruch nehmen. Trotz der geringen Vermoderung des Lagerholzes wurde aber auch hier schon Rannenverjüngung bei der Wiederholungsaufnahme 1997 beobachtet, wenngleich deren Wuchs noch sehr kümmerlich war! Vor allem an den Wurzelanläufen im Übergangsbereich zwischen Wurzelteller und Baumstamm scheint eine Ansamung sehr rasch möglich zu sein. geschlossenes Altholz stehendes Totholz belassener Windwurf


 und die Waid-Hainsimse (Luzula sylvatica).")

72 4.3 Kraut- und Moosschicht Die Ergebnisse der Vegetationsaufnahmen sind in den Vegetationstabellen im Anhang 2a und 2b zusammengestellt. Die vorgefundenen Moose sind in diesen Tabellen noch nicht enthalten, da die Artbestimmung noch redigiert werden muss. Bei der Erstaufnahme 1992 wurden 38 Arten der Farn- und Blütenpflanzen festgestellt, bei der Wiederholungsaufnahme waren es lediglich 34. Am Aufbau der Krautschicht sind meist nur wenige Arten beteiligt. Es sind dies v. a. die Süßgräser Berg-Reitgras (Calamagrostis villosa) und Drahtschmiele (Deschampsia f1exuosal, der Berg-Frauenfarn (Athyrium distentifolium) und Breitblättrige Dornfarn (Dryopteris dilatata), pie Heidelbeere (Vaccinium myrtillus) und die Waid-Hainsimse (Luzula sylvatica). Daneben kommen nur noch wenige Arten mit hoher Stetigkeit, aber ge- ringer Flächendeckung vor: Sauerklee (Oxalis solche mit hygromorphem Bau ein (d. h. sie benötigen hohe Luftfeuchtigkeit, wie sie im Schatten oder Halbschatten eines Waldes vorhanden ist): das Zwei blättrige Schattenblümchen (Maianthemum bifolium), der Grüne Alpenlattich und die Berg-Troddelblume (Soldanella montanal, aber auch die Moose.... ] Cl ~ 40 Vl >< Cl 30 ~ tj CI.I Cl ~ 20 E 'E 10 acetosella) oder Grüner Alpenlattich (Homogyne 0 alpina). Dieser Sachverhalt ist in Abbildung 28 dargestellt. Dabei wurde neben der mittleren Deckung auch die relative Stetigkeit, d. h. die Regelmäßigkeit des Vorkommens einer Art in den Aufnahmeflächen berücksichtigt, indem beide Werte miteinander multipliziert wurden ("Bedeutungs-" oder "I mportanzwert"). Vergleicht man diesen "Importanzwert" der Arten in den verschiedenen Bestandssituationen, so wird deutlich, dass es eindeutige "Gewinner" in der Folge von Störungen wie Windwurf und Borkenkäferbefall gibt: das Berg-Reitgras, das bereits in den Bergfichtenwäldern überaus reichlich vorhanden ist und sich auf Grund der Auflichtung noch üppiger entfalten kann sowie Himbeere (Rubus idaeus) und Schmalblättriges Weidenröschen (Epilobium angustifolium) als "Neueinwanderer" auf der Windwurffläche. Die "Verliererrolle" nehmen v. a. konkurrenzschwache Arten oder... ] Cl Abb.28 Dominante Arten in der Krautschicht. 60 r , 50 '"ß Vl >< Cl geschlossenes Altholz , ",------~.,~-,-'-,,-~-,--f_-- ~ tj CI.I Cl ~ 20 E 'E 10 o gesch lossenes Altholz Berg-Reitgras Drahtschmiele o Berg-Troddelblume stehendes Totholz stehendes Totholz Heidelbeere Berg-Frauenfarn D Himbeere belassener Windwurf belassener Windwurf. Wald-Hainsimse D Grüner Alpenlattich Schmalblättriges Weidenröschen 73


.")
73 Krautschicht 20 r ~ Abb.29 Entwicklung der mittleren Artenzahlen in der Kraut- und Moosschicht. Abb.30 Entwicklung der mittleren Deckung von Kraut- und Moosschicht. o c 15 cu ~ ::i:: 10 ca N C oe( 5 gesch lossenes Altholz stehendes Totholz Moosschicht belassener Windwurf 20 r ~======~ o l 80 Cl c ~ 60 C.I cu Cl ~ 40 cu E 'E 20 gesch lossenes Altholz stehendes Totholz Krautschicht belassener Windwurf 100.., , o l 80 Cl C ~ 60 C.I cu Cl ~ 40 cu E 'E 20 geschlossenes Altholz stehendes Totholz Moosschicht belassener Windwurf 100.r , o geschlossenes Altholz stehendes Totholz belassener Windwurf Insgesamt betrachtet kommt es in der Krautschicht nur zu geringen, in der Moosschicht dagegen zu beträchtlichen Veränderungen (Abbildungen 29 und 30). Während die mittlere Artenzahl bei den krautigen Pflanzen in den verschiedenen Bestandssituationen annähernd gleich geblieben ist bei etwas höherer durchschnittlicher Flächendeckung, war bei den Moosen ein deutlicher Rückgang außerhalb des geschlossenen Altholzes zu beobachten: die mittlere Artenzahl hatte um ca. 40 0/0, die mittlere Deckung sogar um ca. 70 Ofo abgenommen. Betroffen von den veränderten Umweltbedingungen bzw. von der zunehmenden Konkurrenz durch die Gefäßpflanzen waren vor allem die zierlichen, meist in sehr kleinen Gruppen wachsenden foliosen Lebermoose, wie z. B. die Spitzenbartmoose (Barbilophozia floerkei und B. Iycopodioides), das Haarkelchmoos (Trichocolea tomentella) oder das Spatenmoos (Scapania nemorea). Diese Arten wurden im stehenden Totholz und auf der belassenen Windwurffläche nicht mehr gefunden. Lediglich kräftige Laubmoose, wie Schönes und Gemeines Frauenhaarmoos (Polytrichum formosum und P. commune), Schönes Kranzmoos (Rhytidiadelphus loreus) und Besengabelzahnmoos (Dicranum scoparium) sowie das Torfmoos Sphagnum girgensohnii waren in den Störungsflächen noch häufiger vertreten. Es wurde überprüft, ob die Veränderungen in der Zusammensetzung der Krautschicht auch in den mittleren Zeigerwerten für die Standortsfaktoren Licht (L), Temperatur (T), Kontinentalität (K), Feuchtigkeit (F), Reaktion (ph-wert, R) und Stickstoffversorgung (N) (ELLEN BERG 1991) sichtbar werden. Die Zeigerwert-Skala reicht von 1 (geringes Angebot bei L, T, F, N bzw. ozeanisch bei K und "sehr sauer" bei R) bis 9 (reichliches Angebot bzw. kontinental bei Kund basen- bzw. kalkreich bei R). 74


 wurden für die Vegetationsaufnahmen im Übergangsbereich zum stehenden Totholz errechnet.")
.")
 und Kontinentalität (K) signifikant (p < 0,01), eine steigende Tendenz ist beim Faktor Licht (L) zu erkennen.")
74 c::l 9 CI! e LI I---r-- t? 1-- ~ i r I - - r-~-- ~l-< r ' I.~"T I J-t-- -I--~ -.~ i I! I - ~----~-- -i--i---,., --LL 3 e Q.J 2..><: ; - u ß 1! Q dunkel L hell 9 -~~~--~~~--~~~ -5 I. ~ 8._ ~ " ~- 1 z I---t-----t-..."~+_~-t I I-- - ~ ",, z ;. 4.~~~'~-ry-----f----~----~--~ r~ +1 --! ! I I ~ ;- Z I i sauer R basisch ~ e Q.J e '.p e 0..><: ::.:: 5.s;;; ~ 'e: CI! 2 Q.J N 0 1 kalt I T warm Die mittleren Zeigerwerte der einzelnen Vegetationsaufnahmen im geschlossenen Altholz ergaben für den Faktor Licht Werte zwischen 4 und 5, diese weisen auf Halbschatten-Bedingungen hin. Beim Faktor Feuchte liegen die Mittelwerte um 6, d. h. es handelt sich um Pflanzenbestände frischer bis feuchter Standorte mit ausgeglichener, guter Wasserversorgung. Das Stickstoffangebot auf den sauren bis stark sauren Böden ist dürftig, die höchsten Werte (4,3 und 4,7) wurden für die Vegetationsaufnahmen im Übergangsbereich zum stehenden Totholz errechnet. Die niedrigen Temperatur- und Kontinentalitätszahlen bestätigen die hochmontane bis subalpine Lage mit subozeanischer Klimaausprägung (Abbildung 31). Die VeränderUngen in den mittleren Zeigerwerten liegen im belassenen Windwurf und stehenden Totholz bei maximal 0,6 Zeigerwerteinheiten (Abbildung 32). Im Jahr 1992 unterscheiden sich belassener Windwurf und geschlossenes Altholz nur bzgl. der Faktoren Stickstoff-Versorgung (N) und Kontinentalität (K) signifikant (p < 0,01), eine steigende Tendenz ist beim Faktor Licht (L) zu erkennen. Beim Vergleich der mittleren Zeigerwerte zwischen stehendem Totholz und intaktem Altholz konnte kein signifikanter Unterschied nachgewiesen werden, wenngleich auch hier eine Zunahme bei den N- und K-Werten zu beobachten ist. Ebenso unterscheiden sich 1997 lediglich Windwurf und Altholz bzgl. der N- und K-Werte signifikant, eine steigende Tendenz ist nun aber auch bei den T - und F-Werten zu erkennen. Dies lässt eine stärkere Differenzierung zwischen diesen beiden Entwicklungsstadien mit zunehmender Zeit vermuten. t:: Q.J ~ Q.J Cl.~ N Abb.31 Mittlere Zeigerwerte der Vegetationsaufnahmen in den geschlossenen Altholzbeständen. Abb. 32 Entwicklung der mittleren Zeigerwerte in den verschiedenen Bestandssituationen. 7 -~ ~ ~ ~ Q - Q.-a 4 --~--"~"".!:--~±==~=f=;;;; ~ ~ ~ ~ ~ geschlossenes Altholz stehendes Totholz belassener Windwurf Q Licht - Temperatur... Kontinentalität -Feuchte -+- Reaktion... Stickstoffversorgung 75
.")




75 Abb. 33 (o.) Während die Soldanelle... Abb. 34 (m. I.)... und der Sprossende Bärlapp durch die Umweltveränderungen, v. a. aber durch die zunehmende Konkurrenz des Berg Reitgrases in Bedrängnis geraten werden,... Abb. 35 (u. I.)... profitieren Himbeere... Abb. 36 (u. r.)... und Weidenröschen von den Bodenöffnungen und vom erhöhten Licht- und Nährstoffangebot. 76
 je Probequadrat - dies entspricht 13.")
 mit 212 Fichten (davon 209 ~ 20 cm) und 11 Vogelbeeren (alle über 20")


76 4.4 Naturverjüngung Dichte Auf dem gesamten Transekt wurden 1992 insgesamt Pflanzen ~ 20 cm und 749 Pflanzen < 20 cm gezählt. Die Bestandsaufnahme erbrachte beeindruckende Ergebnisse: So wurden bereits unter dem Schirm des intakten Altholzes bis zu 133 Fichten (47 < 20 cm, 86 ~ 20 cm) je Probequadrat - dies entspricht Pflanzen je Hektar! - und bis zu 58 Vogelbeeren (7 < 20 cm, 51 ~ 20 cm) je Probequadrat, also je Hektar gezählt. Im Totholzsaum und im belassenen Windwurf wurden diese Zahlen sogar übertroffen: die maximale Dichte wurde im Totholzsaum (PQ 12) mit 212 Fichten (davon 209 ~ 20 cm) und 11 Vogelbeeren (alle über 20 cm) festgestellt ~ 150 <ti N C <{ :c <ti N C <{ 100 geschlossenes stehendes belassener Altholz Totholz Windwurf Abb. 37 Verjüngungsdichte in den verschiedenen Bestandssituationen in den Jahren 1992 und o : Extremwerte x : Ausreisser Bei den Pflanzen mit mindestens 20 cm Größe waren es im Durchschnitt der Probequadrate 22 Fichten und 8 Vogelbeeren im geschlossenen Altholz, 85 Fichten und 7 Vogelbeeren im stehenden Totholz sowie 67 Fichten und 5 Vogelbeeren auf der belassenen Windwurffläche. Selbst in der belassenen Windwurffläche spielen Pionierarten wie Birke, Weide oder Aspe eine unbedeutende Rolle (es wurden lediglich drei Individuen gefunden!). Da die Verteilung bzw. Dichte in den einzelnen Probequadraten sehr unterschiedlich ist (siehe Stammfußkarte im Anhang), wurde die Häufigkeitsverteilung der Verjüngung in Form sogenannter "Box-Plots" dargestellt (Abbildung 37). Dabei verkörpert die "Box" 50 % der Aufnahmeflächen je Entwicklungsstadium, 25 % liegen zwischen Minimalwert und erstem Quartil (unterer Rand der Box). 25 % befinden sich zwischen drittem Quartil (oberer Rand) und Maximalwert. Der Median (Zentralwert) ist derjenige Messwert, unter- bzw. oberhalb dessen jeweils die Hälfte der Messwerte liegen geschlossenes stehendes belassener Altholz Totholz Windwurf Fichte Vogelbeere Im geschlossenen Altholz wurde 1992 lediglich auf vier von 25 Aufnahmeflächen überhaupt keine Naturverjüngung mit mindestens 20 cm Größe gefunden;jedoch wuchsen in zwei Quadraten bereits Bäume unter 20 cm. Im stehenden Totholz und im Windwurfwaren dagegen bereits alle Probequadrate besetzt. Abb. 38 In den Totholzflächen hat sich bereits wenige Jahre nach Sturm und Borkenkäferbefall die Naturverjüngung - überwiegend Fichten - üppig entwickelt. 77
.")
 vor allem im geschlossenen Wald, obwohl die nächsten Samenbäume relativ weit entfernt sind.")
77 Abb. 39 Baumartenanteile bei der Erstaufnahme Abb. 40 Verteilung der Baumverjüngung auf verschiedene Kleinstandorte im Jahr ~ 60.~ 40 «20 o geschlossenes Altholz Fichte stehendes Totholz. Vogelbeere belassener Windwurf Die Fichte ist die häufigste Baumart (Abbildung 39) betrug ihr Anteil im geschlossenen Altholz 73 % an der gesamten Naturverjüngung, in den Totholzflächen ist dieser sogar auf über 90 Ofo angestiegen. Interessant ist der hohe Anteil der Vogelbeere (27 Ofo) vor allem im geschlossenen Wald, obwohl die nächsten Samenbäume relativ weit entfernt sind. Dies deutet auf eine effektive endozoochore Verbreitung der für viele Tierarten (v. a. Vögel, aber auch Baummarder und Schlafmäuse) offenbar sehr attraktiven Früchte hin. Nach der Verdauung werden die Samen, die selbst unverdaulich sind, mit dem Kot wieder ausgeschieden. Damit erhalten sie zugleich eine kleine Startdüngung! Birken, Weiden oder Aspen spielen auch in den Totholzflächen zahlenmäßig keine Rolle, lediglich in der Windwurffläche nehmen sie einen Anteil von 0,6 Ofo ein. Bis 1997 waren 92 Pflanzen abgestorben oder wurden nicht mehr aufgefunden. Dennoch hat die Gesamtzahl der Pflanzen ~ 20 cm noch einmal um ca. 7 Ofo auf 1987 Individuen zugenommen, die Baumartenanteile an der gesamten Naturverjüngung haben sich nur wenig verändert: Der Anteil der Fichte hat im geschlossenen Altholz um 3 Ofo zu-, der der Vogelbeere entsprechend abgenommen.lm stehenden Totholz ist dagegen dervogelbeerenanteil um 4 Ofo angestiegen. In der belassenen Windwurffläche blieb die Baumartenzusammensetzung nahezu gleich. Lediglich auf drei Probequadraten im intakten Altholz konnte noch keine Naturverjüngung gefunden werden. Auch bei der Wiederholungsaufnahme wurden, neben den beiden Birken und der Weide von der Erstaufnahme, keine weiteren Pioniere gefunden; nur eine Buche hatte sich in diese Höhenlage "verirrt" Keimungssubstrat (Kleinstandorte) Es gibt viele Hinweise aus der Literatur, aber auch aus dem Erfahrungsschatz von Förstern, die mit Bergfichtenwäldern vertraut sind, dass vermoderndes Holz in Form alter Baumstümpfe und liegender Stämme (Abbildung 42) ein sehr günstiges Keimbett für Naturverjüngung bildet ( sog. "Rannenverjüngung"). Um dessen Bedeutung Fichte ,----,--...,...--_ ,...-.,..., 80 ~ 60 'Cij ~ 40 «20 o gesch lossenes Altholz stehendes Totholz belassener Windwurf Vogelbeere , ,...-..,.-..., 80 B s:::: 40 «20 geschlossenes Altholz Moderholz stehendes Totholz Wurzelteller belassener Windwurf Sonstige 78


.")
78 quantifizieren zu können, wurde bei der Verjüngungsaufnahme registriert, ob Bäume auf diesem Substrat wachsen. Da nur zweifelsfrei erkennbares Moderholz erfasst wurde, sind die Verjüngungszahlen auf diesem Keimsubstrat als minimaler Schätzwert der tatsächlichen Verjüngungszah len auf Moderholz anzusehen. Neben dem Moderholz wurden auch die Kleinstandorte Wurzelteller und Fels - der aber auf dem Transekt keine Rolle spielt - erfasst. Es zeigte sich tatsächlich, dass für die Fichte in dieser Höhenlage vermoderndes Holz in Form alter Baumstümpfe oder umgestürzter Bäume das bedeutendste Keimsubstrat bildet. Bei der Erstaufnahme 1992 wurden im geschlossenen Altholz 53 % des Fichtenjungwuchses auf diesem Substrat erfasst (Abbildung 40), 1997 war dort der ~ ~ E I:: «~ ~ E I:: « geschlossenes Altholz Fichte Vogelbeere stehendes Totholz belassener Windwurf Anteil der Moderverjüngung sogar auf 72 Ofo angestiegen (Abbildung 41). Der Unterschied zwischen den beiden Aufnahmejahren ist durch Zubzw. Abgänge in der Fichtenverjüngung zu erklären: nahezu alle zwischen 1992 und 1997 abgestorbenen bzw. verschwundenen kleinen Fichten wuchsen unabhängig von Moderholz, während sich der überwiegende Teil der neu erfassten Individuen auf diesem Substrat etabliert hatte. Bei der Vogelbeere ist dagegen eine Substratpräferenz nicht zu erkennen, lediglich 3 Ofo wachsen aufvermoderndem Holz. Im stehenden Totholz ist der relative Anteil der Moderverjüngung deutlich geringer. Dies liegt aber vor allem an der reichlichen Ansamung von Fichten nach der Vollmast 1989 um die Stammfüße abgestorbener Bäume herum, wo durch herabfallende Nadeln, Feinreisig und Rinde günstige Keimungsbedingungen entstanden sind. Abb.42 Aufvermoderndem Holz umgestürzter Bäume oder alter Baumstümpfe etabliert sich bereits unter dem Schirm des Altbestandes reichliche Fichten-Naturverjüngung ("Rannenverjüngung"). 0 gesch lossenes stehendes Altholz Totholz Moderholz Wurzelteller belassener Windwurf Sonstige In der belassenen Windwurffläche wachsen etwas mehr als 10 % der Fichtenverjüngung auf Wurzeltellern, bei der Vogelbeere sind es ca. 15 % der Individuen (Diese erhöhten Plätze sind sicherlich auch beliebte Sitzwarten fürvögel...). Abb.47 Verteilung der Baumverjüngung auf verschiedene Kleinstandorte im Jahr ,,,w iiwi h 5; se MM f
: Während unter dem Schirm des Altholzes die Hälfte der Fichten kleiner als 20 cm (Höhenklasse (HK) 0) und etwas mehr")

 mehr erfolgt ist und diese deshalb in der Grafik fehlen!")



79 Abb. 43 Höhenentwicklung der Naturverjüngung Höhenentwicklung Bereits bei der Erstaufnahme 1992 wurden deutliche Unterschiede in der Höhenentwicklung bei der Fichte, v. a. aber bei der Vogelbeere augenscheinlich (Abbildung 43): Während unter dem Schirm des Altholzes die Hälfte der Fichten kleiner als 20 cm (Höhenklasse (HK) 0) und etwas mehr als die Hälfte der Vogelbeeren kleiner als 50 cm (HK 1) waren, nahm der Anteil der höheren Pflanzen im lichten Schirm des Totholzes und auf der Windwurffläche markant zu. In den fünf Jahren bis zur Wiederholungsaufnahme 1997 hat sich im geschlossenen Altholz an der Höhenverteilung wenig geändert: das Wachstum der Bäume unter Schirm ist sehr langsam. Bemerkt werden muss hier, dass bei dieser Aufnahme keine Erfassung der Pflanzen unter 20 cm (Jungwuchszählung) mehr erfolgt ist und diese deshalb in der Grafik fehlen! In den Windwurf- und Totholzflächen ist dies ganz anders: Hier haben die Bäume einen bemerkenswerten Höhenzuwachs gemacht. Etwa die Hälfte der Fichten ist mittlerweile über 1 m, der überwiegende Teil der Vogelbeeren in der Sturmwurffläche sogar über 3 m hoch! Deren durchschnittlicher Höhenzuwachs war z. B. im Totholzsaum 24 cm pro Jahr und auf der Windwurffläche 26 cm pro Jahr, während die Vogelbeeren im Altholz nur 8 cm pro Jahr in diesem Zeitraum an Höhe zulegen konnten. Zur Charakterisierung des Verteilungsmusters wurde bei der Verjüngung der Aggregationsindex nach CLARK und EVANS (1954) berechnet (Kapitel 3.3). Dieser schwankt für die Fichte zwischen 0,31 und 0,60 und für die Vogelbeere zwischen 0,43 und 0,78 in den verschiedenen Bestandssituationen. Aggregationswerte unter eins weisen auf eine Tendenz zur Klumpung hin. Für beide Baumarten ist also eine "Rottenbildung" rechnerisch eindeutig nachzuweisen. 60 geschlossenes Altholz 60 stehendes Totholz 60 belassener Windwurf o. ~L...!--II! o o Höhenklasse Höhenklasse Höhenklasse 80






. Abb.")
80 Während bei der Fichte sicherlich die Verteilung verjüngungsfreundlicher Kleinstandorte maßgeblich ist für diese Rottenstruktur, dürfte bei der Vogelbeere v. a. die Art der Samenausbreitung der entscheidende Grund hierfür sein, denn die Samen werden nicht einzeln mit dem Kot von Tieren ausgeschieden. Neben dem Verteilungsmuster innerhalb der einzelnen Baumarten interessiert auch die Verteilung zwischen den Arten, also die Frage, ob eine intensive Durchmischung oder eine klare räumliche Trennung zwischen den Baumarten zu beobachten ist. Für die Naturverjüngung auf dem Transekt wurde ein Segregationsindex (PIELOU 1977; Kapitel 3.3) zwischen 0,21 (Totholzbereiche) und 0,75 (geschlossenes Altholz) berechnet. Bei Fichte und Vogelbeere ist demnach ein eher räumlich getrenntes Vorkommen zu beobachten. Geschlossenes Altholz Probequadrat Nr. 4 Probequadrat Nr x~>i'x _x ~ x x Xx 0 : 0 ':cx+'f 0 ox~.;. t t1j< xx t'> x Xx x x e e x 0 0 'I< " xxt> Stehendes Totholz Probequadrat Nr. 11 Probequadrat Nr. 12 Anhand der Stammfußkarten ausgewählter Probequadrate in den verschiedenen Stadien soll das Verteilungsmuster der Baumverjüngung noch einmal verdeutlicht werden (Abbildung 44). Abb.44 Ausgewählte Stammfußkarten aus den verschiedenen Bestandssituationen (M 1 : 200). Probequadrat Nr. 14 Belassener Windwurf Legende - Fichte - Vogelbeere - Pionierbaumarten: Birke, Aspe, Weiden Baum, BHD > 10 cm + Baum, BHD <" 10 cm, X Höhe> 1,0 m Baum;-Höhe 0,2-0,99 m stehendes Totholz o Baumstumpf y entwurzelter Baum I 10 m 81


.")
81 4.4.4 Einfluss von Reh und Rothirsch Pflanzenfresser können die Vegetationsentwicklung maßgeblich beeinflussen. Der Einfluss von Reh und Rothirsch auf die Naturverjüngung lässt sich einfach dokumentieren. Festgehalten wurden sowohl der aktuelle Verbiss (Leittrieb, Nebentrieb, Leit- und Nebentrieb) in den Aufnahmejahren als auch Beschädigungen der Pflanzen durch Schälen (Abbildung 45) oder Fegen. Abb.45 Von Rothirschen geschälte Vogelbeere neben unbeeinflusster Fichten-Naturverjüngung. Bei der Fichte konnten in beiden Aufnahmejahren kaum Pflanzen beobachtet werden, die in irgendeiner Weise beeinträchtigt waren. Bei den Vogelbeeren war dagegen 1992 ein bedeutender Einfluss von Reh und Rothirsch erkennbar, v. a. im stehenden Totholz. Dort waren lediglich 30 % der Pflanzen völlig unbeschädigt (Abbildung 46). Die Totholzsäume um Windwurfflächen herum scheinen sehr beliebte Aufenthaltsorte zu sein, da sich dort die krautige Vegetation und die Gehölzverjüngung üppig entwickeln und diese Zonen in den ersten Jahren nach dem Absterben der Altbäume gut begehbar sind war ein deutlicher Rückgang beim Verbiss an dervogelbeere zu verzeichnen. Einerseits üben die kreuz und quer herumliegenden Stamm- und Gipfelstücke der zusammenbrechenden toten Bäume einen gewissen "Zauneffekt" aus, andererseits sind viele junge Bäume mittlerweile so hoch gewachsen, dass die jungen Triebe von Reh und Rothirsch nicht mehr erreichbar sind ,12 e., B c: 40 < ~ 60 B c: < Abb.46 Einfluss von Reh und Rothirsrh auf die Naturverjüngung. 0 geschlossenes stehendes belassener Altholz Totholz Windwurf 0 geschlossenes stehendes belassener Altholz Totholz Windwurf Schäl-I Fegeschaden Leit- und Nebentriebverbiss Nebentriebverbiss Leittriebverbiss ohne Schäden 82

 der Sturmwürfe ereigneten sich in den Bergfichtenwäldern.")
82 5.1 Störungen als Motor der Waldentwicklung Die Stürme im Sommer 1983 und Herbst 1984 haben zu den ersten tiefgreifenden Veränderungen in den Wäldern des Nationalparks Bayerischer Wald seit dessen Gründung am 7. Oktober 1970 geführt: Auf 173 ha wurden unzählige Bäume entwurzelt oder - in geringerem Maße - gebrochen und zu Boden gerissen. Meist handelte es sich um flachwurzelnde Fichten auf mineralischen und organischen Nassböden. Betroffen waren vor allem die Wälder in den Tal- und Hanglagen des Grenzkammes, lediglich 14,3 ha (8 0/0) der Sturmwürfe ereigneten sich in den Bergfichtenwäldern. Nicht berücksichtigt sind hier jedoch die zahlreichen über die gesamten Hochlagen verteilten Einzelwürfe, da sie meist in den Luftbildern nicht erkennbar waren. In der Mehrzahl - insgesamt verteilen sich die Windwürfe auf 43 Flächen - waren es in den Hochlagen kleine "Nesterwürfe", die durchschnittliche Flächengröße betrug nur 0,3 ha. Die beiden größten Sturmwurfflächen in den Waldabteilungen Scheerbach (2,1 ha) und Rachelkapelle (1,5 ha) erstrecken sich zu einem bedeutenden Teil in die Hanglagen hinein. Auch in den Hochlagen waren häufig Bestände auf Nassböden betroffen oder bildeten die Ausgangspunkte für die Windwürfe. Diese Beobachtung trifft ebenso für das Untersuchungsgebiet zu (Westteil des Windwurfs, Karte 2, Seite 54). Die Fichten in den Bergwäldern sind hervorragend an die rauen Lebensbedingungen angepasst (On et al. 1997): Mit ihren langen, schlanken Kronen und den abholzigen Schäften (niedriger Schlankheitsgrad oder h/d-verhältnis) trotzen sie auch hohen Belastungen durch Wind und Schnee. Auf der Untersuchungsfläche wurde ein durchschnittlicher Kronenanteil von ca. 60 0J0 der Baumhöhe ermittelt bei sehr geringen Kronendurchmessern (3,6 m bei GrundflächenmitteIund 4,8 m bei Oberhöhenstamm), das Verhältnis Kronendurchmesser zu -länge ("Plumpheitsgrad") betrug 1 :4. Vor allem die stärksten und höchsten Bäume hatten darüber hinaus ein sehr günstiges h/d-verhältnis von 54 (Oberhöhenstamm). Erhöht wird diese individuelle Widerstandskraft durch das Verteilungsmuster der Bäume: häufig zusammengedrängt zu dichten Gruppen ("Rotten") scheinen sie sich gegenseitig zu stützen. Außerdem bilden sie mit ihren teilweise extrem asymmetrischen Kronen - zum Zentrum der Gruppe hin sehr kurz und wenig ausladend, nach außen jedoch sehr lang - einen geschlossenen Kronenmantel aus steil herabhängenden Ästen, auf denen der Schnee gut abgleitet. Trotzdem kommt es auch in natürlichen Fichtenwäldern der Bergstufe und der borealen Nadelwaldzone immer wieder zu Windwürfen verheerenden Ausmaßes: Von großflächigen Sturmwür- Abb.47 Windwürfe geben den Anstoß zur Erneuerung von Waldbeständen. 83
.")

83 fen in Mittelschweden 1931/32 berichtet SER NANDER (1936). In der Folge von "VlVIAN" und "WIEBKE" im Februar 1990 fielen allein in der Schweiz rund 4,5 Mio. Festmeter Schad holz an, der überwiegende Teil in den Gebirgswäldern. Die bedeutendste Windwurffläche mit 150 ha befindet sich in Disentis im Kanton Graubünden (LAESSIG und SCHOENENBERGER 1993). Die orkanartigen Stürme in der zweiten Hälfte des 19. Jahrhunderts verwüsteten weite Teile des Bayerischen Waldes: "... durch die destruktive Wirkung der allgegenwärtigen Natur... hat der Windsturm vom Jahr 1868 hundert Tausende von Fichtenstämmen niedergeworfen... Am 28. Oktober 1870 kam ein noch gewaltigerer Windsturm, der den ersteren an Zerstörungskraft weit übertroffen hat. Es soll eine wahre Höllennacht gewesen sein... " (zeitgenössische Schilderung, zitiert nach SCHERZlNGER 2000). Der großflächige Borkenkäferbefall nach den Stürmen 1983/84 im Nationalpark Bayerischer Wald und die erneute Massenvermehrung seit Mitte der 90er Jahre führen dies sehr eindrücklich vor Augen (HEURICH et al. 2001). Auch nach dem "Großen Sturm" 1868/70 wird von einer Massenvermehrung des Buchdruckers berichtet. Das Verhältnis von aufgearbeitetem Sturmholz zu Käferholz wurde von ZIERL (1972) auf 1 : 1,5 kalkuliert (zit. nach SCHERZlNGER 1996). Verschiedene Autoren (FALINSKY 1976, MAYER 1983, LEIBUNDGUT 1993, KORPEL 1995) kommen nach eingehenden Beobachtungen in Fichten (Nat)urwäldern zu dem Schluss, dass die Erneuerung dieser Nadelwälder häufig durch großflächigen Zerfall ausgelöst wird, hervorgerufen durch abiotische (Wind, Schnee, Feuer) und/oder biotische Faktoren (Insekten, Pilze). Abb.48 Nach den Stürmen 1983/84 wurden zunächst vor allem die Bestände in der unmittelbaren Nachbarschaft der Windwurfflächen von Borkenkäfern befallen und abgetötet. Plattenhauserriegel, Abt. Bärenloch. Durch das reichliche Angebot an bruttauglichem Material für Borkenkäfer nach Sturmwürfen in Form umgestürzter oder noch stehender, aber in irgendeiner Weise geschädigten Fichten (Wurzelschäden durch die mechanische Belastung, Rindenbrand durch plötzliche Freistellung usw.). entwickeln sich in deren Folge häufig Borkenkäfergradationen, sofern der Witterungsverlauf in den nachfolgenden Jahren für die Populationsentwicklung der Insekten günstig ist. Als maßgeblichen Grund für die hohe Schadenanfälligkeit der Fichtenwälder wird deren Wuchsverhalten angesehen, d. h. die starke Neigung zur Gleichförmigkeit mit zunehmendem Bestandesalter trotz großer Altersunterschiede innerhalb der Bestandesglieder. Im urwaldartigen Bödmerenwald im Kanton Schwyz wurden bei der Altersbestimmung von über 100 Bäumen Unterschiede von bis zu 150 Jahren innerhalb einer Durchmesserklasse festgestellt, vermutlich wegen mehr oder weniger langer Überschirmung in der Jugendentwicklung (LEIBUNDGUT 1993). Auch in der Höhenentwicklung konnte eine Angleichung unterschiedlich alter Individuen nachgewiesen werden (On et al. 1997). Im Untersuchungsgebiet kann diese Beobachtung teilweise bestätigt werden: Im nördlich an den Windwurf angrenzenden Altholzteil (PO 1-9) ist die Durchmesser- und Höhendifferenzierung trotz deutlicher Unterschiede im Alter der Bäume (zwischen 80 und 200 Jahrringe) ebenfalls sehr 84
 betrug 90 % des")
 Ansamung und Entwicklung junger Waldbäume in den reifen Bergfichtenwäldern nicht unterschätzt werden.")
84 gering. Im südlich angrenzenden Altbestand dagegen sind sowohl Durchmesser- als auch Höhendifferenzierung wesentlich stärker ausgeprägt: Der errechnete Artprofilindex nach PRETZSCH (1996) betrug 90 % des maximal möglichen! Hier wurden zwischen 90 und 252 Jahrringe bei 21 Bäumen gezählt. Neben der Erneuerung, ausgelöst durch großflächige Störungen, darf jedoch die (episodische) Ansamung und Entwicklung junger Waldbäume in den reifen Bergfichtenwäldern nicht unterschätzt werden. So wurden, trotz hoher Holzvorräte und der damit verbundenen Überschirmung durch die alten Bäume, in den bis 1992 noch unversehrt gebliebenen Altholzbeständen bemerkenswerte Verjüngungsdichten nachgewiesen: lediglich auf zwei von 25 Probequadraten wurde keine Baumverjüngung gefunden; die höchste Anzahl betrug 133 Fichten in einer Aufnahmefläche (100 m 2 ), entsprechend Individuen je Hektar! Der Anteil der Vogelbeere an der Gesamtverjüngung lag 1992 bei 27 Ofo. Letztendlich weisen die erheblichen Altersunterschiede innerhalb der untersuchten Waldbestände, selbst von Bäumen, die in unmittelbarer Nachbarschaft stehen, auf die unregelmäßige Ansamung und Entwicklung einer neuen Baumgeneration auf kleiner Fläche hin. Die Fichtenverjüngung ist aber nicht gleichmäßig verteilt; bereits die Keimlinge und Sämlinge finden sich zu Rotten zusammen. Ursache hierfür ist die überaus dichte Konkurrenzvegetation, die im Kampf um Nährstoffe und Wasser dem Jungwuchs meist überlegen ist. Dadurch werden die kleinen Fichten auf "privilegierte Inseln" (On et al. 1997) verdrängt, die frei oder nur schütter von Berg-Reitgras, Drahtschmiele, Wald-Hainsimse oder Alpen-Frauenfarn bewachsen sind. Die Vogelbeere scheint ein größeres Durchsetzungsvermögen gegenüber dieser Konkurrenzvegetation zu besitzen; möglicherweise ist es auch die "Startdüngung" in Form von Kot, mit dem die Samen ausgeschieden (und stratifiziert) werden. Sie wächst zwar ebenfalls meist in kleinen Gruppen, eine Abhängigkeit vom Vegetationsmosaik oder von besonderen Kleinstrukturen ist allerdings nicht erkennbar. Vielmehr scheint das Verteilungsmuster durch die Art der Samenverbreitung bedingt zu sein. Für die Fichtenverjüngung bildet vermoderndes Holz in Form von alten Baumstümpfen oder umgestürzten Bäumen das bedeutendste Keimbett: 1992 wuchsen mindestens 52 Ofo, 1997 sogar 72 Ofo der kleinen Fichten in der Untersuchungsfläche auf diesem Substrat. Im Rahmen der Verjüngungsinventur in den Hochlagen des Nationalparks Bayerischer Wald wurden 45 Ofo der erfassten Verjüngungspflanzen auf Moderholz bzw. in dessen unmittelbarer Nachbarschaft gefunden (HEURICH 2001 b). Dabei beträgt der Anteil der Baumstümpfe selbst nur 0,3 Ofo, bei Berücksichtigung ihrer unmittelbaren Umgebung 2-3 Ofo der Gesamtfläche. Auch STOECKLI (1995) beobachtete positive Effekte in der Nachbarschaft von Totholz: erhöhter Nährstoffeintrag durch Auswaschung aus dem Moderholz, geringere Gefahr der Austrock.nung, Durchtränkung des Bodens mit Rindeninhalts- und Holzabbaustoffen mit fungizider (pilzhemmender) Wirkung und - sicherlich von großer Bedeutung - den Zugang zum Mykorrhizapotenzial des Bodens. Abb.49 Bereits in geschlossenen Bergfich ten wä Idern etabliert sich Naturverjüngung, sofern geeignete Kleinstandorte vorhanden sind. 85


85 Flächenmäßig bedeutender als Baumstümpfe ist das liegende Totholz umgestürzter oder zusammengebrochener Bäume. Auf einzelnen Probequadraten in den lebenden Altholzbeständen wurde eine Totholzbedeckung von bis zu 12 % ermittelt. Auch REIF und PRZYBILLA (1998) beobachteten im Gebiet des Lusen, dass der Hauptteil der Verjüngung auf Moderholz wächst, obwohl es den geringsten Anteil an der Bestandesfläche einnimmt. Außerdem wird durch die liegenden Stämme das bestehende Verteilungsmuster der Bäume durchbrochen, indem verjüngungsfreundliche Strukturen in Bereichen entstehen, die vorher von krautiger Konkurrenzvegetation beherrscht wurden. FALINSKY (1976) konnte im Urwald von Bialowieza feststellen, dass Stellen von vielen krautigen Pflanzen nicht besiedelt wurden, auf denen Holz lag und zerfiel, auch zehn Jahre nach dessen Verschwinden. Darüber hinaus beobachtete er, dass aufzwei Drittel seiner Untersuchungsflächen im Laufe von zehn Jahren eine intensive Anhäufung, aber auch Zerfall von liegendem Totholz stattfand. Als die wichtigsten, die Ansamung und Entwicklung von Baumjungwuchs fördernden Effekte neben der Eindämmung von Konkurrenten nennt EICHRODT (1969) die physikalischen Eigenschaften des Moderholzes wie ausgeglichener Wasserhaushalt, bessere Durchwurzelbarkeit, geringere Verdichtungstendenz und die im Vergleich zur Umgebung höheren Tempe- ''Zu großer Vorsicht ermahnt der pfadlose Boden, der aus einem Gewirr von zerbrochenen, dahin gestreckten, halb oder ganz vermoderten, mit Moos, Farm und anderen Waldpflanzen bedeckten Stämmen und wunderlich untereinander verwachsenen Wurzeln besteht, aus denen sich die Kolosse des Waldes erheben. Mit kaum glaublicher Schnelligkeit entwickelt sich überall die junge Fichten waldung, die alle Lücken einnimmt und die zahlreichen mit Moos bedeckten Lagerhölzer mit Legionen vonjungen Stämmchen überzieht" (GOEPPERT 1868). raturen (zit. nach MAI 1997). Außerdem apern die erhöhten Bereiche im Frühjahr rascher aus, die Vegetationszeit wird dadurch verlängert. Mangel an Licht und Wärme in geschlossenen Waldbeständen der klimatisch benachteiligten Hochlagen wird oft als Grund für das Ausbleiben von Baumverjüngung genannt. Dies kann zumindest für das Untersuchungsgebiet nicht bestätigt werden. Vielmehr zeigt sich, dass in Bestandeslücken Berg-Reitgras und Drahtschmiele die effektiveren Nutzer des höheren Lichtangebots sind (siehe Stammfuß- und Kronenkarten!). Außerdem sammeln sich dort während eines Winters die größten Schneemengen an und bleiben auch am längsten liegen! Letztlich deuten auch die überwiegend fehlgeschlagenen Versuche der Vergangenheit, die natürliche Verjüngung der Hochlagenwälder durch Plenterhiebe, Femelschläge und Bodenbearbeitung einzuleiten (ZIERL 1972). in diese Richtung. Die Beobachtungen hier wie in anderen Bergfichtenwäldern zeigen, dass sowohl Fichte als auch Vogelbeere sehr lange Zeit im Schatten alter Bäume ausharren können. Für die Fichte ermittelte KORPEL (1995) in Urwäldern der Westkarpaten 100 bis 160 Jahre, die in Unterdrückung überdauert werden können; LEIBUNDGUT (1993) berichtet von Fichten im Urwald Scatle in der Schweiz, die sich nach 280 Jahren Überschirmung noch normal entwickelten. Unbestritten ist, dass für ein zügiges Wachstum sowohl bei Fichte als auch bei Vogelbeere ein hohes Lichtangebot benötigt wird. Durch ein gedämpftes Jugendwachstum wird aber die Hauptwachstumsphase in ein höheres Alter verschoben, wodurch sich die Lebenserwartung der Bäume erhöht (LEIBUNDGUT 1993). 86
.")

86 5.2 Veränderungen nach dem Sturm Stürme reißen unterschiedlich große Lücken in den Wald und verändern dadurch die Umweltbedingungen ganz entscheidend. Neben der Öffnung des Kronendaches ist es vor allem die Schaffung eines außerordentlich vielfältigen Mosaiks unterschiedlichster Kleinstandorte (JEHL 1995). In den ersten Jahren nach dem Sturm versperren ineinander verkeilte Baumstämme und Kronen, oft mehrere Meter über dem Waldboden hängend, aber auch die herausgehebelten Wurzelteller den Blick über die Windwurffläche. In dieser Zeit sind die Wurzelteller und umgestürzten Bäume vor allem Schattenspender, tragen zur Windruhe in bodennahen Schichten bei und schaffen so ein besonderes Kleinklima. Durch die herausgerissenen Wurzeln wird der Grasfilz zerstört und der Mineralboden auf beträchtlicher Fläche freigelegt. Im Durchschnitt wurde eine Wurzeltellergröße - und somit auch die Größe des freigelegten Bodens - von 5 m 2 ermittelt. Im Untersuchungsgebiet mi~ etwas mehr als 400 Bäumen je Hektar beträgt die. Gesamtfläche des offenen Bodens hochgerechnet etwa 2000 m 2 /ha oder 20 0/0. Dort bietet sich die Chance für Neueinwanderer ("Pioniere"). die unterschiedliche Strategien zur Besiedlung solcher spontan entstandenen Lebensräume entwickelt haben: Birken, Aspen und Weiden sind bereits im jungen Alter geschlechtsreif und bilden dann regelmäßig und reichlich Samen aus, die mit dem Wind sehr weit verfrachtet werden. Die Früchte von Himbeere und Holunder werden von Tieren effektiv verbreitet, da sie eine begehrte Nahrung sin.d. Im Boden eingeschlossen bleiben deren Sa '~en sehr lan'ge keimfähig,(fischer 1987). Wird der Boden durch die herausgehebelten Wurzeln umr,, gestürzter Bäume freigeleg~, entfalten sie sich aus dem Samenreservoi~ des Bodens ("Diasporenbank"). In dieser Höhenstufe scheinen aber nur wenige Samenpflanzen diese neuen Lebensräume besiedeln zu können. Neben Schmalblättrigem Weidenröschen und Himbeere wurden nur noch die Fadenbinse (Juncus filiformis) und einige kleine Horste von Igel- (Carex echinata) und Hasenpfotensegge (C. ovalis) entdeckt, Pioniergehölze wurden ebenfalls nur wenige beobachtet. In den belassenen Windwurfflächen in der Bergmischwaldstufe wurden die offenen Bodenflächen sehr dicht von sog. Schlagflurarten besiedelt, jedoch waren insgesamt betrachtet die Veränderungen in der Artenzusammensetzung der Krautschicht nach dem Sturm ebenfalls gering (JEHL 1995). Bei den Moosen wurde ein Rückgang sowohl in der Deckung als auch in der Artenzahl auf den Sturmwurfflächen beobachtet. PALISAAR (1997) stellte dagegen bei eingehenden Untersuchungen der Moosvegetation in Au-Fichtenwäldern und Bergmischwälder,n des Nationalparks Bayerischer Wald fest, dass dort Windwürfe eine deutlich höhere Artenzahl beherbergten als die angrenzenden Altholzbestände und dass die belassenen Windwürfe wiederum höhere Artenzahlen aufwiesen als die geräumten. Als entscheidenden Faktor nennt er die Entstehung von Sonderstandorten: Auf freigelegtem Mineralboden und an der Wurzeltellerunterseite entwickelten sich Arten aus der Diasporenbank des Bodens. Diese wurden in der Windwurffläche Gfeichtethöh sicherlich übersehen, da sie in der Anfangsentwicklung sehr Abb.50 Durch Windwürfe entsteht ein vielfältiges Mosaik unterschiedlicher Kleinstandorte. 87
 klein und unscheinbar sind und deshalb ausgezeichnete Artenkenntnisse und sehr genaue Erhebungsmethoden voraussetzen.")



87 Abb. 51 Auf der überwiegenden Fläche bleibt die typische Waldbodenvegetation erhalten. (Im Vordergrund Vermarkung eines Probequadrats) klein und unscheinbar sind und deshalb ausgezeichnete Artenkenntnisse und sehr genaue Erhebungsmethoden voraussetzen. Bei den Deckungsanteilen der Moose beobachtete er jedoch ebenfalls einen Rückgang, verursacht durch die Ausbreitung der wesentlich konkurrenzkräftigeren Samenpflanzen. Schreitet die Vermoderung des lagernden Holzes fort, sackt es allmählich zu Boden und bildet im Lauf der Zeit ein wichtiges Keimbett für die neue Waldgeneration. Von HOFGAARD (1993) wird für Südschweden eine Zeitspanne von 50 Jahren, von HILGARTER (1971) im Urwald Scatle (Schweiz) eine Spanne von 40 Jahren genannt, die erforderlich ist, bis das Totholz ausreichend vermodert ist, um von Baumverjüngung besiedelt werden zu können. Bei der ersten Wiederholungsaufnahme in der Windwurffläche Gfeichtethöh wurden bereits 13 Jahre nach dem Sturm die ersten kleinen Fichten auf vermodernden Baumstämmen beobachtet! Besonders rasch scheinen sich im Übergangsbereich zwischen Wurzelteller ~ und Baumstamm günstige Keimungsbedingungen zu entwickeln: zwischen den Wurzelanläufen sammeln sich Wasser und herabfallende humose Erde, die Holzzersetzung scheint dort auch besonders rasch fortzusch reiten. fallenen Nadeln und die absterbenden Feinwurzein liefern leicht hydrolysierbare, stickstoffreiche Biomasse, die sehr schnell von Bodenmikroorganismen mineralisiert wird (BEUDERT 1999). Dabei werden auch Anteile des älteren, chemisch recht stabilen Humus abgebaut. Darüber hinaus fördert der erhöhte Lichteinfall und das damit verbundene höhere Wärmeangebot die Aktivität der Bodenorganismen. Dies fördert das Wachstum der vorhandenen Krautschicht, insbesondere des Berg-Reitgrases enorm. Am Stammfuß der abgestorbenen Bäume jedoch häufen sich neben Fichtennadeln herabfallendes Feinreisig und insbesondere abblätternde oder von Spechten abgehackte Rindenstücke an. Dieses organische Material dürfte eine vor allem verdämmende Wirkung auf Gräser ausüben (Mulcheffekt). Außerdem können sich Gräser und Kräuter auf den Ca-reichen und N-armen Rindenbruchstücken (BEUDERT, mündliche Mitteilung) offensichtlich nicht etablieren. Abb.52 Rascher als erwartet samen sich kleine Fichten auf dem liegenden Totholz an. Unter den von Borkenkäfern abgetöteten Bäumen sind die Veränderungen etwas anders: die abge 88

 auf einer Windwurffläche in den Hanglagen des Nationalparks Bayerischer Wald (Abteilung Schachtenriegel): Dort waren 80 % der drei Jahre nach")
88 5.3 Entwicklung einer neuen Waldgeneration Da der Sturm die meisten Bäume nicht abgebro~ chen, sondern entwurzelt hatte und die Stämme und Kronen oft ohne Bodenkontakt übereinander liegen, wurde der Großteil des bereits vor dem Sturm vorhandenen Baumjungwuchses nicht zerstört (Bei der Fällung von Bäumen ist dies ganz anders: Hier schlagen Stamm und Krone flach auf dem Boden auf und erdrücken alles, was sich darunter befindet!). Das Verteilungsmuster der kleinen Bäume auf der Windwurffläche macht dies deutlich. Ähnliche Beobachtungen machten HOMANN und ENGELS (1991) auf einer Windwurffläche in den Hanglagen des Nationalparks Bayerischer Wald (Abteilung Schachtenriegel): Dort waren 80 % der drei Jahre nach dem Windwurf vorgefundenen Naturverjüngung schon vor dem Sturm gekeimt, von den erfassten kleinen Fichten waren es sogar 94 Ofo! Letztendlich weisen auch die ermittelten Alter und der Zuwachsverlauf von 47 Bäumen im Untersuchungsgebiet darauf hin, dass sie sich bereits im Schatten des alten Bergfichtenwaldes etabliert hatten und teilweise Jahrzehnte ausharrten, bevor dieser von den gewaltigen Stürmen 1868 und 1870 zu Boden geworfen wurde. Die veränderten Umweltbedingungen, die aber keinesfalls auf der ganzen Fläche gleich sind, sondern durch das vielfältige Mosaik unterschiedlicher Kleinstrukturen wie aufgeklappte Wurzelteller und liegendes Totholz sehr kleinräumig variieren, bieten günstige Bedingungen für die Entwicklung der neuen Baumgeneration. Die Vogelbeeren hatten sehr rasch auf die Veränderungen reagiert; bereits 1992 war ein Drittel der Individuen über 3 m hoch. Die Fichten zeigten sich anfangs wesentlich zaghafter im Wuchs, jedoch waren bei der Wiederholungsaufnahme bereits 40 Ofo über einem Meter hoch. Abb. 53 Der Großteil der bereits vor dem Sturm vorhandenen Naturverjüngung überlebte das Ereignis. Die intensive Blüte und Samenbildung der noch lebenden alten Fichten in den Hochlagen in den Jahren 1989, 1992 und 1995 wurde auch auf der Windwurffläche und in den abgestorbenen Altbeständen sichtbar. Auf Wurzeltellern, freigelegten Mineralbodenflächen und um die Stammfüße der abgestorbenen alten Bäume hatte sich ein Heer von Sämlingen eingestellt. Einige besiedelten sogar bereits die im Herbst 1984 geworfenen Bäume! Pionierbaumarten wie Aspe, Birke oder Weide haben auch 13 Jahre nach dem Sturm zahlenmäßig keine Bedeutung, weder in der Windwurffläche, noch in den abgestorbenen Altbeständen. Das Verteilungsmuster des bisher beobachteten Baumjungwuchses ähnelt sehr dem des Altbestandes. Der Großteil der jungen Fichten befindet sich auf oder in unmittelbarer Nachbarschaft von alten Baumstümpfen, um die Stammfüße der abgestorbenen Altbäume oder auf dem durch die herausgerissenen Wurze ln freigelegten Boden


89 Abb. 54 Die neue Waldgenera tion entwickelt sich im Standortsmosaik sehr vielfältig und reich strukturiert (Au fnahme vom Mai 1993). Durch die - zeitlich sicherlich um einige Jahrzehnte verzögerte - Besiedelung des bereits liegenden oder in den folgenden Jahren zusammenbrechenden Totholzes wird sich aber vielleicht - über einen längeren Zeitraum betrachtet - eine veränderte Bestandesstruktur entwickeln. Die horizontale Struktur des zukünftigen Waldes, d. h. das derzeitige Verteilungsmuster der Bäume, wird ergänzt durch lineare, möglicherweise stabilitätsfördernde Bestandesstrukturen. Nachdem die meisten Bäume entsprechend der Windrichtung des Sturmes gefallen sind, werden die "Rannen" auf den liegenden Stämmen zukünftig schmale und langgezogene Gruppen bilden, ausgerichtet nach dem Muster des geworfenen Holzes. Dadurch werden auch Flächen von Waldbäumen besiedelt, die über sehr lange Zeit von Gras, Farnen oder Zwergsträuchern domin iert waren. Auch die Vertikalstruktur wird sich vermutlich ändern, denn neben der bereits vorhandenen und durch die veränderten Umweltbedingungen zügig aufwachsenden neuen Baumgeneration wird sich in deren Schatten die zukünftige Rannenverjüngung sehr langsam entwickeln. Dies führt aber auch wieder zu einer großen AItersdifferenzierung der zukünftigen WaIdbestände. Die Befürchtung, dass aus den Windwurfund Borken käferflächen g leicha Itrige und gleichförmige Wälder entstehen werden, ist nach den bisherigen Beobachtungen unbegründet, zu kleinräumig wechseln die Rahmenbedingungen für das Wachstum der neuen Waldgeneration. Interessant ist in diesem Zusammenhang auch der Einfluss von Reh und Rothirsch: In den ersten Jahren schützte der "Windwurfverhau" relativ gut die jungen Waldbäume. Nur wenige Pflanzen waren dort verbissen oder geschält. Dagegen bildeten der angrenzende Totholzsaum mit der sich üppig entwickelnden Vegetation und die unversehrt gebliebenen Altbestände beliebte Aufenthaltsorte. Die wesentlich höhere Verbissbelastung macht dies deutlich. Im Laufe der Zeit entwickelte sich jedoch durch die kreuz und quer liegenden Stammteile der zusammenbrechenden toten Bäume ein dem Windwurf vergleichbarer "Zauneffekt". Während bei der Wiederholungsaufnahme 1997 der Anteil der verbissenen Pflanzen in den Altholzteilen annähernd gleich geblieben ist, hat dieser im Totholzsaum bereits deutlich abgenommen. In der Windwurffläche waren mittlerweile die meisten Vogelbeeren so groß geworden, dass Reh und Rothirsch die jungen Triebe nicht mehr erreichen konnten. Die Beobachtungen in der belassenen Windwurffläche im Bergfichtenwald decken sich sehr gut mit den bisherigen Ergebnissen von den Untersuchungsflächen in den Aufichtenwäldern der Tallagen (FISCHER et al. 1990, SCHMID 1994, BREIT 1999) und Bergmischwäldern der Hanglagen (HOMAN und ENGELS ;991, JEHL.1995, TRAUT,und WAGNER 1997), wenngleich die Geschwindigkeit der Entwicklung und die Vielfalt der beteiligten Arten im Bergfichtenwald wesentlich geringer ist. Auch KORPEL (1995) kommt zu dem Ergebnis, dass Windwürfe kleiner als 10 ha verhältnismäßig schnell von den sog. Schlusswaldbaumarten besiedelt werden. Seine Beobachtung, dass in der Anfangsphase der Entwicklung Pionierbaumarten innerhalb weniger Jahre einen geschlossenen Vorwald bilden, unter deren Schirm sich dann in der Folgezeit die Fichte etabliert, kann mit den nun 13-jährigen Beobachtungen im Nationalpark Bayerischer Wald nicht bestätigt werden. 90



90 6. Schlussberner kungen Welche Schlüsse lassen sich aus den bisherigen Beobachtungen im Nationalpark Bayerischer Wald ziehen? Anfang der 90er Jahre waren viele optimistisch: Die Borkenkäfer-Massenvermehrung nach den Sturmwürfen 1983 und 1984 schien innerhalb weniger Jahre ohne Zutun des Menschen zusa-mmengebrochen zu sein. Aber bereits vier Jahre später wurden die Hoffnungen zerstört. Die Beobachtungen vieler Fachleute, dass die Erneuerung Nadelholz-dominierter Waldbestände oftmals durch großflächigen Zerfall auf Grund abiotischer und biotischerstörfaktoren ausgelöst wird, scheinen sich zu bestätigen: Zwischen den Jahren 1983 und 2000 sind etwa ha der alten Bergwälder im Nationalpark Bayerischer Wald in der Folge von Windwürfen und Borkenkäferbefall abgestorben. Inwieweit die Dimension dieser Störungen jedoch durch die Vorgeschichte der Wälder - Glashüttennutzung, Waldweide, Forstwirtschaft, Räumung und Aufforstung nach den heftigen Stürmen in der zweiten Hälfte des 19. Jahrhunderts - oder die Einflüsse der jüngsten Vergangenheit - Umweltbelastung durch Luftschadstoffe, "Saurer Regen" - und die sich erhärtende Befürchtung einer deutlichen Klimaveränderung gesteuert wird, lässt sich bis heute nicht abschließend klären. Aber auch in dicht mit Baumjungwuchs bestandenen Bereichen ist ein sehr differenziertes Verteilungsmuster erkennbar. Es sind also über die gesamte Fläche betrachtet sehr deutliche Unterschiede in der Entwicklung der zukünftigen Wälder zu erwarten, rasch mit Bäumen bestockte Flächen wechseln mit solchen, in denen die Wiederbewaldung etwas mehr Zeit benötigt. Zehn Jahre nach dem Ausbruch des Mt. St. Helens, der ha Wald umgeworfen, umgebrochen oder verbrannt hatte, kommt DAYTON (1990) zu dem Schluss: "... die Wiederherstellung e'ines durch Naturereignisse massiv gestörten Ökosystems überlässt man am besten der Natur selbst... denn menschliche Eingriffe schadeten mehr als sie nutzten... beispielsweise das Aufarbeiten geworfener Bäume zugunsten vorhandener Naturverjüngung... wo die Hänge weder geräumt noch künstlich bepflanzt wurden, wachsen heute mehr Pflanzen in einergrößeren Vielfalt..." (zit. nach KENKetal. 1991) Offensichtlich wird jedoch, dass die Regenerationsfreudigkeit der Wälder, auch in den rauen Hochlagen des Mittelgebirges, ungebrochen ist. Dies zeigen sowohl die Untersuchungen auf Dauerbeobachtungsflächen als auch die Ergebnisse der Verjüngungsinventuren, die seit 1996 im zweijährigen Turnus durchgeführt werden. Dabei lässt sich beobachten, dass dicht mit jungen Bäumen bestandene Flächen wechseln mit solchen, in denen die Verjüngung derzeit noch spärlich ist. 91
91 Die Waldentwicklung nach Windwurf in den Hochlagen des Nationalparks Bayerischer Wald Im Frühjahr 1992 wurde analog den Tal- und Hanglagen in einem 0,83 ha großen naturbelassenen Windwurf, der 1984 in den Hochlagen entstanden war, eine Dauerbeobachtungsfläche eingemessen und vermarkt. Ausgehend von unversehrt gebliebenen Altbeständen quert der Transekt den durch Borkenkäferbefall entstandenen Totholzsaum und den Windwurf. Anhand detaillierter Aufnahmen, die zunächst im 5-jährigen Turnus durchgeführt werden, wird die Waldentwicklung nach Windwurf ohne direkte menschliche Einflussnahme beobachtet und dokumentiert. Neben den Vegetationsveränderungen kann die Bedeutung des Mosaiks unterschiedlichster Kleinstandorte für die Wiederbewaldung und die zukünftige Waldstruktur untersucht werden. Der untersuchte Waldbestand ist gemessen an der Höhenlage mit 816 VfmD/ha - 23 % davon liegendes und stehendes Totholz - sehr vorratsreich. Die Bäume sind in unterschiedlich dichten Rotten angeordnet. Trotz erheblicher Altersunterschiede wurde eine geringe Durchmesserdifferenzierung festgestellt. In den Altbeständen war bereits eine sehr üppige Krautschicht entwickelt (mittlere Deckung ca. 70 0/0), beteiligt waren aber nur wenige Arten. Kleinstrukturen spielten mit durchschnittlich 11 % Flächenanteil nur eine geringe Rolle wurde dort reichlich Naturverjüngung - im Mittel 2200 Fichten (73 0/0) und 800 Vogelbeeren (27 Ofo) je Hektar - gefunden. Fichten und Vogelbeeren wachsen überwiegend in Gruppen. Während bei Fichten begünstigte Kleinstandorte maßgeblich sind für die Klumpung wuchsen 53 Ofo, Ofo der Pflanzen auf Moderholz - scheint bei der Vogelbeere die Art der Samenverbreitung entscheidend zu sein. Auf den Totholzflächen hatte sich die Krautschicht nur wenig verändert: profitiert hatte v. a. das Berg Reitgras, dessen mittlere Deckung 1997 ca. 40 Ofo höher war als im Altholzteil. Auf den offenen Bodenflächen hatten sich Schlagflurarten (Himbeere, Weidenröschen) etabliert, 1997 betrug deren mittlere Deckung nahezu 20 Ofo. In der Moosschicht waren dagegen mittlere Artenzahl und Deckung deutlich geringer als in den Altbeständen. Im Totholzsaum wurden 8500 Fichten und 700 Vogelbeeren je Hektar, im Windwurf 6700 Fichten und 500 Vogelbeeren je Hektar erfaßt. Der Großteil der Baumverjüngung über 20 cm Höhe war sicherlich bereits vor dem Sturm vorhanden und hatte diesen überlebt. Pionierbaumarten spielen selbst im Windwurf nur eine unbedeutende Rolle, es wurden lediglich drei Individuen gefunden. Etwa 50 Ofo der Fichten im Windwurf wachsen auf Moderholz, 10 Ofo haben sich im Bereich der Wurzelteller etabliert. Im Totholzsaum ist der relative Anteil der Moderverjüngung deutlich geringer. Dies liegt vor allem an der reichlichen Ansamung von Fichten in der Folge der Mast 1989 um die Stammfüsse toter Bäume herum. Dort waren durch herabfallende Nadeln, Feinreisig und Rinde günstige Kei m u ngsbed i ngu ngen entsta nden. Bis 1997 hat die Zahl der Pflanzen ~ 20 cm um 7 % zugenommen bei annähernd gleichen Baumartenanteilen. Die Pflanzen in den Altbeständen hatten bis 1997 nur einen sehr geringen Höhenzuwachs, in den Totholzflächen war dagegen etwa die Hälfte der Fichten bereits über 1 m, der Großteil der Vogelbeeren sogar über 3 m hoch gewachsen. Deren mittlerer Höhenzuwachs betrug 25 cm/jahr. Fichten wurden kaum von Reh oder Rothirsch beeinträchtigt. Bei Vogelbeeren betrug 1992 der Anteil mit Verbiss-, Schäl- oder Fegeschäden 70 Ofo. Bis 1997 war der Anteil beschädigter Pflanzen auf ca. 30 % zurückgegangen. Die höchste Verbissbelastung wurde in den geschlossenen Althölzern und im Totholzsaum registriert. Die Beobachtungen in der belassenen Windwurffläche im Bergfichtenwald ähneln sehr denen auf Windwürfen im Aufichtenwald und Bergmischwald des Nationalparks, wenngleich dort die Geschwindigkeit der Entwicklung und die Vielfalt der Arten wesentlich höher sind. Im Gegensatz zu Beobachtungen in Naturwäldern anderer Regionen spielen Pionierbaumarten in den Hochlagen des Nationalparks Bayerischer Wald auch in der Anfangsphase der Wiederbewaldung nach großflächiger Störung nur eine untergeordnete Rolle. 92
92 Forest development after windthrow in high elevations of the Bavarian Forest National Park In 1992 a permanent research plot was established in a mountain spruce forest, comparable to the plots in the lowland spruce forests and mountain mixed forests. The plot is located in the northeastern part of the national park at an elevation of between 1140 and 1180 m. The transect starts in an undamaged tree stand, passes through a stand killed by bark beetle before passing into the windthrowarea. Detailed measurements have been conducted eve-: ry 5 years. The objective of the research is to document forest development in unmanaged areas. Besides the change of species diversity, one can also study the influence of the versatile mosaic of different forest sites such as Iying and standing dead trees or tree trun ks and root plates. The studied forest stand had high timber volume relative to the elevation: 814 m 3 /ha - about one quarter of the timber consisted of Iying and standing dead trees - were measured in the living stand. The trees generally stood in clusters. Tree cores (n=47) showed that the age difference is very high with trees of between 80 and 256 years old being identified. Diameter difference was minimal. Herbai vegetation covered 70% of the ground. The number of species present was low at an average of 12. Small structured sites were less prominent covering only 11 % of the stand area. The first survey results showed that natural regeneration was high, with 2200 spruce (73 %) and 800 mountain ash trees (27 %) per hectare being present. Both species were found to be growing in clusters. The decomposing wood is thought to be responsible for the grouping of the 'spruce as in 1992 and 1997, 53% and 72% respectively of the spruce regeneration grew there. The grouping of mountain ash is believed to be caused by seed spread. In the dead wood stands the herbai vegetation showed no significant change, in contrast to moss vegetation, which was significantly less. The Calamagrostis villosa benefits from the changing site conditions. On mineral soil areas, fireweed and raspberry had started to grow, covering an area of 20 % in In the area of standing dead trees around the windthrow 8500 spruce and 700 mountain ash trees per hectare were measured. In the windthrow area 6700 spruce and 500 mountain ash trees were counted. Pioneer trees playa minor role making up only 0.6 % of natural regeneration. Most of the natural regeneration over 20 cm in height was probably present before the storm. Approximately 50 % of the natural spruce regeneration are growing on decomposing wood. Approximately 10% of this growth are established around root plates. In the surrounding area of standing dead trees, regeneration on decomposing wood is clearly less. This is due to the natural seeding action of spruce around the base of dead trees. In 1989 the spruce had a seed year and the germination conditions were good, because of the falling needles, bark and small branches from the dead trees. By 1997 there had been an increase of 7% in trees over 20 cm. There was little change in the tree species. In the living stand, the growth of regeneration was smali, however in other areas the spruce had grown by over 1 m and the mountain ash by up to 3 metres in five years. The average tree growth was ca. 25 cm/year. In 1992 there were few signs of row deer and red deer browsing the spruce, whilst damage was evident on 70 % of all measured mountain ash individuals. By 1997 the damage to trees by wildlife was down to 30%. Most of the browsed trees are found in the living stands and the area of standing dead trees around the windthrow. The results found in the windthrow area within the mountain spruce forest are similar to those found in the windthrow areas ofthe mountain mixed and lowland spruce forests. However, the rate of forest development and species diversity is lower in mounta in spruce forests. In contrast to observations in natural forests of other regions, birch, aspen and willow do not playa major role in forest development even at the beginning of reforestation following large area disturbance. 93
93 Literaturverzeichnis ALBRECHT, L. (1990): Grundlagen, Ziele und Methodik der waldökologischen Forschung in Naturwaldreservaten. Schriftenr. Bayer. StMinELF, Naturwaldreservate in Bayern Bd.l; 219 S. BAYERISCHE LANDESANSTALT FUER WALD UND FORST WIRTSCHAFT (1995): Waldbauliche Dokumentation der flächigen Sturmschäden des Frühjahrs 1990 in Bayern und meteorologische Situation zur Schadenszeit. Freising; 336 S. BEUDERT, B. (1999): Veränderungen im Stoffhaushalt eines abgestorbenen Fichtenökosystems im Forellenbachgebiet des Nationalparks Bayerischer Wald. In: Nationalparkverwaltung Bayer. Wald, Bayer. Landesanstalt für Wald und Forstwirtschaft, Bayer. Landesamt für Wasserwirtschaft und Lehrstuhl für Bioklimatologie und Immissionsforschung der TU München (Hrsg.): Wasserhaushalt und Stoffbilanzen im naturnahen Einzugsgebiet Große Ohe 7; S BLASCHKE, M. (1998): Waldverjüngung in montanen und subalpinen Fichtenwäldern Mitteleuropas. Bayerische Landesanstalt für Wald. und Forstwirtschaft, Freising; 29 S. BRAUN-BLANQUET, J. (1964): Pflanzensoziologie. 3. Aufl., Springer, Wien; 865 S. BREIT, S. (1999): Waldentwicklung auf Sturmwurfflächen im Nationalpark Bayerischer Wald Unveröff. Dipl.arbeit an der TU München, Freising; 93 5., Anh. CLARK, P. J. und F. C. EVANS (1954): Distance to nearest neighbor as a measure of spatial relationships in populations. Ecology Vol. 35, No. 4; S DEUTSCHER WETTERDIENST (1999): Amtsblatt des Deutschen Wetterdienstes vom 27. Dezember Offen bach ; 8 S. EICHRODT, R. (1970): Über die Bedeutung von Moderholz für die natürliche Verjüngung im subalpinen Fichtenwald. Zeitschr. Schweiz. Forstvereins, Beiheft 45; 122 S. ELLENBERG, H., H. E. WEBER, R. DUELL, V. WIRTH, W. WERNER und D. PAULISSEN (1991): Zeigerwerte von Pflanzen in Mitteleuropa. Verlag Erich Goltze KG, Göttingen; 248 S. ELLING W., E. BAUER, G. KLEMM und H. KOCH (1976): Klima und Böden - Waldstandorte. Wiss. Schriftenr. Bayer. StMinELF 1; 150 S. ELLING W., E. BAUER, G. KLEMM und H. KOCH (1987): Klima und Böden - Waldstandorte. 2. Aufl., Wiss. Schriftenr. Bayer. StMinELF 1 ; 150 S. FALINSKY, J. B. (1976): Windwürfe als Faktor der Differenzierung und der Veränderung des Urwaldbiotops im Licht der Forschungen auf Dauerflächen. Phytocoenosis, Biuletyn Fitosocjologiczny, Warschau ; S FISCHER, A. (1987): Untersuchungen zur Populationsdynamik am Beginn von Sekundärsukzessionen. Dissertationes Botanicae 110, J. Cramer, Berlin; 234 S. FISCHER, A., G. ABS und F. LENZ (1990): Natürliche Entwicklung von Waldbeständen nach Windwurf. Ansätze einer "Urwaldforschung" in der Bundesrepublik. Forstw. Cbl. 109, Hamburg; S FISCHER, A. und H. JEHL (1999): Vegetationsentwicklung auf Sturmwurfflächen im Nationalpark Bayerischer Wald aus dem Jahre In: Forstl. Forschungsberichte. Schriftenr. Forstwiss. Fakultät der TU München und der Bayer. Landesanstalt für Wald und Forstwirtschaft; S FRAHM, J. P. und W. FREY (1983): Moosflora. Ulmer, Stuttgart, 522 S. FRANZ, F. und E. KENNEL (1973): Bayerische Waldinventur1970/71, Inventurabschnitt I: Großrauminventur, Basistabellen. Forstl. Forschungsberichte, München, 12; 346 S. GOEPPERT, H. R. (1868): Skizzen zur Kenntnis der Urwälder Schlesiens und Böhmens. Verlag Blochmann und Sohn, Dresden; 57 5., Anh. 94
94 HEURICH, M., REINElT, A. und FAHSE, L. (2001): Die Buchdruckermassenvermehrung im Nationalpark Bayerischer Wald. In: Nationa I pa rkverwa Itu ng Bayerischer Wa Id: Waldentwicklung im Bergwald nach Windwurf und Borkenkäferbefall ; S HEURICH, M. (2001): Waldentwicklung im montanen Fichtenwald nach großflächigem Buchdruckerbefall im Nationalpark Bayerischer Wald. In: Waldentwicklung im Bergwald nach Windwurf und Borkenkäferbefall. S HlllGARTER, F. W. (1971): Waldbauliehe und ertragskundliehe Untersuchungen im subalpinen Fichtenwald Scatle/Brigels. Diss. ETH Zürich; 80 S. HOFGAARD, A. (1993): Structure and regeneration patterns in a virgin Picea abies forest in northern Sweden. Journal of Vegetation Science 4; S HOMANN, M. und F. ENGELS (1991): Was kommt nach dem Sturm? Allg. Forst Zeitschrift, München, 12; S JEHl, H. (1995): Die Waldentwicklung auf Windwurfflächen im Nationalpark Bayerischer Wald. In: Nationalparkverwaltung Bayer. Wald (Hrsg.): Nationalpark Bayerischer Wald - 25 Jahre auf dem Weg zum Naturwald. Grafenau; S KAATZ, W. (1987): Kronenmerkmale und Jahrringbau an autochthonen Hochlagenfichten im Nationalpark Bayerischer Wald. Unveröff. Diplomarbeit an der FH Weihenstephan/FB Forstwirtschaft, Freising; 131 S. KARIUS, K. (1990): Orkanschäden ein Einzelfall? Allg. Forst Zeitschrift, München; S KENK, G., U. MENGES und R. BÜRGER (1991): Natürliche Wiederbewaldung nach Sturmwurf? Allg. Forst Zeitschrift, München 2; S KENNEL, E. (1973): Bayerische Waldinventur 1970/71. Inventurabschnitt I: Großrauminventur, Aufnahme- und Auswertungsverfahren. Forstl. Forschungsberichte, München; Nr.ll. KORPEl, S. (1995): Die Urwälder der Westkarpaten. G. Fischer Verlag, Stuttgart; 310 S. KRAMER, H. und A. AKCA (1995): Leitfaden zur Waldmesslehre. J. D. Sauerländer's Verlag, Frankfurt/Main; 298 S. LAESSIG, R. und W. SCHOENENBERGER (1993): Forschung auf Sturmschadensflächen in der Schweiz. Forst und Holz, 48. Jg. Nr.9; S LEIBUNDGUT, H. (1993): Europäische Urwälder: Wegweiser zur naturnahen Waldwirtschaft. Verlag Paul Haupt, Bern; 260 S. LORIMER, C. G. und L. E. FRELICH (1994): Natural Disturbance Regimes in Old-Growth Northern Hardwoods. Journal of Forestry; S LOZAN, J. L. (1998): Angewandte Statistik für Naturwissenschaftler. 2. Aufl., Pareys Studientexte 74, Blackwell-Wissenschafts-Verlag, Berlin; 280 S. MAI, W. (1998): Naturverjüngung auf Moderholz - Ergebnis einer Literaturstudie. Bayer. Landesanstalt für Wald und Forstwirtschaft, Freising; 59 S. MANTEL, W. (Hrsg., 1950): Ganghofers praktischer Holzrechner. 7. Aufl., Verlag der Schmid 'sehen Buchhandlung, Augsburg; 228 S. ' MAYER, H. (1983): Hemmnisse für den Gebirgswaldbau - Wege zur Überwindung. Allg. Forstzeitung, München Nr. 6; 5 S. 95
95 MEYER, P., J ACKERMANN, P. BALCAR, J BODDENBERG, R. DETSCH, B. FÖRSTER, H. FUCHS, B. HOFFMANN, W. KEITEL, M. KÖLBEL, C. KÖTHKE, H. Koss, W. UNKRIG, J WEBER und J WILLIG (2001): Untersuchung der Waldstruktur und ihrer Dynamik in Naturwaldreservaten. IHW Verlag, Eching; 107 S. NUESSLEIN, S. (1996): Hochlageninventur Bayerischer Wald Bayerische Landesanstalt für Wald und Forstwirtschaft Freising; 165. OBERDORFER, E. (1972): Süddeutsche Pflanzengesellschaften, Teil Aufl., G. Fischer Verlag, Stuttgart; 355 S. OBERDORFER, E. (Hrsg., 1992): Süddeutsche Pflanzengesellschaften, Teil IV: Wälder und Gebüsche (Text- und Tabellenband). 2. Aufl., G. Fischer Verlag, Stuttgart; 282 S. (Textb.), 580 S. (Tab.) On, E., M. FREHNER, H. U. FREY und P. LÜSCHER (1997): Gebirgsnadelwälder. Verlag Paul Haupt, Bern; 287 S. PALISAAR, J (1997): Untersuchungen zur Moosvegetation verschiedener Waldflächen des Nationalparks Bayerischer Wald nach Windwurf. Unveröff. Dipl.arbeit Philipps-Universität, Marburg; 87 S. PEnERSON, H. (1955): Die Massenproduktion des Nadelwaldes. Mitt. d. Forstl. Forsch.-Anstalt Schwedens, Bd. 45, H. 1 B (deutsch); PIELOU, E. C. (1977): Mathematical Ecology. John Wiley and Sons; 385 S. PRETZSCH, H. (1993): Analyse und Reproduktion räumlicher Bestandesstrukturen. Versuche mit dem Strukturgenerator STRUGEN. Schriften aus der Forstlichen Fakultät der Universität Göttingen und der Niedersächsischen Forstl. Versuchsanstalt, Bd. 114; 87 S. PRETZSCH, H. (1996): Strukturvielfalt als Ergebnis waldbaulichen Handeins. Allg. Forst- und Jagdzeitung, 167 (11); S PROJEKTGRUPPE NATURWALDRESERVATE (1993): Empfehlungen für die Einrichtung und Betreuung von Naturwaldreservaten in Deutschland. Forstarchiv 64; S PUTZ, F. E., P. D. COLEY, K. Lu, A. MONTALVO und A. AIELLO (1983): Uprooting and snapping of trees; structural determinants and ecological consequences. Can. J For. Res. Vol 13; REIF, A. und M. PRZYBILLA (1998): Regeneration der Fichte in den Hochlagen des Nationalparks Bayerischer Wald. AFZ/Der Wald, München 8; S ROST-SIEBERT, K. (1988): Ergebnisse vegetationskundlicher und bodenchemischer Vergleichsuntersuchungen zur Feststellung immissionsbedingter Veränderungen während der letzten Jahrzehnte. Ber. Forschungszentrums Waldökosysteme/Waldsterben, Reihe B, Bd. 8; 158 S. ROTHMALER, W. (Hrsg., 1990): Exkursionsflora von Deutschland. Bd. 4. Kritischer Band. 8. Aufl., Volk und Wissen Verlag, Berlin; 811 S. RonMANN, M. (1986): Wind- und Sturmschäden im Wald. J D. Sauerländer's Verlag, Frankfurt/Main; 128 S. SCHAETZL, R. J, S. F. BURNS, D. L. JOHNSON und T. W. SMALL (1989): Tree uprooting: review of impacts on forest ecology. In: Vegetatio 79, Kluwer Academic Publishers; S SCHERZlNGER, W. (1996): Naturschutz im Wald: Qualitätsziele einer dynamischen WaIdentwicklung. Verlag Eugen Ulmer, Stuttgart; 447 S. 96
96 SCHERZlNGER, W. (2000): Wilde Waldnatur. Der Nationalpark Bayerischer Wald auf dem Weg zur Waldwildnis. Nationalparkverwaltung Bayer. Wald (Hrsg.), Grafenau; 43 S. SCHMID, K. (1994): Natürliche Bestandesentwicklung auf Dauerbeobachtungsflächen von Windwürfen im Nationalpark Bayerischer Wald - fünf Jahre nach der Ersterhebung. Unveröff. Dipl.arbeit an der LMU München; 97 S. SCHOENENBERGER, W., H. KASPER und R. LAESSIG (1992): Forschungsprojekte zur Wiederbewaldung von Sturmschadenflächen. Schweiz. Z. Forstwesen, Zürich, 143(1992) 10; S SERNANDER, R. (1936): Granskär och Fiby urskog (The primitive forests of Granskär and Fiby, engl. Zus.fass.). Acta Phytogeographica Suecia 8; 232 S. SPURR, S. H. (1956): Natural Restocking of Forests Following the 1938 Hurricane in Central New England. Ecology 37(3); S STOECKLI, B. (1995): Moderholz für die Naturverjüngung im Bergwald. Wald und Holz 76, 16; S TRAUT, A. und W. WAGNER (1997): Beschreibung der wald baulich-ökologischen Situation auf einer nicht aufgearbeiteten Sturmwurffläche im Nationalpark Bayerischer Wald. Unveröff. Dipl.arbeit an der LMU München/Freising; 147 S., Anh. v. GADOW, K. und K. FUELDNER (1992): Bestandesbeschreibung in der Forsteinrichtung. Tagungsbericht der Arbeitsgruppe Forsteinrichtung, Klieken bei Dessau. VAN DER MAAREL, E. (1979): Transformation of cover-abundance values in phytosociology and its effects on community similarity. Vegetatio 39 (2); S WHITMORE, T. C. (1988): The Influence oftree Population Dynamics on Forest Species Composition. In: Plant Population Ecology, Oxford; 478 S. WILMANNS, O. (2. Aufl., 1993): Pflanzensoziolog ie. Quelle und Meyer, Stuttgart; 351 S. ZIERL, H. (1972): Der Hochwald. Untersuchungen über die Fichtenbestände in den Hochlagen des Bayerischen Waldes. Verlag Paul Parey, Hamburg; 80 S. 97
97 Abbildungsverzeichnis Archiv Nationalparkverwaltung : 7 Jehl:3, 5, 8, 9,10,14,16,17,23,42,45,50,53,54 Kiener: 47 Pöhlmann: S. 91 Scherzinger: 33, 34, 35, 36 Strunz: 1,2,4,21,22,24,48,49,51,52 Zellner: 6, 38 98
98 Waldentwicklung iin Inontanen Fichtenwald nach großflächigein Buchdruckerbefall iin Nationalpark Bayerischer Wald von Marco Heurich 1. Einleitung Standörtliche Besonderheiten der Hochlagen Die Geschichte des Bergfichtenwaldes Methoden Stichprobeninventur Kleinstrukturen Luftbildinterpretation Ergebnisse...: Entwicklung der Verjüngungsdichte Entwicklung der Baumartenanteile Entwicklung der Höhenstruktur Entwicklung der "Schäden" an der Verjüngung Schalenwild Rüsselkäfer Buchdrucker Schwarzer Schneeschimmel Schneebruch Verteilungsmuster der Verjüngung innerhalb der Bestände Klumpung der Verjüngung Kleinstrukturen der Waldbodenoberfläche Verteilungsmuster der Verjüngung im Gelände Bodenform Meereshöhe Exposition Geländeneigung Zeitpunkt des Absterbens der Altbestände Prägung durch die Altbestände Konkurrenzvegetation
99 5.7 Verteilungsmuster in den Hochlagen Überlebende Baumbestände Ameisenkolonien Diskussion Der alte Bergfichtenwald Sukzession - die "Erste" Sukzession - die "Zweite" "Schäden" an der Verjüngung Muster der Verjüngungsverteilung im Gelände Störungsregime hochmontaner Waldformationen Was bleibt vom alten Wald? Wie reagieren die Tiere? Verändern sich die Störungsmuster? Schlussfolgerungen für das Nationalpark-Management Zusammenfassung Summary Literaturverzeichnis
.")

100 Waldentwicklung im montanen Fichtenwald In Folge der Massenvermehrung des Buchdruckers Ups typographus) im Nationalpark Bayerischer Wald sind bisher Totholzflächen mit einer Ausdehnung von über ha entstanden. Der Schwerpunkt des Befalls liegt in den Fichtenbeständen der Hochlagen. Dort sind mittlerweile 85 % der ursprünglichen Bestockung abgestorben (HEURICH etal. 2001). Für viele Nationalparkbesucher und für große Teile der einheimischen Bevölkerung war und ist es immer noch schwer zu verstehen, warum die ganzen Bäume sterben mussten. Hätte man nicht durch rechtzeitige Borkenkäferbekämpfung den charakteristischen Hochwald erhalten können? Jetzt aber breiten sich große, zunächst unästhetisch anmutende Totholzflächen über weite Teile des Nationalparks aus. Für viele Menschen ist durch den Verlust der alten Wälder auch ein Stück Heimat verloren gegangen. Nach anfangs heftigen Diskussionen über notwendige Maßnahmen zur Käferbekämpfung wurde mit der Vergrößerung der Randzone zum Schutz der dem Nationalpark benachbarten Wälder ein Kompromiss gefunden. Danach rückte immer mehr die Frage, ob und wie sich der Wald auf den betroffenen Flächen entwickelt, in den Mittelpunkt der Diskussion. "Ist der Fortbestand des Waldes gesichert? Wird es größere waldfreie Flächen geben? Wie lange dauert es, bis sich ein neuer Wald entwickelt? Wie wird der neue Wald aussehen? Ist es notwendig, der Natur mit Stützungsmaßnahmen unter die Arme zu greifen?", sind einige zentrale Fragen. Für die Höhenstufen des Aufichtenwaldes und des Bergmischwaldes konnte die Nationalparkverwaltung anharid einer ganzen Reihe von Beispiels- und Dauerbeobachtungsflächen zeigen, dass die Wälder dort über ein ausreichendes natü rl iches Regenerationsvermögen verfügen (FISCHER 1990, FISCHER und JEHL 1999, JEHL 1995). In den Wäldern der Hochlagen stellt sich die Situation, insbesondere wegen des sehr schnellen und großflächigen Absterbens der Altbäume und der ungünstigen Standortsbedingungen, nicht so eindeutig dar. Dass die Fragen der Bevölkerung durchaus berechtigt sind, zeigt auch die in Forstfachkreisen schon seit dem vorletzten Jahrhundert z. T. sehr heftig geführte Auseinandersetzung über die richtige Waldbehandlung im Hochwald. Dreh- und Angelpunkt dieser Diskussion war die Frage, wie eine ausreichende Verjüngung der Bestände gewährleistet werden kann. Auch Abb.l Abteilung Ebengfeichtet {74. Juni 200 7}. 101
101 heute gehen die Meinungen darüber, ob und wenn ja welche Maßnahmen zur Walderhaltung notwendig sind, weit auseinander. Während die Nationalparkgegner fordern, die toten Bäume aufzuarbeiten und als Brennholz zu verkaufen, um anschließend die Flächen aufforsten zu können, ist es das Ziel der Nationalparkverwaltung, entsprechend den Vorgaben der internationalen Naturschutzunion (IUCN) nicht lenkend in die natürlichen Abläufe einzugreifen. Als Kompromiss wurde eine Formel gefunden, die beide Lager zufrieden stellte: Nur wenn sich längerfristig auf größerer Fläche keine Verjüngung etabliert, muss die natürliche Entwicklung durch Pflanzungen unterstützt werden. Damit bekam die Nationalparkverwaltung auch den Auftrag, die Entwicklung der Waldverjüngung wissenschaftlich zu beobachten. Aus Sicht der Forschung bot sich ein höchst interessantes Studienobjekt. Es liegen zwar bereits Studien über die Regeneration von naturnahen Fichtenökosystemen nach Störungen vor, diese stammen aber meist aus Nordamerika oder aus relativ kleinen Reservaten (PERRY 1997, ROMME und DESPAINE 1989, RYKIEL 1988, TURNER et al. 1997, LAESSIG et al. 1995, WERMELINGER et al. 1995). Erfahrungen über eine nicht vom Menschen beeinflusste Entwicklung nach großflächigem Absterben der Altbäume und ihre Auswirkungen auf die Lebensgemeinschaften konnten im mitteleuropäischen Raum bisher noch nicht gesammelt werden, da Borkenkäfermassenvermehrungen in bewirtschafteten Wäldern bekämpft und die dabei entstandenen Kahlflächen umgehend aufgeforstet werden. Um die einzigartige Chance zu nutzen, die Veränderungen im Waldökosystem zu dokumentieren, wurde eine ganze Reihe von Forschungsprojekten ins Leben gerufen. Dazu gehören unter anderem Untersuchungen zur Entwicklung derverjüngung, über den Sameneintrag aus größeren Distanzen, über den Einfluss auf die Böden und den Wasserhaushalt und über die Veränderungen in den Lebensgemei nschaften. Schwerpunkt der vorliegenden Arbeit ist es, die Entwicklung der Waldverjüngung in der Hochlagenstufe zu dokumentieren. Dafür werden seit 1996 in zweijährigem Abstand 572 Probeflächen untersucht, die gleichmäßig über die gesamten Hochlagen verteilt sind. Als Referenz für die Ergebnisse dienen die Daten der Waldinventur von Damals waren die Wälder zwischen Rachel und Lusen noch zum überwiegenden Teil geschlossen. Von großem politischen und wissenschaftlichen Interesse ist insbesondere die zeitliche und räumliche Entwicklung der Verjüngungsdichte und die Analyse der sie beeinflussenden Faktoren. Darüber hinaus wird auch.der aktuelle Kenntnisstand über den Einfluss der Absterbeprozesse auf die Tierwelt diskutiert. 102



 in den organischen Auflagen und bis 4,5 in den")
102 2. Standörtliche Besonderheiten Der Hochlagen Die Hochlagen oder auch hochmontane Höhenstufe des Inneren Bayerischen Waldes weist relativ ungünstige Bedingungen für di~ Entwicklung der Waldverjüngung auf. In einer Meereshöhe zwischen 1050 mund 1453 m stellt die Temperatur einen Minimumfaktor für das Wachstum der Pflanzen dar. Die Jahresdwchschnittstemperatur beträgt hier nur 3,0 bis 4,5 C (ELLING et al. 1987). Die hohen Niederschläge von 1300 bis 1800 mm und ihre Verteilung, mit einem Hauptmaximum im Sommer und einern zweiten Maximum im Winter, führen zu einer im allgemeinen günstigen Wasserversorgung in der kurzen Vegetationsperiode. 50 0J0 der Niederschläge fallen als Schnee, was in Kombination mit den geringen Temperaturen eine mittlere Schneedeckenzeit von sieben Monaten, also von Oktober bis Mai, zur Folge hat. Vorherrschende Bodentypen sind Braunerden mit mehr oder weniger ausgeprägten Podsolierungs"': erscheinungen, die aus Verwitterungsmaterial der anstehenden Granite und Gneise entstanden sind. Wie auf Grund der Basenarmut dieser Ausgangsgesteine zu erwarten ist, sind die Böden sauer bis sehr sauer und weisen ph-werte von 2,7 bis 2,8 (in KCI) in den organischen Auflagen und bis 4,5 in den tieferen Bodenhorizonten auf. Ein Kennzeichen dieser Böden ist eine Schicht von verfestigtem Schutt in cm Tiefe. Dieser ist selbst mit Werkzeugen nur schwer zu bearbeiten und reduziert die Durchwurzelungstiefe und die Wasserdurchlässigkeit. Insbesondere das Wachstum der Rotbuche (Fagus sylvatica) wird unter diesen Voraussetzungen gehemmt, so dass sie sich auf verfestigem Schutt nur zu Kümmerformen entwickeln kann. Der "mittelgründige Lehm über verfestigtem Schutt" ist mit einem Anteil von 57 0J0 die häufigste Bodenform der Hochlagen. Vor allem auf Grund der hohen Niederschläge findet man auf 12 0J0 der Hochlagenfläche mineralische und organische Nassböden. In nennenswertem Umfang (ebenfalls 12 0J0) treten auch Fels- und Blockböden auf. Sie konzentrieren sich auf steilere Lagen und sind durch das flächige Auftreten von Fels und Grobskelett gekennzeichnet. Die Durchwurzelung beschränkt sich auf diesen Böden auf das mineralische Füllmaterial zwischen den Blöcken und auf eine mehr oder minder mächtige Rohhumusauflage. Die Verteilung der. wichtigsten Bodengruppen ist in Karte 2 dargestellt. Karte 1 Orohydrografische Karte des Rache/-Lusen-Gebietes. Die Hochlagen sind mit einer weißen Linie abgegrenzt. Abb.2 Schneehöhen von bis zu 3 m sind keine Seltenheit in den Hochlagen. 103

. Die räumliche Abgrenzung der Hochlagen erfolgte durch ELLING et al.")

103 Block - Humus - Böden Lehm organische und rrineralische Naßböden rritlelglündiger Lehm überverfestg~m Schutt 100iiiO_==:::i0 1i1001:0===5~O M:ta- Karte 2 Verteilung der zu vier ökologischen Gruppen zusammengefassten Boden formen in den Hochlagen. Abb. 3 Die Bergmischwälder der Hanglagen werden oberhalb des Hangknicks durch die Fichten wälder des Böhmerwaldkammes abgelöst. Unter diesen ungünstigen klimatischen und edaphischen Bedingungen bleibt die Mineralisation der anfallenden ober- und unterirdischen Streu unvollständig, so dass sich 2 bis 19 cm, im Durchschnitt 8 cm mächtige Auflagehorizonte entwickeln, die typischerweise als Rohhumus und rohhumusartiger Modervorliegen (ELLING et al. 1987). Die räumliche Abgrenzung der Hochlagen erfolgte durch ELLING et al. (1987) nach geländemorphologischen und edaphischen Kriterien: Die Hochlagen beginnen dort, wo die relativ steilen Hänge mit einem Hangknick oder einer Wölbung in das sanfte Gelände des Böhmerwaldkammes übergehen. Die Dauer der Schneebedeckung ist daher oberhalb des Hangknicks deutlich länger als unterhalb. Es handelt sich zugleich um den Bereich, in dem der Bergmischwald der Hanglagen dem Bergfichtenwald weicht. Die genaue Festlegung der Hochlagen erfolgte nach einem Merkmal des Bodens, nämlich nach der Obergrenze der Verbreitung der Lockerbraunerden. Sie überschreitet jedoch nicht 1250 m Meereshöhe. 104
104 3. Die Geschichte Des Bergfichtenwaldes Untersuchungen von TRAUTMANN (1952) weisen darauf hin, dass sich die reinen Fichtenwälder in den Hochlagen erst im Verlauf des 14. Jahrhunderts aus Bergmischwäldern entwickelt haben. Allerdings kam die Fichte seit der Kiefern Haselzeit (6500 bis 5500 v. ehr.) in dieser Höhenstufe kontinuierlich vor und hatte immer einen höheren Anteil als in den tieferen Stufen (FIRBAS 1952). Auf Grund der späten Besiedelung des Gebietes - die ersten Ortschaften um das Kloster St. Oswald werden 1396 erwähnt - scheint es sehr unwahrscheinlich, dass dieser Baumartenwechsel durch den Menschen verursacht wurde. Auslöser für das Zurückweichen von Rotbuche und Tanne war vermutlich die Abkühlung des Klimas während des Hochmittelalters. Die ersten Glashütten wurden im 16. Jahrhundert angelegt, ihr Einfluss blieb aber zunächst auf die Tallagen beschränkt. Erst im 17. Jahrhundert drang man tiefer in die Wälder vor. Welchen Einfluss der Holzbedarf der Glashütten auf den Hochwald hatte, lässt sich nur schwer abschätzen. Als Konsens unter den Forsthistorikern gilt, dass die Hochlagen vor allem zur Gewinnung von Pottasche (Schmelzpunkterniedrigung bei der Glaserzeugung) genutzt wurden, da sich diese im Gegensatz zu dem ebenfalls von den Glashütten benötigten Brennholz leichter aus den abgelegenen Gebieten abtransportieren ließ. Die zur Pottaschegewinnung ausgewählten Bäume mussten vor allem stark und alt sein, dabei war es unwichtig, ob sie bereits abgestorben waren oder noch lebten. Sie wurden meist an Ort und Stelle verbrannt, egal ob noch stehend oder bereits am Boden liegend. Auf Grund ihres hohen Kaligehaltes wurde vor allem die Rotbuche, weniger die Tanne und in geringerem Maße die Fichte "geaschelt". Als die Wälder in der Mitte des 19. Jahrhunderts in Staatsbesitz überwechselten, waren sie vermutlich in einem sehr naturnahen Zustand: Die ersten primitiven Forstoperate dieser Zeit sprechen jedenfalls alle von "Urwald, welcher der Ausnützung bisher zu fern gewesen war". Erst nach dem Ausbau des Triftsystems (lizer Trift-Komplex, 1836/37) konnten auch die Bäume aus den Hochlagen zu den Orten des Holzverbrauches gebracht werden. Die sich anschli.eßende Zeitspanne der regulären Waldnutzung wurde von ZIERL (1972) als "Periode der Einzelplenterung" bezeichnet. Die wesentlichen Ziele und Behandlungsgrundsätze dieser Zeit wurden bereits in einer Besprechung zwischen dem königlichen Ministerialkommissär Waldmann und den Amtsvorständen der Forstämter des Inneren Bayerischen Waldes im Jahre 1840 formuliert: Die "hochgelegenen Fichtenwaldungen" eignen sich "nicht zum schlagweisen Betrieb, vielmehr sollten sie mit großer Sorgfalt in periodisch wiederkehrenden Plenterhieben genutzt werden. (...) Das Hauptaugenmerk wolle man vorerst auf die Reinigung der betreffenden Waldungen von dem noch brauchbaren dürren und abständigen Holze richten. Erst am Schlusse dieser Operation gewinne die Entscheidung der Frage ihre entsprechende Wichtigkeit, ein Zeitpunkt, der übrigens noch sehr entfernt liegt". Das Ergebnis dieser Besprechung findet sich dann auch in den 1849 aufgestellten Wirtschaftregeln für den Bayerischen Wald wieder. Nach einer Auswertung der anschließend gefertigten Forsteinrichtungswerke kommt ZIERL (1972) zu dem Schluss, dass es zu den eigentlichen Plenterhieben mit dem Ziel, das Auflaufen und die Weiterentwicklung der Verjüngung zu sichern, nicht in ausreichendem Umfang gekommen ist. "Es scheinen überwiegend Reinigungsund Auszugshiebe geführt worden zu sein". Insgesamt hatte die Holzproduktion in den Hochlagen keine besondere Bedeutung. Nebennutzungen wie die Waldweide waren da wichtiger. Die Hochlagen waren eine geschätzte Viehweide. 105



105 In der Zeit von Pfingsten bis Michaeli (29. September) weideten dort große Herden mit jungen Ochsen (ZIERL 1972). Als Beispiel für die hohe Weidebelastung kann das Weiderecht der Stadt Grafenau angesehen werden. In ihrem Weidedistrikt nördlich von Waldhäuser, an der Grenze zum heutigen Tschechien, durften auf einer Fläche von 422 ha rund 80 Waldstiere weiden (SEYFERT 1975). Die Verjüngung von Laubbäumen wird unterdiesen Bedingungen wohl sehrschwierig gewesen sein und 1870 folgten zwei Sturm katastrophen, die, zusammen mit der nachfolgenden, bis 1876 andauernden Borkenkäferkalamität, ein ge-zieltes forstwirtschaftliches Arbeiten auf Jahre hinaus unmöglich machten. Insgesamt entstanden damals im Rachel-Lusen-Gebiet durch die Aufarbeitung der Windwürfe und die Bekämpfung des Borkenkäfers Kahlflächen mit einer Ausdehnung von ha, alleine 870 ha davon in den Hochlagen (ELLING et al. 1987). Gängige Methode der Wiederbestockung dieser geräumten Flächen war die Pflanzung, die Saat spielte nur eine untergeordnete Rolle. Je Hektar wurden bis Pflanzen ausgebracht. Zusammen mit den immer wieder notwendigen Nachpflanzungen dauerte es 30 Jahre, bis die Aufforstungen der Kahlflächen abgeschlossen waren. Ein Erbe aus dieser Zeit ist es, dass während der Aufforstungen nicht auf die Herkunft des Saat- und Pflanzgutes geachtet wurde und sich deshalb bis heute eine größere Anzahl nicht autochthoner Fichten in den Hochlagen befindet. Abb. 4 In der Vergangenheit tra ten immer wieder heftige Stürme au f, die immense Schäden in den Wäldern des Bayerischen Waldes veru rsachten. Windwurf im Juli In den zwanziger und dreißiger Jahren des 20. Jahrhunderts kam dann das Ende der Einzelplenterung. Grund hierfür war vor allem, dass es mit diesem Waldbauverfahren nicht gelungen war, eine ausreichende Verjüngung der Bestände zu erreichen. Vielmehr wurden die ohnehin weitständigen Hochlagenbestände durch die Einzelplenterung weiter aufgelichtet, was zwar den Graswuchs förderte, aber für eine zufriedensteilende Entwicklung der Verjüngung noch nicht ausreichte. REBEL (1922) hat diesen Zustand treffend charakterisiert, indem er schrieb: "Jungwuchs geht hier nur in die Höhe, wenn es an zwei Dingen nicht fehlt: an Seitenschutz und Wärme. Gleichmäßiges Auflichten der ohnehin schon lichten Bestände nützt gar nichts, kann vielmehr alles verderben". Auf Grund dieser Erfahrungen führte man den Femelschlag als Waldbauverfah ren für die Hochlagenwälder ein. Als aber auch dieser keine zufriedenstellende Lösung dieses "Verjüngungsproblems" brachte, 106

106 versuchte man mit einer bis auf den Mineralboden reichenden Bodenbearbeitung und einer "markanten Femelung" mit mindestens 0,1 ha großen Lücken bessere Ergebnisse zu erzielen (BUETINER 1950). So wurde während der Fichtenvollmast 1958 der Scheibenpflug zur Vorbereitung des Saatbetts eingesetzt und 20 Zentner Kalk je Hektar eingebracht. In den Folgejahren wurde die Verjüngung von Hand ausgegrast und teilweise auch Herbizide verwendet. Zur Stärkung bekamen die Fichten zwei Jahre lang den Dünger "Nitrophoska blau". Dieser wurde mit Hilfe eines Löffels an jeder Pflanze ausgebracht. Zunächst stellte sich auf den vorbereiteten Flächen reichlich Fichtenverjüngung ein, auch von einer "freudigen" Verjüngung von Bergahorn (Acer pseudoplo- tonus) und Vogelbeere (Sorbus oucuporio) - allerdings hinter Zaun - wird berichtet (BUELOW 1964). Trotz des hohen Aufwandes und der anfänglichen Erfolge war einige Jahre später der Großteil der Verjüngung bereits wieder verschwunden (FRITSCH, pers. Mitteilung). Die von ZIERL (1972) empfohlene Gruppenplenterung kam im Untersuchungsgebiet nicht mehr zur Anwendung, da die Bestände in den Hochlagen die ersten Bereiche des Nationalparkes waren, die in Hiebsruhe gestellt wurden. Seit der Forstplanung von 1972 finden dort keine forstlichen Eingriffe mehr statt. Spätestens nach den Windwurfereignissen 1983/84, und dem daran anschließenden Borkenkäferbefall, der bis heute anhält, verloren die Hochiagenwälder auf großen Flächen ihre forstliche Prägung und konnten sich nach eigenen Gesetzmäßigkeiten entwickeln. Eine genaue Beschreibung der Totholzflächenentwicklung findet sich in NUESSLEIN (1999) und HEURICH etal.(2001). Windwurf _ 1983 und 1984 Windwurf und Buchdruckerbefall _ nach 1868 _ nach OOiiO_==jO.1 Oi::0===5~'O ~er Karte 3 Große Windwürfe und Borkenkäferereignisse in den Hochlagen (verändert nach EWNGetal. 1987). 107




107 4.1 Stichprobeninventur Die Außenaufnahmen fanden in den Monaten Juni und Juli 2000 statt und wurden von vier Aufnahmetrupps mit je zwei Forstwirten durchgeführt. Ein weiterer Aufnahmetrupp wurde vorwiegend zu r Qualitätssicherung eingesetzt. Das Aufnahmekonzept der vergangenen Jahre wurde beibehalten, um die Vergleichbarkeit der /'... ~ 500 m' Ergebnisse zu gewährleisten. Dadurch wird es möglich, die Entwicklungsdynamik der Verjüngung zu beschreiben. Da der Schwerpunkt der Abb.5 Aufnahmedesign Abb.6 Die Fortbe wegung fü r die Trupps in den To tholzflöchen ist sehr anstrengend. Tab. 7 Höhenklassifizierung der Verjüngung. Grundlage für die Erhebung der Verjüngung ist das 1991 für die Waldinventur angelegte Stichprobennetz. Die permanenten Aufnahmepunkte sind systematisch in Form eines Quadratrasters mit 200 m Seitenlänge über die ganze Fläche des Nationalparks verteilt und im Gelände mit Rohrmarken und Eisenstäben vermarkt (Abbildung 5). Auf Grund der spezifischen Fragestellung werden die Daten, wie bereits 1996 und 1998, nur auf den 572 Inventurpunkten im Hochlagenbereich des Rachel-Lusen-Gebietes erhoben. Klasse Höhe in cm >= 300 Untersuchungen auf der Waldverjüngung liegt, werden nur die Bäume zwischen 10 cm und 5 m Größe auf konzentrischen Probekreisen mit gestaffelten Kluppschwellen erfasst. Das heißt, für die Bäume von 0 bis 5,9 cm Brusthöhendurchmesser (BHD) beträgt die Probekreisfläche 25 m' und für die Bäume von 6,0 bis 11,9 cm BHD 50 m'. Bei jedem Baum werden die Art, evtl. aufgetretene Schäden und die Höhenklasse erfasst. Das Probeflächendesign und die Aufnahme der Parameter entsprechen somit dem in der Bayerischen Staatsforstverwaltung eingesetzten Standardinventurverfahren (BAYERISCHES STAATSMINISTERIUM FUER LANDWIRTSCHAFT UND FORSTEN 1982). 108


108 4.2 Kleinstrukturen Um die Bedeutung von Kleinstrukturen für die Verjüngung und deren Entwicklung zu untersuchen, wurden die Flächenanteile der aspektbestimmenden Kleinstrukturen auf den Probeflächen der Hochlagen erfasst. Insgesamt wurden zwölf Kleinstrukturen differenziert, die im Folgenden beschrieben werden: Moderholz Diese Kleinstruktur umfasst Stubben oder liegendes Totholz ab einem Durchmesser von mehr als 10 cm. Bewuchs auf Moderholz, wie z. B. Moos oder Heidelbeere wird nicht berücksichtigt. Moderholz wird nur ausgeschieden, wenn der Stamm oder die Stammteile auf dem Boden aufliegen. Baumkontakt Die unmittelbare Umgebung von liegendem Totholz mit mehr als 10 cm Durchmesser wird als eigener Kleinstandort ausgeschieden. Diese Kleinstruktur grenzt an das Holz an und reicht bis in eine Entfernung, die dem halben Durchmesser des Stammes entspricht. Baumkontakt wird nur ausgeschieden, wenn der Stamm oder Stammteile auf dem Boden aufliegen. Zu dieser Kleinstruktur zählt auch der Bereich um Kategorie zugerechnet. Dieses Kleinstruktur wird ausgewiesen, wenn das Material das Wachstum der krautigen Vegetation hemmt und gleichzeitig das Wachstum der Verjüngungspflanzen möglich ist (Mulcheffekt). also mehr als zwei Drittel bis drei Viertel des Bodens bedeckt werden. Bewachsener Felsen Mit Vegetation überwachsene Felsen werden als eigene Kleinstruktur ausgeschieden. Hierbei spielt es keine Rolle, welche Pflanzenart auf dem Fels wächst. Liegt der Fels ebenerdig, wird diese Kleinstruktur nicht ausgeschieden. Die Feisteile müssen mindestens einen Durchmesservon 10 cm aufweisen. Abb. 7 Nach dem Zusammenbruch der Altbestände bildet sich auf dem Waldboden ein vielfältiges Standortsmosaik. die Stockachseln von stehendem Totholz und lebenden Bäumen, wenn diese einen Durchmesser von über 10 cm aufweisen. Reisig/Rinde Insbesondere um stehendes Totholz können sich herabfallendes Reisig und Rinde anhäufen. Kronenteile und Stammstücke mit einem Durchmesser von weniger als 10 cm werden dieser Wurzelteller Zur Kleinstruktur Wurzelteller gehören neben dem aufgeklappten Wurzelteller selbst auch die vom Wurzelstock ausgehobene Bodenmulde, solange dort der Mineralboden noch offen liegt. Humus (unbewachsener Auflagehumus) Diese Kleinstruktur besteht aus sich zersetzenden Nadeln, Blättern, Flechten und Feinreisig und liegt dem Mineralboden auf. Der Zersetzungsgrad reicht von noch unzersetzter Streu bis hin zum feinstrukturierten Humus. Sie weist keinen Bewuchs auf. Abb.8 Abgrenzung der Kleinstruktur Baumkontakt. 109
 und der Breitblättrige Dornfa rn (Dryopteris dilatata). Abb.9 Typische Ausprägung der Kleinstruktur Reisig/Rinde.")

109 ~ ~ ~ Gras-Fazies Die dominierenden Arten sind Berg-Reitgras (Calamagrostis villosa), Drahtschmiele (Deschampsia flexuosa) und Wald-Hainsimse (Luzula sylvatica). Farn-Fazies Die häufigsten Arten sind der Alpenfrauenfarn (Athyrium distentifolium) und der Breitblättrige Dornfa rn (Dryopteris dilatata). Abb.9 Typische Ausprägung der Kleinstruktur Reisig/Rinde. Tab. 2 Schlüssel für die Ansprache der Flächenanteile der Kleinstrukturen. Zwergstrauch-Fazies Die dominierende Art ist die Heidelbeere (Vaccinium myrtillus). Moos-Fazies Moosstandorte werden ausgewiesen, wenn die Moose auf dem Boden, nicht aber auf Felsen oder Totholz wachsen. Insgesamt wurden über 30 Moosarten zu dieser Kleinstruktur zusammengefasst. Die häufigsten Moose sind Schönes Frauenhaarmoos (Polytrichum formosum) und Besengabelzahnmoos (Dicranum scoparium). Bärlapp-Fazies Die am häufigsten vorkommende Art ist der Sprossende Bärlapp (Lycopodium annotinum). Kräuter und Himbeere Vorkommende Kräuter sind z. B.: Grüner Alpenlattich (Homogyne alpina), Soldanelle (Soldanella montanal, Siebenstern (Trientalis europaeal, Sauerklee (Oxalis acetosellal, Schattenblümchen (Maianthemum bifolium). Zum Ausscheiden dieser Kleinstruktur müssen diese Pflanzenarten mindestens des Bodens bedecken. Die Kleinstrukturen werden in der 25 m' großen Probefläche des permanenten Inventurpunktes erfasst. Ihre Flächenanteile werden in 10 lo-stufen angeschätzt. Bei Flächenanteilen kleiner erfolgt eine Unterteilung in die Stufen und 6-10 %. Kleinstandorte mit sehr geringer Flächenausdehnung «1 %, z. B. ein kleiner Baumstumpf) werden mit einem "+" notiert. Die Summe der Schlüsselnummern muss immer 1 0 ergeben. Liegt die Summe leicht über oder unter diesem Wert, werden die Flächen der Kleinstrukturen entsprechend ab- bzw. aufgestuft. Bei der Inventur 1996 wurde zusätzlich für jede Pflanze bestimmt, in welcher Kleinstruktur sie wächst _. ~- ~--.. ~. Fläche Schlüssel-Nr. geringe Fläche + 1-5% % % % % % / % % / % X 110
.")


110 4.3 Luftbildinterpretation Die Entwicklung der. Altbestände wird durch regelmäßige Luftbild-Befliegungen im Intervall von einem Jahr dokumentiert. Dabei werden Color-Infra rot-luftbilder im Maßstab von 1 : bis 1 : erstellt (HEURICH 2001). Um die Bearbeitung der Luftbilder zu erleichtern, wurden sie mit einem Liesegang-Antiskop auf eine Kartengrundlage mit dem Maßstab 1 : projiziert und anschließend die verbliebenen Bestände und Einzelbäume kartiert und digitalisiert. Grundlage für die Kartierung waren die Luftbilder der Befliegung vom Herbst Folgende Klassen wurden in Anhalt an die Definition der Waldentwicklungsstadien nach RALL (1995) bei der Luftbildinterpretation ausgeschieden (Tabelle 3). Da eine eindeutige Klassifikation auf Grund des Luftbildes nicht immer möglich war, wurde die terrestrische Kartierung der Waldentwicklungsstadien aus dem Jahr 1992 bei der Interpretation mit berücksichtigt. Fichte-Jugendstadium, Fichte-Wachstumsstadium, Fichte-Einzelbaum (es wurden nur die EinzeIbäume erfasst, die mindestens dem Mittelstand zugerechnet werden können), Laubholz-Einzelbaum (Es wurden nur die Einzelbäume erfasst, die mindestens dem Mittelstand zugerechnet werden können), Laubholz-Wachstumsstadium, Laubholz-Reifungsstadium, frischer Käferbefall. Abb. 10 CIR-Luftaufnahme vom Lusen. Rot = lebende Bestände Grau = abgestorbene Bestände. 111

 (lebende) Bestandsglieder räumig (sehr licht). Al Stehendes Totholz ist von der Menge her ZS weit geringer als Lebendholz + A2 Bestandsglieder räumig (sehr licht).")


111 Tab. 3 Fragenabfolge zur Kartierung der Waldentwicklungsstadien O. Ist die Fläche bestockt? 1. Entspricht Fläche nach alter Einordnung JP? Stehendes und/oder liegendes Totholz kann vorhanden sein, wenn ja, dieses als Besonderheit einpunktieren 2. Sind auf der Fläche überstand und Verjüngung unterscheidbar? nein : Blöße (unterbestockt), wenn stehendes Totholz vorhanden + MS ja: weiter mit Ziffer 1 ja: JS nein: weiter mit Ziffer 2 nein: Fallgruppe A ja: Fallgruppe B Fallgruppe A: nur Oberstand (ohne Verjüngung) (lebende) Bestandsglieder räumig (sehr licht). Al Stehendes Totholz ist von der Menge her ZS weit geringer als Lebendholz + A2 Bestandsglieder räumig (sehr licht). Stehendes Totholz ist von der Menge her weit mehr als Lebendholz + MS A3 Wenn Bestand mindestens licht geschlossen: mittl. BHD 8-25 cm + WS A4 mittl. BHD > 25 cm und ein - oder zweifache Höhenschichtung + RS A5 mittl. BHD > 25 cm und dreifache Höhenschichtung + PS Fallgruppe B: Oberstand mit Verjüngung lebende Bestandsglieder vereinzelt stehend (ähnlich Bl Überhälter) oder nicht vorhanden und nicht nur vereinzelt MS stehendes Totholz über spärlicher Verjüngung + B2 wenn Bestand mindestens licht geschlossen: überstand mittl. BHD < 25 cm WS B3 überstand mittl. BHD >25 cm und einsch ichtig über Verjüngung + VS B4 überstand mittl. BHD > 25 cm und zweischichtig über Verjüngung PS Abkürzungen: JS = Jugendstadium, RS = Reifestadium, ZS = Zerfallsstadium, JP = Jugendpflege, VS = Verjüngungsstadium, MS = Mortalstadium, PS= Plenterstadium (Größe der zu beurteilenden Fläche gleichen Charakters> 0,1 halo 112





112 113
")



113 114 (Abb. 1 7)

. Damit können Pflanzen größer 20 cm als etabliert angesehen werden.")

114 5.1 Entwicklung der Verjüngungsdichte Von Bedeutung ist vor allem die Entwicklung bei den Pflanzen, die bereits als etabliert angesehen werden können, bei denen also nur noch geringe Ausfälle zu erwarten sind. Während bei frisch gekeimten Individuen die Verluste z. B. durch Trockenheit noch sehr groß sind, weisen Untersuchungen aus dem Nationalpark darauf hin, dass Pflanzen ab einer Größe von 20 cm eine hohe Überlebenswahrscheinlichkeit besitzen. Die Ausfallquote in der Schicht zwischen 20 und 40 cm liegt bei nur etwa 5-10 % (BAUER pers. Mitteilung, JEHL pers. Mitteilung). Damit können Pflanzen größer 20 cm als etabliert angesehen werden. Abbildung 18 zeigt die Entwicklung der durchschnittlichen Zahlen etablierter Individuen von 1991 bis Nachdem die Zahlen zwischen 1991 und 1996 nur leicht von 978 auf Pflanzen je Hektar zugenommen hatten, ergab die Inventur 1998 einen deutlichen Anstieg auf Pflanzen je Hektar. Bis zum Jahr 2000 nahm die Anzahl der Pflanzen um über 60 % auf nunmehr Pflanzen je Hektar zu. Die Aufschlüsselung nach Baumarten zeigt, dass dieser Anstieg fast ausschließlich durch die Fichte bedingt ist. Die Vogelbeeren haben zwar auch um über 30 % zugenommen, dies fällt jedoch mit 100 Pflanzen je Hektar zahlenmäßig kaum ins Gewicht. Bei allen anderen Baumarten konnte eine Zunahme der durchschnittlichen Pflanzenzahlen je Hektar nicht beobachtet werden. Im Jahr 2000 wurden hierim Mittel 681ndividuenje Hektargezählt. Bezieht man darüber hinaus die Pflanzen zwischen 10 und 20 cm in die Analyse mit ein, wird der Anstieg zwischen 1998 und 2000 noch eindrucksvoller: Die Gesamtzahl der erfassten Pflanzen hat sich seit 1998 von auf Pflanzen je Hektar fast verdoppelt. Die Dominanz der Fichte ist hier noch stärker ausgeprägt als bei dem Kollektiv größer 20 cm. Ihr Anteil konnte sich 2500, _-, 2000 ~..s:::: u :0 ~1000 -~::------~~========:;" ::3 Cl c: :::3 ~ (Q..s:::: -=-! (Q..s:::: -=li= ~ 2500 u +'..s:::: u : '" Cl c: ::3 Cl 1500 c: :::3 '':- ~ Pflanzen größer 10 cm Jahr Fichte Vogelbeere o Sonstige Baumarten mehr als verdoppeln. Die Anzahl der Vogelbeeren stieg in diesem Stratum ebenfalls um über 30 % auf nun 436 Pflanzen je Hektar. Auch die Anzahl der Rotbuchen erhöhte sich stark, sie stieg um 60 %, erreichte damit aber nur ein Niveau, weiches sie bereits 1996 hatte. Abb. 18 Entwicklung der mittleren Verjüngungsdichte der etablierten Pflanzen. 115




115 0 5.2 Entwicklung der Baurnartenanteile Baumart Inventur 1991 N/ha 0/0 Fichte ,7 latsche 7 0,7 Buche 32 3,3 Bergahorn Sandbirke Vogelbeere ,0 I- Weide 14 1,4 Gesamt ,0 Inventur 1996 Inventur 1998 Inventur 2000 N/ha 0/0 N/ha 0/0 N/ha 0/ , , ,3 1 0,1 1 0,1 1 0, ,2 61 5,1 55 2,9 ; ,4 1 0,1 2 0,1-1 0,1 2 0,2 3 0, , , ,2 20 1,8 2 0,2 7 0, , , ,0 anderen Baumarten auf 0,5 0/0. Erst bei der Inventur 2000 ergab sich durch den Anstieg des Fichtenanteils ein verändertes Bild. Der Anteil der Vogelbeere sank, obwohl die absoluten Zahlen um mehr als ein Drittel anstiegen, von 22,4 auf 19,2 Ofo. Auch der Anteil der Rotbuche ging wieder von 5,1 auf2,9 Ofo zurück. Alle anderen Baumarten erreichten 2000 nur einen Anteil von 0,8 Ofo. Tab. 4 Durchschnittliche Pflanzenzahlen und Baumartenanteile der Verjüngung ab 20 cm. Abb. 79 Baumartenanteile der etablierten Pflanzen.' ab 20 cm ' Probeflächen mit Buche Probeflächen mit Vogelbeere Die dominierende Baumart in den Altbeständen der Hochlagen war bis 1991 die Fichte; andere Arten spielten nur eine untergeordnete Rolle. Die Baumartenzusammensetzung der Verjüngungsschicht wich 1991 allerdings erheblich von derjenigen der Altbestände ab. Der Anteil der Fichte lag damals bei "nur" 73 0/0. Wichtigster Begleiter der Fichte war die Vogelbeere, die knapp ein Fünftel der Verjüngungspflanzen stellte. Der Rotbuche (3 Ofo) und den,sonstigen Baumarten (2 0/0) kamen bereits damals weder in der Baum- noch in der Verjüngungsschicht eine größere Bedeutung zu. Im Laufe der folgenden Verjüngungsinventuren blieben die Baumartenanteile in etwa konstant, erwähnenswert ist hier nur der Anstieg der Rotbuche auf 5 Ofo.....,.. ". Ncht mtvobe und Bu besetzte In~nturpunkte 1<XX> o 1 <XX> Karte 4 Verteilung von Vogelbeere und Buche auf den Probeflächen. 116 ;nx) rv1eter... und der relative Rückgang der Noch deutlicher wird dieser Trend bei den 10 bis 20 cm großen Pflanzen, hier stieg der Anteil der Fichte zwischen 1996 und 2000 kontinuierlich 19% Fichte. Vogelbeere Sonstige Baumarten ' ' " ' ,.,....
 und Douglasie (Pseudotsugo menziesil) beobachtet werden (SCHERZlN GER, pers. Mitteilung). Abb.")



116 n=8) und Weide (So/ix spec., n=l1) zusammengefasst. Ihr Anteil betrug ,7 %, es handelte sich jeweils nur um wenige Exemplare. Abseits der Probeflächen konnte auch das Vorkommen von Waldkiefer (Pinus sy/vestris) und Douglasie (Pseudotsugo menziesil) beobachtet werden (SCHERZlN GER, pers. Mitteilung). Abb.20 Die Vogelbeere spielt bei den Verjüngungsprozessen eine wichtige Rolle. Karte 4 zeigt die räumliche Verteilung von Vogelbeere und Rotbuche in den Hochlagen. Während die Vogelbeere mit Ausnahme des Rachel-Gebietes mehr oder weniger gleichmäßig über die Fläche verteilt ist, tritt Rotbuchenverjüngung nur in den tieferen Lagen im Verzahnungsbereich mit dem Bergmischwald auf. Interessanterweise [----_. kommen Vogelbeere und Rotbuche nie gemeinsam auf einer Probefläche vor... _.. _---_... _ Abb.21 Baumartenanteile im Jahr 2000 in Abhängigkeit von der Höhenklasse.... _._.... _-_... _.... _.... von 84 auf 92 0 /0. Die verbleibenden 8 % verteilen sich zu etwa gleichen Teilen auf Rotbuche und Vogelbeere. Mit zunehmender Pflanzengröße steigt der Anteil der Mischbaumarten und erreicht mit 29 % in der Höhenklasse von ein bis zwei Meter sein Maximum. 19,7 % davon sind Vogelbeeren, 7,3 % Buchen, die sonstigen Baumarten haben einen Anteil von 1,7 %. Unter der Sammelposition sonstige Baumarten wurden, neben der Buche, Latsche (Pinus mugo, n=3). Bergahorn (n=2), Birke (Betu/o pendu/o, [ größer 200 E 100 bis 199 ~ ~ '" ca :i:: c:: OIJ.s= :0 ::I: 20 bis bis 20 0% 20% Fichte 40% 60% 80% 100% Baumartenanteil. Vogelbeere Sonstige Bau marten 117
 I 140")

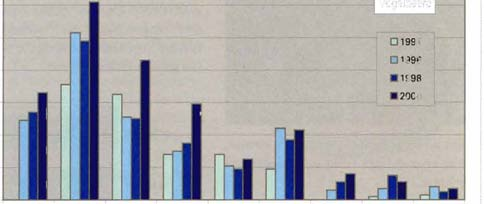

117 ~ I I 'Cii' c ~ ~ I I i I 200! I I L o 10 bis >300 Höhenklasse (ern) I 140 r ~ ~ J I I I I 20 I (cm; >300 i I 0 10 bis BO HÖhe:~;a:~~ L ~~ I I I I ~ 40..c u :0 11\ ~ 30 ::3 Cl s::: :::3 ~ I 1 I I 10 I Abb. 22, 23 u. 24 Entwicklung der Höhenstruktur bei den häufigsten Baumarten. L o 10bis Höhenklasse (ern) > '. I 118
118 5.3 Entwicklung der Höhenstruktur Um die Entwicklung der Höhenstruktur der Verjüngung darzustellen, wurden insgesamt neun Höhenklassen ausgeschieden. Die Auswertung erfolgte anschließend getrennt nach den drei bedeutendsten Baumarten Fichte, Vogelbeere und Rotbuche. Betrachtet man die Entwicklung bei der Fichte, so wird deutlich, dass die Pflanzenzahlen bis 1998 in allen Höhenklassen nur geringfügig anwuchsen. Erst die Aufnahmen 2000 brachten eine erhebliche Steigerung der Pflanzenzahl. Diese Entwicklung beschränkt sich jedoch zum überwiegenden Teil auf,die Höhenklasse kleiner 40 cm. Die Anzahl der Pflanzen zwischen 10 und 20 cm wuchs von 560 auf 1.510, die der Pflanzen zwischen 20 und 40 cm von 233 auf 801 Pflanzen je Hektar. Dies ist darauf zurückzuführen, dass in den vergangenen zwei Jahren viele Bäumchen die Höhenklasse cm durchwachsen haben und deshalb in der Höhenklasse cm erfasst wurden. In den Klassen zwischen 40 und 80 cm wuchs die Anzahl der Fichten nur leicht, über 80 cm veränderten sich die Zahlen fast nicht und waren teilweise sogar leicht rückläufig. Bei der Vogelbeere zeigt sich ein ganz anderes Bild. Der Anstieg von 1998 auf 2000 ist hier nicht so stark ausgebildet und beträgt in keiner Klasse mehr als 70 0/0. Auffällig ist, dass die Pflanzenzahlen im Gegensatz zur Fichte in (fast) allen Klassen ansteigen. In der Klasse cm befinden sich weniger Pflanzen als in der Klasse cm, was auf die großen Jahrestriebe, die ein rasches Durchwachsen der kleinsten Klasse zur Folge haben, zurückgeführt werden kann. Interessant ist auch die Entwicklung seit 1991: Während 1991 nur 24 Pflanzen je Hektar größer als 100 cm waren, erhöhte sich ihre Anzahl bis 2000 auf 77. Diese positive Entwicklung wird in erster Linie auf einen verminderten Verbiss durch Huftiere zurückgeführt. Bei der Rotbuche zeigt sich auf Grund der relativ wenigen erfassten Individuen (n < 100 Pflanzenje Hektar), ein sehr schwierig zu interpretierendes und sehr inhomogenes Bild. Es ist auffällig, dass sich auch bei der Rotbuche die stärksten Veränderungen bei den Pflanzen kleiner 40 cm abspielen. So stieg die Anzahl der Pflanzen je Hektar zwischen 10 und 20 cm von 14 auf 61 und die der Pflanzen zwischen 20 und 40 cm von 3 auf 17 an. Die Anzahl der Bäume in der Klasse zwischen 3 und 5 m Höhe nahm im Gegensatz dazu in den vergangenen Jahren kontinuierlich ab. Grund hierfür ist vermutlich das Aufsteigen dieser Rotbuchen in die Schicht über 5 m. 119




119 5.4 Entwicklung der "Schäden" an der Verjüngung 45 r-~ , Fast 60 % des Gesamtverbisses ist Sommerverbiss, entsteht also, nachdem der neue Trieb bereits gebildet wurde. 'B 14,6 14,4 ],1 c 10. < 4,4 3, Vogelbeere Abb,25 Entwicklung des Leittriebverbisses der etablierten Pflanzen ab 20 cm, Karte 5 Verteilung der Probeflöchen mit Leittriebverbiss,....,, ' '.. - Fichte - Gesamt Schalenwild Wie schon in den vergangenen Jahren wurde nur der Leittriebverbiss durch Schalenwild erfasst. Die im Gebiet vorkommenden Arten sind Reh (Capreolus capreolus), Rothirsch (Cervus elaphus) und Wildschwein (Sus scrofa), wobei letzteres keinen Verbiss verursacht. Es ist nicht möglich, den Verbiss von Rothirsch und Reh voneinander zu differenzieren. Verbiss durch den Feldhasen (Lepus europaeus), der auf Grund einer schrägen, glatten Schnittfläche eindeutig zugeordnet werden kann, wurde nicht aufgenommen. Die Verbissbelastung bei der Vogelbeere hat sich seit 1998 in etwa stabilisiert bzw. ist sogar leicht zurückgegangen. Sie weist aber nach wie vor mit 14,4 % verbissener Individuen die höchste Verbissbelastung auf. Gegenüber der ersten Hälfte der neunziger Jahre, als der Verbiss noch bei 44,7 % lag, hat sich die Situation jedoch deutlich entspannt. Dies hatte auch zur Folge, dass sich die Höhenstruktur (s. dort) der Vogelbeerverjüngung deutlich verbessert hat. Die Vogelbeeren kleiner 80 cm weisen einen hohen Anteil verbissener Pflanzen auf. Das Maximum der Verbissbelastung liegt in der Klasse der cm hohen Pflanzen, dort sind 35 % der Individuen verbissen. Vogelbeeren mit einer Höhe von über 2 m wurden nicht verbissen. Der Anteil von Rotbuchen mit Verbiss liegt aktuell bei 5,4 %, allerdings wurden bei dieser Baumart nur relativ wenige Individuen erfasst, so dass der Rückgang seit 1998 vorsichtig zu interpretieren ist. Damals lag der Leittriebverbiss bei 16,2 % Der Verbiss der Fichte liegt bei 0,7 0/0. Er bewegt sich weiterhin auf sehr niedrigem Niveau und spielt bei der Entwicklung der Pflanzen eine sehr untergeordnete Rolle. Der Gesamtverbiss erreichte 2000 mit 3,5 % den niedrigsten Wert seit Beginn der Untersuchungen. Die Verbissschwerpunkte liegen im Bereich zwischen Hochfilzberg und dem Großen Schwarzbach..'.... ' Prcbeflächen ome Verbiss. Prcbeflächen mit Leittrieb\erbiss 1000 o Meter. 120 ' '.'.'..,.... '..',..... '.... '.'......,,,....,... ',,., '. '.'.'....,.. '.... '.'

 und der Große Schwarze Rüsselkäfer (Otiorrhynchus niger) sein.")

120 Der Große Schwarze Rüsselkäfer schädigt die Fichtenverjüngung doppelt: er frisst ebenfalls oberirdisch an der Rinde von jungen Fichten, zusätzlich ernähren sich seine Larven, die sich im Boden entwickeln, von der Rinde der Wurzel, die sie abschaben. Beide Arten sind schwierig festzustellen, da sich die Käfer bei Erschütterungen sofort zu Boden fallen lassen. Abb. 26 Verbiss durch Schalenwild, zu erkennen an der ausgefransten Biss-Stelle Rüssel käfer Von Bedeutung für die Entwicklung der Fichtenverjüngung können der Große Braune Rüsselkäfer (Hylobius abietis) und der Große Schwarze Rüsselkäfer (Otiorrhynchus niger) sein. Es handelt sich bei diesen Arten um mittelgroße (1-1,5 cm lange) Käfer mit einem ausgeprägten "Rüssel", an dessen Ende sich die Mundwerkzeuge befinden. Die Larven des Großen Braunen Rüsselkäfers entwickeln sich in den Stockwurzeln frisch gefällter Fichten, eventuell auch in frisch abgestorbenen Fichten. Der Käfer selbst frisst an der Rinde von jungen Fichtenpflanzen vor allem im Bereich oberhalb des Wurzelknotens, aber auch an deren Ästchen. Bei starkem Befall kann dieser plätzeweise, trichterförmige Fraß zur Schwächung und zum Absterben der Pflanzen führen. Obwohl die beiden Arten doch recht verbreitet sind konnten Rüsselkäfer auf jeder siebten Probefläche beobachtet werden - sind die durch sie verursachten Schäden sehr gering. So wurden 1998 nur an 1,5 % der Fichten Fraßspuren festgestellt. Im Jahr 2000 ging der Befall noch weiter zurück, nur sechs Pflanzen wiesen Rindenschäden auf (0,06 % der Gesamtpflanzenzahl) Buchdrucker Normalerweise leben Buchdrucker (Ips typographus) unter der Rinde von Fichten, die mindestens einen BHD von 20 cm haben. Wenn während Massenvermehrungen große Mengen von Käfern auf der Suche nach geeigneten Fichten sind und gleichzeitig das Angebot an solchen Fichten knapp wird, bohren sich die Käfer auch in viel kleinere Bäume ein. Hier können sie zwar nicht erfolgreich brüten, aber doch ihre Nährstoffreserven auffüllen. Abb.27 Buchdruckerbefall an Jungfichten. Abb. 28 u. 29 Großer Brauner Rüsselkäfer (links); Großer Schwarzer Rüsselkäfer (rechts). aus AMANN (7990): Kerfe des Waldes. (Mit freundlicher Genehmigung durch Frau Claudia Summerer, Ottobeuern) 121

 sich nicht wesentlich")
 ist ein Blattparasit, der in Gebieten mit hoher Schneelage vorkommt.")

121 1996 und 1997 konnte dieses Verhalten auch im Nationalpark - vor allem im Lusenbereich - beobachtet werden. Damals bohrten sich die Käfer sogar in nur daumenstarke Verjüngungspflanzen ein. Bei einer Erhebung an ausgewählten Probeflächen in den ausgedehnten Totholzkomplexen stellten die Aufnahmetrupps 1997 fest, dass ca. 10 % der Verjüngung dur~h den Buchdrucker zum Absterben gebracht wurden (NUESSLEIN, 1997). Dieses Phänomen wurde in den folgenden Jahren nicht mehr beobachtet, obwohl die Rahmenbedingungen (großflächiges Absterben von Altbeständen, hoher Befallsdruck beim Buchdrucker) sich nicht wesentlich geändert hatten Schwarzer Schneeschimmel Abb. 31 Durch Schnee gebrochene Fichte. Abb.30 Schneeschimmelbefall an Fichten. Der Schwarze Schneeschimmel (Herpotrichia juniperi) ist ein Blattparasit, der in Gebieten mit hoher Schneelage vorkommt. Das dunkelbraune bis schwarze Mycel dieses Pilzes entwickelt sich unter der Schneedecke und überwuchert Zweige und Nadeln, was zum Absterben von Verjüngungspflanzen führen kann. Obwohl in der Vergangenheit immer wieder Schäden durch den Schwarzen Schneeschimmel beobachtet wurden, konnten im Verlauf der Inventuren auf den Probeflächen bisher keine Schäden durch den Pilz nachgewiesen werden Schneebruch In den beiden vergangenen Wintern 1998/99 und 1999/2000 kam es zu umfangreichem Schneebruch im gesamten Nationalparkgebiet. 1998/99 waren vor allem die höheren Lagen, 1999/2000 vor allem die unteren und mittleren Lagen betroffen. Auch in der Verjüngung der Hochlagenstufe konnten Schäden auf Grund dieser Ereignisse nachgewiesen werden. Diese hielten sich jedoch in Grenzen. Nur 2,5 % der Pflanzen größer 1 m hatten Wipfel- und Schaftbrüche erlitten. Vogelbeere und Fichte waren gleichermaßen betroffen. 122



122 5.5 Verteil ungsrn uster der Verjüngung innerhalb der Bestände Klumpung der Verjüngung Wie bereits bei der Entwicklung der Verjüngungsdichten erwähnt, sind die Verjüngungspflanzen nicht gleichmäßig auf die einzelnen Probeflächen verteilt. Ein Großteil der Flächen - im Jahr 2000 waren es 46 % - weist keine Verjüngung über 20 cm auf. Im Vergleich dazu ist der Anteil von Kreisen mit 1 bis 999 Pflanzen je Hektar mit 22 0/0 relativ gering. Nur 32 % der Probeflächen haben eine Verjüngungsdichte von über Pflanzen je Hektar. Aus Abbildung 32 wird aber auch deutlich, dass sich die Situation seit 1991 erheblich verändert hat. Insbesondere der Anteil unverjüngter Probeflächen ging von 70 auf 46 % zurück. Dementsprechend waren im Jahr 2000 fast doppelt so viele Flächen verjüngt wie im Jahr l c QJ.s::. tj :(11 ii= QJ.s::J e ~ Qj ~ 'Qj... t:: <C Pflanzen größer 20 cm I o >5000 Verjüngungsdichte (Pfl./ha) 80. Betrachtet man die Häufigkeitsverteilung aller Pflanzen größer 10 cm, so zeigt sich ein sehr ähnliches Bild. Allerdings ist hier der Anteil der unverjüngten Probeflächen mit 33 % wesentlich geringer, die anderen Klassen sind entsprechend stärker besetzt. Erwähnenswert ist hier vor allem der Anstieg der Anzahl der Probeflächen mit einer Verjüngungsdichte von über Pflanzen je Hektarvon 10auf190f0. l 60 c QJ.s::. 50 tj :(11 ii= QJ.s::J 0 ~ QJ 70 1 Pflanzen größer 10 cm I 40 ~ 30 'Qj... t:: <C Diese Häufigkeitsverteilung der Verjüngungsdichten auf den Probeflächen deutet darauf hin, dass die Verjüngung in den Hochlagen stark geclustert, also geklumpt auftritt. Bezieht man die Entwicklung seit 1991 mit in die Betrachtung ein, deutet sich ein Trend hin zu einer weniger stark geclusterten, etwas homogeneren Verteilung der Verjüngung an. Der Anteil der unverjüngten Flächen darf allerdings nicht dahingehend interpretiert werden, dass 46 % bzw. 33 Ofo der Hochlagen ohne Verjüngung sind. Die Wahrscheinlichkeit, ob eine Probefläche Verjüngung aufweist oder nicht, 10 0 o >5000 Verjüngungsdichte (Pfl./ha) hängt ganz entscheidend von deren Größe ab, da sich mit zunehmender Probeflächegröße automatisch der Anteil der unverjüngten Probeflächen verringert. Es war deshalb zu klären, auf welcher räumlichen Skala die Klumpung auftritt und ob eventuell größere Bereiche der Hochlagen ohne Verjüngung sind. NUESSLEIN (1998) hat diese Frage im Zuge der Inventur von 1998 geprüft, indem er Abb.32 Entwicklung der Häufigkeitsverteilung der Probeflächen in Abhängigkeit von der Verjüngungsdichte. 123
123 Abb. 33 Flächenanteile der aspektbestimmenden Kleinstrukturen im Jahr Zwergsträ ucher 8,2% Bewachsener Felsen 10,5% die Verjüngung zusätzlich auf 500 m großen Probeflächen untersuchen ließ. Diese Fläche entspricht in etwa dem Standraum eines Altbaumes. Darüber hinaus wurde das Inventurrasterverdichtet. Ergebnis war, dass nur auf 6 0J0 der insgesamt Probeflächen keine Verjüngung größer 20 cm gefunden werden konnte. Die Klasse 1 bis 999 Pflanzen je Hektar war mit 57 % der Probeflächen besetzt und auf 36 0J0 der Kreise wuchsen mehr als Pflanzen je Hektar. Im Vergleich dazu wiesen 58 0J0 der 25 m' großen Probeflächen keine Verjüngung auf, 17 0J0 waren in der Klasse und 25 0J0 in der Klasse mit mehr als Pflanzen je Hektar. Diese Ergebnisse lassen darauf schließen, dass die Klumpung der Verjüngung in den Hochlagen auf kleiner Skala erfolgt und großflächige Bereiche ohne Verjüngung eher die Ausnahme sind. Im Folgenden werden die Gründe für die Klumpung derverjüngung näher untersucht Kleinstrukturen der Waldbodenoberfläche 4,9% 3,4% 2,0% Ziel der Aufnahme der aspektbestimmenden Kleinstrukturen war es, detaillierte Informationen über die Bedeutung dieser Strukturen für die Verteilung der Verjüngung zu gewinnen. Darüber hinaus sollte der Einfluss des Absterbens der Fichtenaltbestände auf die Entwicklung der Kleinstrukturen untersucht werden. Die Ansprache der Flächenanteile der Kleinstrukturen erfolgte 1996 und Die Verteilung der einzelnen Fichtenpflanzen auf die verschiedenen Kleinstrukturen wurde nur 1996 erhoben. In den Hochlagen dominiert die Gras-Fazies, fast die Hälfte der untersuchten Fläche ist von "Gräsern" bedeckt. Größere Flächenanteile nehmen auch noch die bewachsenen Felsen mit 10,5 0J0 sowie die Zwergstrauch-Fazies, die Farn-Fazies und der Humus mit jeweils ca. 8 0J0 ein. Die verbleibenden Flächen verteilen sich auf die Moos-Fazies (4,9 0J0), den Baumkontakt (4,9 0J0) und das Moderholz (3,4 0J0). Die Kleinstrukturen Rinde/Reisig, Kraut und Himbeere, Wurzelteller und Bärlapp nehmen mit einem Anteil von jeweils unter 2 0J0 nur eine geringe Fläche ein. Auf die Veränderungen der Flächenanteile der einzelnen Kleinstrukturen wurde durch einen Vergleich der Aufnahmen von 1996 und 2000 geschlossen. Von den zwölf ausgeschiedenen Strukturelementen konnten, ausgelöst durch das Absterben der Altbestände, sieben ihren Flächenanteil vergrößern, fünf verloren Flächenanteile. Auf Grund des verstärkten Lichteinfalls breiteten sich vor allem die Gräser weiter aus. Aber auch Farne und Kräuter profitierten von den Veränderungen in der Baumschicht. Der Anteil der Kraut-Fazies wuchs um 69 0J0. Das Herabfallen von Ästen und Rinde abgestorbener Bäume hat auch zu dem sprunghaften Anstieg von 152 0J0 bei der Kreinstruktur Rinde/Reisig geführt. Durch das Umstürzen toter Bäume stiegen auch die Flächenanteile von Moderholz und Baumkontakt, jedoch nur in 124



124 geringem Ausmaß. Dass diese Flächenveränderungen so gering al,jsfielen, ist vor allem dadurch begründet, dass zum Aufnahmezeitpunkt nur ein Teil der abgestorbenen Altbäume umgebrochen war und viele umgestürzte Stämme nur teilweise Bodenkontakt hatten. Diese Stämme wurden (noch) nicht der Kleinstruktur Moderholz zugeordnet. Die geringe Zunahme bei "bewachsene Felsen" wird auf die schwierige Ansprache dieses Elementes auf Blockfeldern zurückgeführt. Grund könnte aber auch der Rückgang von Auflagehumuspolstern und dadurch verursacht eine genauere Ansprache der Kleinstruktur sein. Wegen der Ausbreitung von konkurrenzkräftigen Gräsern und Farnen wurden die Zwergstrauch- und Moos Fazies sowie der Humus verdrängt bzw. überwachsen. Das Gleiche gilt auch für die Bärlapp Fazies und den Wurzelteller, hier wurde die Wurzeltellermulde meist von Gräsern überwuchert. Die letztgenannten Kleinstrukturen verloren über 35 % ihrer Fläche. Veränderung Veränderung in m 2 in 0/0 Moderholz 16,56 3,39 Baumkontakt 63,32 9,71 Bewachsener Felsen 28,16 1,86 Rinde/Reisig 177,13 Wurzelteller -20,79 Humus -179,77-13,00 Zwergsträucher -185,12-13,38 Moos -177,36-19,77 Bärlapp -26,29-37,72 Kraut 64,57 68,68 Gras 209,68 3,07 Farn 127,97 12,20 Abbildung 34 zeigt, wie stark sich die Verjüngungsdichten der einzelnen Kleinstrukturen unterscheiden. Berücksichtigt wurden alle aufgenommenen Pflanzen. Die höchsten Verjüngungsdichten finden sich mit Pflanzen je Hektar auf Moderholz und mit Pflanzen je Hektar _._._ _._ _.-_ _--_._--_._ , ,... C'O..s:::: ~ ~ 6000 Q.. ~ s:::: tj ~ Cl c::: ;:, Cl c::: :;:,.~ ~ im Kontaktbereich zu stehenden und liegenden Stämmen. Auch die Kleinstruktur Wurzelteller weist mit Pflanzen je Hektar relativ hohe Werte auf. Oberhalb der mittleren Individuenzahl der Inventur 1996 (1.698 Pflanzen je Hektar) liegen auch die Verjüngungsdichten auf bewachsenem Fels, auf Humus und auf der Zwergstrauch-, der Moos- und der Bärlapp-Fazies. Eine unterdurchschnittliche Verjüngungsdichte finden wir bei Reisig/Rinde, Kraut und Himbeere und insbesondere bei der Farn- und Gras-Fazies. Auf Gras beträgt die durchschnittliche Pflanzenzahl je ha nur361 bei Farn 575. Um die Bedeutung der einzelnen Kleinstrukturen für die Verjüngung in den Hochlagen abschätzen zu können, wurde der Anteil der Verjüngungspflanzen bestimmt, der auf dem jeweiligen Strukturelement wächst. Als Maß für die relative Bedeutung der jeweiligen Kleinstruktur wurde der Quotient aus dem Anteil der Fichtenindividuen auf der entsprechenden Kleinstruktur und dem Flächenanteil der Kleinstruktur bestimmt. Bei der Analyse wurden nur die Pflanzen kleiner 60 cm Abb.34 Durchschnittliche Verjüngungsdichte der Pflanzen ab 10 cm in Abhängigkeit von der Kleinstruktur im Jahr Tab. 5 Flächenveränderungen der Kleinstrukturen seit
.")


125 Abb.35 Anteil der Fichte und der Vogelbeere< 60 cm auf der jeweiligen Kleinstruktur (absolute Bedeutung) und Quotient aus diesem Anteil und dem Flächenanteil der entsprechenden Kleinstruktur (relative Bedeutung). l25 berücksichtigt, da mit zunehmender Größe und damit auch höherem Alter der Bäume zum einen die Bedeutung der Kleinstruktur für die Entwicklung der Pflanzen immer geringer wird, zum anderen die Wahrscheinlichkeit steigt, dass sich die Kleinstruktur veränderte, seit sich die Pflanze etabliert hat. 35 -r t Fichte 35 -~ ~ Vogelbeere o ~ relative Bedeutung Cl r::::.a 6 :::l QJ "0 5 ~ 4 ~ CI! "ii 3 CI: 2 o 7 6 ~ :::l... 5 ~ QJ ce - 4 QJ > +' - 3 ~ CI: 2-1 o Vergleicht man die Bedeutung der Kleinstrukturen für die Verjüngung getrennt nach Fichte und Vogelbeere, so zeigen sich große Unterschiede zwischen den beiden Baumarten: Die meisten Vogelbeeren finden sich in der Gras- Fazies, fast ein Drittel der Individuen dieser Baumart wächst hier. Im Gegensatz dazu hat diese Kleinstruktur als Wuchsort für die Fichtenverjüngung keinen großen Stellenwert: Nur 5,7 % der Fichten wachsen hier. Von entscheidender Bedeutung für die Verjüngung der Fichte sind Moderholz und Baumkontakt. 44 % der Fichten wachsen auf diesen beiden Kleinstrukturen. Für die Verjüngung der Vogelbeere spielt Moderholz mit unter 3 % der gesamten Verjüngungspflanzen nur eine untergeordnete Rolle. Der Schutz durch stehende und liegende Bäume bzw. Baumleichen ist auch für die Entwicklung der Vogelbeere von Bedeutung, wenn auch nicht in gleichem Maße wie bei der Fichte. 11 % der Vogelbeeren und knapp 22 % der Fichten wachsen auf der Kleinstruktur Baumkontakt. Auf Humus und der Zwergstrauch-Fazies entwickeln sich Fichte und Vogelbeere in etwa gleich gut, auch der Anteil der dort wachsenden Pflanzen ist bei beiden Elementen mit ca. 10 % in etwa gleich. Während sich auf der Moos-Fazies mehr Vogelbeeren entwickeln können, sind die Wachstumsbedingungen für die Fichte auf bewachsenen Felsen günstiger. Auffällig ist auch der hohe Anteil der Vogelbeerenverjüngung in Rinde/ Reisig. Die Vogelbeere kommt hier neunmal öfter vor, als es bei einer Gleichverteilung der Vogelbeeren auf die Kleinstrukturen zu erwarten wäre. Die Fichte ist hier nur zweimal häufiger vertreten. Die verbleibenden Kleinstrukturen sind auf Grund ihrer geringen Flächenausdehnung von untergeordneter Bedeutung. Extrem verjüngungshemmend ist die Farn-Fazies. Obwohl sie 8 % der Flä che bedeckt, konnte hier nur knapp 1 % der aufgenommenen Pflanzen gefunden werden. Weder Vogelbeere noch Fichte können hier Fuß fassen. 126
 Dichte derverjüngung größer 20 cm. Sie dient als Maß für den aktuellen Zustand der etablierten Verjüngung.")

.")
 sowie organische und anorganische Nassböden.")
126 5.6 Verteil ungsin uster der Verjüngung iin Gelände Im Folgenden wird der Einfluss verschiedener Standortseigenschaften auf die Verjüngungsdichte untersucht. Zur Charakterisierung der Verjüngungssituation wurden drei Kennzahlen verwendet: a) Dichte derverjüngung größer 20 cm. Sie dient als Maß für den aktuellen Zustand der etablierten Verjüngung. b) Die Differenz der Pflanzenzahl zwischen 1991 und 2000 bei den Pflanzen größer 20 cm. Sie beschreibt die Entwicklung der etablierten Verjüngung. c) Dichte derverjüngung zwischen 10 und 20 cm Größe. Sie gibt Hinweise auf die mögliche Entwicklung einer sich künftig etablierenden Verjüngung Bodenform Die Attributierung der Bodenformen erfolgte durch Verschneidung der Bodenkarte und der Inventurpunkte aus dem Referenz-GIS der Nationalparkverwaltung. Grundlage dafür war die Bodenkartierung durch ELLING et al. (1987). Anschließend wurden die damals ausgeschiedenen Bodenformen zu vier ökologischen Gruppen zusammengefasst, nämlich Block-Humus-Böden, Lehmböden, mittelgründiger Lehm über verfestigtem Schutt (das Wurzelwachstum behindernde Schicht in cm Tiefe) sowie organische und anorganische Nassböden. Für alle drei Kennzahlen gilt, dass die Verjüngungsdichte auf mittelgründigem Lehm über verfestigtem Schutt am geringsten ist. Die Anzahl der etablierten Pflanzen je Hektar beträgt hier nur Auch der Zuwachs seit 1991 ist hier mit 298 Pflanzen je Hektar nur gering. Dass sich in der Schicht zwischen 10 und 20 cm Individuen je Hektar befinden, lässt in Zukunft eine höhere Anzahl etablierter Pflanzen erwarten. Mit im Cl! 2500 ~ c;:: e:, 2000 CI.)....J:: u ~ Cl c: :J Cl c: ::J. ~ CI.) > 'Ci 3000 ~ ~ 2500 ~.J:: U ~ 2000 Cl c: :l Cl c: ::l 1500.~ ~ ~ 1000 "'C Cl c: :l ~ 500 "'C ::ii ~ 'Cii' 2500 ~ """7' ~ 2000 QJ +" ~ t.i 1500 ~ VI Cl c: ::: Cl c: ::::3 ';::- 500 ~ c: CI.) "'C I :0 """ U ce I o VI iij ~ I VI :J :J: E c: :J CI.) :J:"'C I :0 """ U ce 0 iij I VI :J E c: :J CI.) :J:"'C I :0 t)ce 0 iij ] E CI.) c: CI.) CI.)....0 "'C ~ ::::l.~!; :0 E t:i.j:: ~~~.0 'iil z CI.) > ~ 60 c: CI.) ~ 50 c;:: CI.) ~ ~ Cl 30 c: ::J. ~ CI.) 20 > 10 OE c: oe:( Da die abhangige n Vanablen nicht und die unabhanglgen Vanab len zum überwiegenden Teil nicht normalverteilt sind, wurde die Stärke des Zusammenhangs mit Rangkorrelationskoeffizienten gemessen. Eingesetzt wurden die Rangkorrelationskoeffizienten "Kendalls Tau-b" und "Spearmans Rho ". Zum Ve rg leich unabhängiger Stichproben wurde der "Mann-Whitney-U-Test" verwendet. Die statistischen Berechnungen wurden mit dem Softwarepakt SPSS und die räumlichen Analysen mit der GIS-Softwa re Arc View durchgeführt. Abb.36 Du rchschn i ttl iche Verjüngungsdichte (Balken) und Anteil verjüngter Probeflächen (Linie) der etablierten Pflanzen ab 20 cm im Jahr 2000 in Abhängigkeit von der Boden form. Abb.37 Entwicklung der durchschnittlichen Verjüngungsdichte der etablierten Pflanzen ab 20 cm zwischen 1991 und 2000 in Abhängigkeit von der Bodenform. Abb,38 Du rchsch n i ttl iche Verjüngungsdichte der Pflanzen zwischen 10 und 20 cm im Jahr 2000 in Abhängigkeit von der Boden form. 127



127 Abb. 39 Du rchsch n i ttl iche Verjüngungsdichte (Balken) und Anteil verjüngter Probeflächen (Linie) der etablierten Pflanzen ab 20 cm im Jahr 2000 in Abhängigkeit von der Meereshöhe. Abb.40 Entwicklung der durchschnittlichen Verjüngungsdichte der etablierten Pflanzen ab 20 cm zwischen 7997 und 2000 in Abhängigkeit von der Meereshöhe. Abb. 47 Du rchschn ittl iche Verjüngungsdichte der Pflanzen zwischen 70 und 20 cm im Jahr 2000 in Abhängigkeit von der Meereshöhe. Durchschnitt Pflanzen je Hektar ist die Anzahl etablierter Pflanzen auf Lehm zwar höher wie auf Lehm über verfestigtem Schutt, der Unterschied ist jedoch nicht signifikant. Allerdings verlief die Entwicklung auf Lehmböden seit 1991 mit einem Zuwachs von 741 Pflanzen je Hektar wesentlich günstiger. Die Anzahl der Individuen zwischen 10 und 20 cm ist hier mit Pflanzen je Hektar fast doppelt so hoch wie auf Lehm über verfestigtem Schutt und sogar höher als auf Nassböden. Die höchsten Verjüngungsdichten finden sich auf den Nassböden und den Block-Humus-Böden. Alle drei Kennzahlen zeigen hier vergleichsweise hohe Werte. Die durchschnittliche Verjüngungsdichte liegt bei den etablierten Pflanzen auf Nassböden bei Pflanzen je Hektar und bei Block-Humus-Böden bei Pflanzen je Hektar. Auch bei den 10 bis 20 cm großen Individuen sind die Unterschiede zwischen diesen Bodengruppen signifikant, allerdings ist hier die Verjüngungsdichte auf den Nassböden mit Pflanzen je Hektar geringer als auf Block- Humus Böden, wo sich im Durchschnitt Pflanzen je Hektar befinden. Keinen signifikanten Unterschied gibt es zwischen diesen Bodengruppen bei der Entwicklung der Verjüngungsdichte der etablierten Pflanzen seit Auch der Anteil von Probeflächen mit Verjüngung folgt dem beschriebenen Muster. Während auf Lehm über verfestigtem Schutt und Lehm nur 49 % bzw. 52 % der Kreise Verjüngung aufweisen, sind es bei den Fels-Blockböden 61 % und bei den Nassböden 74 Ofo Meereshöhe Die Zuo rdnung der Meereshöhe erfolgte mit Hilfe eines digitalen Geländemodells, dessen räumliche Auflösung 50 m (Grid) beträgt l (;j' c.s::: QJ.s::: (J " "" ~ '7: 3000 QJ 50.&l :g 2500 ~ 0-40 ~ 2000 ~ g, Cl C :;:3.0=- : QJ > ~ 'E c 0 0 ct < > ~ 4000 ~ 3500 i g, 2500 ::s.c' t.., 1500 u CI c: 1000 ~ 500! 0 ~, IV.J: ""': ~ 3500 QJ '.J: 2500 c.j :g cn c :::l 1500 cn c ::::l.:- ~ Meereshöhe (m) < > Meereshöhe (m) < > Meereshöhe (m) 128


128 Diagramm 39 zeigt eine deutliche Abhängigkeit der Verjüngungsdichte etablierter Pflanzen von der Meereshöhe. Während die Höhenlagen unter 1200 m mit mehr als Pflanzen je Hektar als gut verjüngt bezeichnet werden können, findet man über 1200 m nicht mehr als etablierte Pflanzen je Hektar. In der Höhenstufe über 1300 m wachsen sogar nur 925 Pflanzen je Hektar. Auch der Anteil von Probeflächen mit etablierter Verjüngung nimmt kontinuierlich mit der Meereshöhe ab. Während unter 1150 m 70 % aller Probeflächen verjüngt sind, sind es über 1300 m nur 36 0/0. Für die Entwicklung der Pflanzen seit 1991 ergibt sich ein ähnliches Bild: eine starke Zunahme der Verjüngungsdichte um über Pflanzen je Hektar in den Lagen unterhalb 1200 m und nur geringe Steigerungen zwischen 311 und 98 Pflanzen je Hektar oberhalb von 1200 m. Deutlich günstiger zeigt sich die Situation, wenn auch die Pflanzenzahlen zwischen 10 und 20 cm berücksichtigt werden: bis in eine Meereshöhe von 1250 m befinden sich über Pflanzen je Hektar in dieser Schicht und selbst darüber sind es noch etwa 800 Pflanzen je Hektar, von denen sich in den nächsten Jahren vermutlich ein Großteil etablieren wird Exposition Die Exposition wurde mit Hilfe eines Kompasses in der direkten Umgebung des Aufnahmekreises bestimmt und in acht Himmelsrichtungen klassifiziert. Für die Auswertung wurden die schattigen, kühlen und feuchten Bereiche, also Nord, Nordost, Nordwest und Ost, sowie die sonnigeren, warmen und trockeneren Lagen Südost, Süd, Südwest und West zu jeweils einer Gruppe zusammengefasst. Die Verjüngungsdichten der Pflanzen größer 20 cm waren in den warmen Lagen höher (1.760 Pfl./ha) als in den kühlen (1.445 Pfl./ha). Auch der Anteil verjüngter Probeflächen ist in den wärmeren Lagen mit 59 % höher, als in den kühleren mit nur 46 % Auch die Entwicklung der Verjüngungsdichte seit 1991 verlief in den wärmebegünstigten Expositionen mit einer Zunahme von 605 Pflanzen je Hektar günstiger als in den kühlen mit einem Zugang von nur 577 Pflanzen je Hektar. Obwohl der Unterschied nur gering ausfällt, ist er doch signifikant. Im Gegensatz dazu waren die Unterschiede zwischen den Gruppen bei den Pflanzen der Schicht von 10 bis 20 cm nicht signifikant. Die Werte liegen hier in den kühlen Lagen bei und in den wärmebegünstigten bei Pflanzen je Hektar Geländeneigung Die Geländeneigung wurde am jeweiligen Inventurpunkt mit dem Neigungsmesser Suunto bestimmt. Die nachträgliche Klassifikation der Messwerte wurde in Anhalt an die Richtlinien des Arbeitskreises Standortskartierung in der Arbeitsgemeinschaft Forsteinrichtung (1980) durchgeführt. Die Neigung in Grad Neigungsstufe 0-4,9 eben bis schwach geneigt,",--- 5 bis 10 mäßig geneigt 10 bis 20 stark geneigt >20 steil - - höchsten Verjüngungsdichten etablierter Fichten findet man mit Pflanzen je Hektar in den steilen Hängen. Im Gegensatz dazu beträgt die Verjüngungsdichte in den stark geneigten Bereichen Pflanzen je Hektar. Die wenigs- Tab. 6 Klassifizierung der Hangneigung. 129

 und in den schwach geneigten (1.035 Pfl./ha) Bereichen. Die Unterschiede zwischen den ausgeschiedenen Gruppen sind nicht signifikant.")
129 Abb.42 Durchschnittliche Verjüngungsdichte (Balken) und Anteil verjüngter Probeflächen (Linie) der etablierten Pflanzen ab 20 cm im Jahr 2000 in Abhängigkeit vom Absterbezeitpunkt. Abb.43 Entwicklung der durchschnittlichen Verjüngungsdichte der etablierten Pflanzen ab 20 cm zwischen 1991 und 2000 in Abhängigkeit vom Absterbezeitpunkt. Abb.44 Durchschnittliche Verjü ngu ngsd ich te der Pflanzen zwischen 10 und 20 cm im Jahr 2000 in Abhängigkeit vom Absterbezeitpunkt. ten Verjüngungspflanzen befinden sich in mäßig (1.105 Pfl./ha) und in den schwach geneigten (1.035 Pfl./ha) Bereichen. Die Unterschiede zwischen den ausgeschiedenen Gruppen sind nicht signifikant. Auch der Anteil der mit etablierten Pflanzen verjüngten Probeflächen zeigt keine deutlichen Unterschiede zwischen den Klassen: Auf den schwach geneigten Hängen sind 51 Ofo, auf den mäßig geneigten Hängen 57 Ofo, auf den stark geneigten Hängen 53 Ofo und auf den steilen Hängen 51 Ofo der Probeflächen verjüngt. Auch zwischen der Veränderung der Verjüngungsdichte seit 1991 und der Geländeneigung konnte kein signifikanter Zusammenhang festgestellt werden. Bei den Pflanzen zwischen 10 und 20 cm findet man ein etwas anderes Muster wie bei den etablierten Individuen. Die höchste Verjüngungsdichte weisen hier die stark geneigten Hänge mit Pflanzen je Hektar, gefolgt von den steilen Hängen mit Pflanzen je Hektar und den mäßig geneigten Hängen mit Pflanzen je Hektar auf. Die geringste Verjüngungsdichte findet sich mit Pflanzen je Hektar auf den schwach geneigten Hängen. Die Unterschiede zwischen den schwach und mäßig geneigten Hängen sind ebenso signifikant, wie die Unterschiede zwischen den schwach und stark geneigten Hängen Zeitpunkt des Absterbens der Altbestände Das Jahr, in dem die Altbestände über der jeweiligen Probefläche abgestorben sind, wurde durch Verschneidung der Inventurpunkte und der Ergebnisse der jährlichen Auswertung der Totholzflächenentwicklung bestimmt. Es konnte bei allen Kennzahlen ein Zusammenhang zwischen Absterbezeitraum und Verjüngungsdichte nachgewiesen werden. Darauf deuten schon die hohen Verjüngungszahlen auf den ,- 80 (;j' 3000.s:: ~ 2500 v i 'ö [ 1500 Cl :~ 1000 ~ 500 cu -0 Cl r::: 2 cu -0 r::: :ca ~ o 3500 Ci > Uj c.o " CX) m m m m m m m m Absterbejahr 70 ~ r:: 60 ~ i;: 50 ~ o ct 40 E Cl 30 :5. ~ 20 ~ Cü 10 ~ o r , ro 3000 ~ ~ 2500 E 2000 tj '-ö ~ 1500 ::I Cl : f 500 o o '-<:t--<:t--uj--c.o--,,--cx)- -o--' r::: m m m m m m ~.D ~ Ci > Ci > Uj m Absterbejahr c.o " m m Absterbejahr.s:: tj o r::: -0 r::: ~.D ~.s:: tj o r::: 130


130 Probeflächen hin, über denen die Bestände im Jahr 1996 und vor 1994 abgestorben sind. Unter den 1996 abgestorbenen Beständen ist die Verjüngungsdichte mit Pflanzen je Hektar doppelt so hoch wie auf den Probeflächen über denen die Bestände in den Jahren 1995 und 1997 abgestorben sind. Auch wenn man die Entwicklung seit 1991 betrachtet, fällt besonders die hohe Zunahme der Verjüngungsdichte auf den Probeflächen auf, über denen die Bestände im Jahr 1996 abgestorben sind. Auf diesen Flächen nahm die Verjüngungsdichte um Pflanzen je Hektar zu. In den 1995 abgestorbenen Bereichen stieg die Verjüngungsdichte nur um 502 Pflanzen je Hektar, in den 1997 abgestorbenen Bereichen nahm sie um 164 Pflanzen je Hektar ab. Noch deutlicher wird dieses Muster bei den Pflanzen zwischen 10 und 20 cm. Im Jahr 1996 liegt hier die durchschnittliche Verjüngungsdichte bei Pflanzen je Hektar, das sind 144 bzw. 205 Ofo mehr als 1995 und 1997.Nur die Inventurpunkte, über denen die Bestände vor 1994 abgestorben sind, weisen bei den etablierten Pflanzen mit Pflanzen je Hektar noch höhere Verjüngungsdichten auf. Auch verlief die Entwicklung seit dem Jahr 1991 auf diesen Probeflächen besonders günstig. Bei den Individuen zwischen 10 und 20 cm liegt die Verjüngungsdichte mit Pflanzen je Hektar nur etwas niedriger als in den Probeflächen, über denen die Bestände im Jahr 1996 abgestorben sind. Ausgelöst wurden die hohen Verjüngungsdichten auf den im Jahr 1996 bzw. in den vor 1994 abgestorbenen Probeflächen durch die starke Fruktifikation der Fichte 1988, 1992 und 1995, die zu einer großen Anzahl von Keimlingen im Folgejahr führte. Als dann die Bestände im Jahr nach der Fruktifikation abstarben, gelangten vermehrt Licht und Wärme auf den Waldboden, was die Entwicklung der Keimlinge ermöglichte. Die Samenjahre haben somit nur dann einen deutlichen Effekt, wenn ihnen der Zusammenbruch der Altbestände folgt. Der Anteil der mit etablierten Individuen verjüngten Probeflächen gibt diese Entwicklung nicht so deutlich wieder. Er liegt 1995, 1996 und 1997 mit einem Wert zwischen 56 Ofo und 60 Ofo auf gleichem Niveau. Bei den Probeflächen, über denen die Altbestände vor 1994 abgestorben sind, ist der überwiegende Teil, nämlich 73 Ofo, verjüngt Prägung durch die Altbestände Grundlage für diese Auswertungen waren die im Rahmen der Waldinventur 1991 terrestrisch aufgenommenen Waldentwicklungsstadien. Die Zuordnung der Waldentwicklungsstadien zu den Verjüngungsdaten erfolgte durch Verschneidung der Waldentwicklungsstadien mit den Probeflächenmittelpunkten. Die Definitionen der Waldentwicklungsstadien sind in Tabelle 3 dargestellt. Die Analyse nach Waldentwicklungsstadien bestätigt, dass die Verteilung der Verjüngung noch stark durch die Struktur der ehemaligen Altbestände beeinflusst wird. So konnten in den 1991 als Jugend- und Verjüngungsstadien klassifizierten Beständen auch noch im Jahr 2000 signifikant mehr Pflanzen größer 20 cm gefunden werden. So betrug die Verjüngungsdichte im Jugendstadium und im Wachstumsstadium Pflanzen je Hektar. Im Wachstums- und Reifungsstadium waren die Verjüngungsdichten mit bzw. 914 Pflanzen je Hektar viel geringer. Auch das Plenterstadium wies mit Pflanzen je Hektar nur eine geringe Verjüngungsdichte auf. Der Anteil verjüngter Probeflächen zeigt ebenfalls dieses Abb. 45 Der Zeitpunkt der Mast ist von entscheidender Bedeutung für eine erfolgreiche Verjüngung der Wälder. 131




131 Abb. 46 Du rchsch n ittl iche Verjüngungsdichte (Balken) und Anteil verjüngter Probeflächen (Linie) der etablierten Pflanzen ab 20 cm im Jahr 2000 in Abhängigkeit vom Flächenanteil der Gras- und Farn-Fazies. Abb.47 Entwicklung der durchschnittlichen Verjüngungsdichte der etablierten Pflanzen ab 20 cm zwischen 1997 und 2000 in Abhängigkeit vom Flächenanteil der Gras- und Farn-Fazies. Abb.48 Durchschnittliche Verjüngungsdichte der Pflanzen zwischen 10 und 20 cm im Jahr 2000 in Abhängigkeit vom Flächenanteil der Gras- und Farn-Fazies. Muster. Während im Jugend- und Verjüngungsstadium 74 bzw. 68 % der Probeflächen Pflanzen aufweisen, sind es im Wachstumsstadium 63 0/0, im Plenterstadium 56 % und im Reifungsstadium 45 Ofo. Allerdings zeigt schon die Entwicklung der Pflanzen größer 20 cm seit 1991, dass der Einfluss der Altbestände immer mehr schwindet und beispielsweise im Reifungsstadium die Pflanzenzahlen fast genauso stark anstiegen wie im Verjüngungsstadium. Auffallend ist, dass im Plenterstadium zurückgehende Verjüngungsdichten auftreten. Die naheliegende Annahme, dass diese Bereiche auf Grund geringeren Kronenschlusses stärker vergrast sind und deshalb eine geringere Verjüngungsdichte aufweisen, konnte nicht bestätigt werden. Betrachtet man die Verjüngungsschicht zwischen 10 und 20 cm wird deutlich, dass der Einfluss der Altbestände immer mehr schwindet, da sich die Individuen viel gleichmäßiger auf die Entwicklungsstadien verteilen. So gibt es zwischen Jugend-, Wachstums- und Reifungsstadium keinen signifikanten Unterschied in der Verjüngungsdichte, nur in den Beständen, die 1991 als Verjüngungsstadium ausgeschieden wurden, findet sich (noch) signifikant mehrverjüngung Konkurrenzvegetation Um die Einflüsse der Vegetation auf die Verjüngungsdichte zu untersuchen, wurden die Flächenanteile der "verjüngungsfeindlichen" Kleinstrukturen, nämlich der Gras-Fazies und der Farn Fazies je Probefläche zusammengefasst und in Beziehung zurverjüngungsdichte gesetzt. Es besteht ein Zusammenhang in der Form, dass mit zunehmender Konkurrenzvegetation die Verjüngungsdichte sinkt. Dieser Zusammenhang ist sowohl für die etablierte Verjüngung als auch für deren Entwicklung seit 1991 und die Pflanzen zwischen 10 und 20 cm Größe signifikant ,..., 4000 ~ 3500 ~ 3000.:g 2500 'ö g, 2000 g, 1500 ~ 1000,-., <11.:E. ~ ~.J:: C.J ~ CI :::l CI s::::: ::::l.~ ~ 'Cä'.:E. ~ ~.J:: C.J ~ CI s::::: :::l CI s::::: ::::l.~ ~ ~ '0 CI s::::: :::l ~ '0 s::::: :<11 ~ 500 o < > 80 Anteil von Gras- und Farn-Fazies (Ofo) < > 80 Anteil von Gras- und Farn-Fazies (Ofo) < >.80 Anteil von Gras- und Farn-Fazies (Ofo) 70 ~ c: QJ 60 ~ e+: 50 ~ o 40 ~ ~ 30 : 20.~ 'öj 10 ~ «o 132



132 Bei den Probeflächen, die weniger als 40 % mit Konkurrenzvegetation bedeckt sind, liegt die Verjüngungsdichte der etablierten Pflanzen durchchnittlich noch über Individuen je Hektar. Steigt der Anteil der Konkurrenzvegetation auf über 60 0/0, sinkt die Verjüngungsdichte auf und bei über 80 % auf nur 833 Pflanzen je Hektar. Noch deutlicher zeigt sich der Einfluss der Konkurrenzvegetation bei der Entwicklung der Verjüngungsdichte der etablierten Pflanzen seit Während auf den Probeflächen, die mit bis zu 40 0 /0 Konkurrenzvegetation bedeckt sind, die Verjüngungszahlen noch um über Pflanzen je Hektar zunahmen, lag dieser Wert in den Kreisen mit mehr als 40 % Konkurrenzvegetation unter 540 Pflanzen je Hektar. Nicht so günstig ist die Verjüngungssituation also auf Probeflächen, die zu mehr als 60 % mit Gras und Farn bedeckt sind. Dies trifft für 408, also 60 % der Probeflächen zu. Auch der Anteil der Probekreise, die Verjüngung aufweisen, sinkt mit zunehmender Flächenbedeckung durch Gras- und Farn-Fazies. Dieser Zusammenhang ist jedoch nicht so stark ausgeprägt, wie bei der Verjüngungsdichte. Während bei weniger als 40 % Flächendeckung durch Konkurrenzvegetation mehr als 65 % der Kreise Verjüngung aufweisen, sind es bei mehr als 80 % nur 41 %. Betrachtet man die Verteilung der Konkurrenzvegetation in Abhängigkeit von der Bodenform, zeigt sich, dass die Böden, auf denen nur eine geringe Verjüngungsdichte vorgefunden wurde, gleichzeitig hohe Anteile an Gräsern und Farnen aufweisen. So liegt der Anteil der Konkurrenzvegetation auf Lehm über verfestigtem Schutt bei 69 %, auf Lehm bei 52 %, auf Block-Humus Böden bei 46 % und auf Nassböden bei 41 %. Ebenso wie die Verjüngungsdichte mit zunehmender Höhenlage sinkt, steigt der Flächenanteil der Gras- und Farn-Fazien. Während er bei Standorten bis 1150 m an der Grenze zum Bergmischwald nur 40 % beträgt, steigt er bis zu den Gipfellagen über 1300 m auf mehr als 70 %. Ein signifikanter Unterschied besteht auch zwischen Böden, die sich aus Gneis und Granit gebildet haben. Böden, die aus Gneis hervorgegangen sind, zeigen eine höhere Bedeckung durch Konkurrenzvegetation und weisen eine geringere Verjüngungsdichte auf. Gras und Farn-Fazies haben somit eine große Bedeutung für die Verteilung der Verjüngung und sind ein Schlüsselfaktor für die Wiederbewaldung in den Hochlagen. 80 -r ~ :B ~ 10 o bis > Meereshöhe (m) e.- 80 T ~ ~ ~- \11 cu 'N ca u.. I E ~ 40 c:: :::l 30 J, ca c!i 20 B c:: 10 c:t: o ' <:... "0 :0.D "&i z Abb. 49 Flächenanteil von Gras- und Farn-Fazies in Abhängigkeit vo'n der Meereshöhe (links) und von der Bodenform (rechts), 133






133 Karte 6 Verjü ngu ngsd ich te der Pflanzen größer 20 cm. Karte 7 Veränderung der Verjüngungsdichte der Pflanzen größer 20 cm zwischen 7997 und Karte 8 Verjüngungsdichte der Pflanzen zwischen 70 und 20 cm. e Verjüngungsdichte bzw. Veränderung der Verjüngungsdichte wengier als bis -1 1 bis bis bis und mehr nicht besetzte Probeflächen Meter _::::::::J_-====:=J 134
134 5. 7 Verteil ungsrn uster in den Hochlagen Die Karten 6 bis 8 zeigen die Verteilung der Probeflächen mit den etablierten Pflanzen größer 20 cm, die Entwicklung seit 1991 und den Pflanzen zwischen 10 und 20 cm, jeweils mit Angabe der Verjüngungsdichte bzw. deren Veränderung. Aufgrund des Versuchsdesigns (kleine Probeflächen, große Abstände der Probeflächen) können nur grobe Angaben über die Verteilung der Verjüngung in den Hochlagen gemacht werden. Auffällig ist, dass östlich des Böhmweges der Anteil unverjüngter Probeflächen weit geringer ist, als in den Bereichen weiter westlich. Auch die Verjüngungsdichte auf den Probeflächen ist dort meist höher. Diese Beobachtung trifft sowohl für die Pflanzen zwischen 10 und 20 cm, als auch für die größer 20 cm zu. Dies hängt vor allem damit zusammen, dass große Bereiche des Gebietes unter 1250 m liegen und dass dort in den höher gelegenen Bereichen die Bestände großflächig unmittelbar nach dem Mast jahr 1995 abgestorben sind. Im mittleren Bereich der Hochlagen zwischen Böhmweg und Steinschachten konzentriert sich die Verjüngung vor allem auf die an den Bergmischwald angrenzenden Hanglagen, dort ist der überwieg.ende Teil der Probeflächen bei meist hohen Pflanzenzahlen verjüngt. Oberhalb des Hangknicks ist die Situation nicht so günstig, insbesondere um den Plattenhausenriegel sind einige Probeflächen ohne Verjüngung. lich damit zusammen, dass große Bereiche des Rachelmassivs über 1250 m Meereshöhe liegen und eine große Bedeckung mit Konkurrenzvegetation aufweisen. Die Probeflächen im Rachelmassiv sind im Mittel zu knapp über 70 Ofo mit Gräsern und Farnen bedeckt. Das ist weit mehr als der Durchschnitt für die gesamten Hochlagen, der nur bei 56 Ofo liegt. Wie zu erwarten, ist deshalb die mittlere Verjüngungsdichte relativ gering, sie beträgt nur 677 Pflanzen je Hektar. Bezieht man allerdings das Potenzial der Pflanzen größer 10 cm mit in die Berechnung ein, stellt sich die Situation viel günstiger dar, die Verjüngungsdichte erreicht dann mit Pflanzen je Hektar einen mehr als doppelt so hohen Wert. Ein Blick auf die Karte mit der Entwicklung der etablierten Verjüngung seit 1991 zeigt, dass bei einigen Probeflächen die Pflanzenzahlen zurückgegangen sind. Diese Punkte sind relativ homogen über die Fläche verteilt. Es besteht ein auffälliger Zusammenhang zwischen dem Rückgang der Verjüngungsdichte und der Verjüngungsdichte der jeweiligen Probefläche. Meist kommt es nur an Punkten mit ohnehin relativ hohen Verjüngungsdichten zu einem Rückgang der Pflanzenzahlen. Obwohl insgesamt Cln 83 Punkten ein Rückgang der Verjüngungsdichte festgestellt werden konnte, trat ein kompletter Verlust aller Verjüngungspflanzen nur in sehr wenigen Fällen auf (2,6 Ofo der Probeflächen). Im Bereich des Rachelmassivs finden sich viele noch unverjüngte Probeflächen ; auch hier konzentriert sich die Ve~jün gung auf die tieferliegenoen Lagen im Verzahnungsbereich mit dem Bergmischwald. Diese Beobachtungen gelten für die etablierte Verjüngung, aber auch für die Pflanzen zwischen 10 und 20 cm. Dies hängt vermut- 135


135 5.8 Uberlebende BaUIllbestände Abb.50 Selbst in ausgedehnten Befallsgebieten wurden einzelne Altbäume und Baumgruppen bisher vom Borkenkäfer verschont. Diese Untersuchung wurde im Bereich zwischen der Abteilung Steinschachten und dem Großen Schwarzbach durchgeführt. Dieses ha große Gebiet wurde ausgewählt, da hier die Massenvermehrung bereits mehr oder weniger zusammengebrochen ist, so dass ein Vergleich der verbleibenden Waldstrukturen mit dem Waldaufbau von 1991 möglich ist. Allerdings konnte insbesondere zwischen Sulzriegel und Kleinem Schwarzbach immer noch frischer Borkenkäferbefall nachgewiesen werden. Die betroffene Fläche war zwar mit 2,3 ha relativ klein, verteilte sich aber auf i nsgesa mt 30 Befa Ilsherde. Die ungebremste Entwicklung des Buchdruckers hat seit 1994 das Bild des Waldes grundlegend verändert: Von den Altbeständen des Reifungs-, Verjüngungs- und Plenterstadiums, die im Jahr 1991 noch 76 % der Fläche bedeckten, haben bis auf einige kleine Rotbuchenbestände nur einzelne Altbäume überlebt. Selbst der Anteil des Wachstumsstadiums ist um 27 % von 11,7 % auf 8,3 0/0 zurückgegangen. Angesichts immer noch vorhandenen Käferbefalls muss hier wahrscheinlich mit einem weiteren Rückgang gerechnet werden. Die noch lebenden Bestände sind nicht älter als ca. 55 Jahre, was in den Hochlagen einem maximalen 136



136 D Jugendstadium Wachstumsstadium Reifestadium o Verj ü ng u ngsstadi u m Plenterstadium o Mortalstadium Sonstiges Abb. 51a u. b Flächenanteile der verschiedenen Waldentwicklungsstadien in einem 1174 ha großen Untersuchungsgebiet innerhalb der Hochlagen. BHD von etwa 20 cm entspricht. 11,7 % der noch vorhandenen Bestände setzen sich aus Laubbäumen zusammen. Von den 93,4 ha, die 1991 als Jugendstadium kartiert wurden, konnten nur 15,5 ha auf den Luftbildern erkannt werden. Ihr Anteil dürfte allerdings weit darüber liegen und sogar die 1991 vom Boden aus als Jungwuchsstadium kartierte Fläche übersteigen. Für die Darstellung der Ergebnisse wurde deshalb die 1991 ermittelte Fläche verwendet. Die Fläche des Mortalstadiums liegt mittlerweile bei 964,3 ha, das sind 83 % des Untersuchungsgebietes. Individuen je Hektar entspricht. Mehr als zwei Drittel (69 0/0) davon sind Fichten, der Rest Laubbäume, vor allem Rotbuchen, aber auch Bergahorne, beide Arten konnten jedoch im Luftbild nicht unterschieden werden. Die überlebenden Fichten sind allerdings nicht gleichmäßig über die Fläche verteilt. Insbesondere in ehemals großflächig gleichförmigen Waldbeständen mit wenigen Strukturen finden sich weniger Individuen als in strukturierten Bereichen mit einem Wechsel von älteren und jüngeren Beständen sowie beigemischtem Laubholz. Besonders interessant ist das Phänomen, dass einige zumeist einzeln oder in kleinen Trupps stehende Bäume den Käferbefall überlebt haben. Es handelt sich dabei vor allem um mittelständige Bäume, aber auch um Mitglieder der ehemals herrschenden Baumschicht. Insgesamt wurden in den Luftbildern Bäume erkannt, was 1,97 137





137 - Hochlagengrenze - I'v'arkierter Wanclerweg - 10m-Höhenlinien Waldflächen _ Jugendstadium _ WachstulT6stadium _ Reifungsstadium Verjüngungsstadi um _ Plenterstadi um Zerfallsstadi um M>rtalstadium Dunbestockt _ Latschen Ni chtwa I dft ächen _ Nichtholzboden _ sonstige Flächen o c===... c=~1 000 tv'eter Karte 9 Waldaufbau





138 -- I-b:hlagengrenze - - M:lrkierter Wanderv.eg -- 10m-l-bhenlinien Überleberde Bnzelbäl.l1le Rchte laubballtl V\aldflächen _ Jugendstadium _ IJ\OChstulT5Stadium MJrtaI stad il.m laubholzbestärde _ latschen Ncht\\eldflächen _ Nchttnlztx:>den _ sonstige Flächen _ Frischer Käferbefall o.r------, ~ 000 IVeter Karte 10 Waldaufbau





139 5.9 Am.eisenkolonien Auf Grund der einfachen Erfassbarkeit wurden nur die Nester von Waldameisen (Formica lugubris) aufgenommen. Insgesamt konnten im Rahmen der Inventur aktive Ameisenkolonien auf 16 verschiedenen Probeflächen festgestellt werden. 13-mal wurde eine, zweimal zwei und einmal fünf Kolonien auf einer Probefläche gefunden waren es noch 32 Kolonien auf insgesamt 28 Probeflächen. Damit ging die Anzahl der Kolonien um fast ein Drittel zurück, die Anzahl der Standorte reduzierte sich um über 40 Ofo. Abb.52 Die Ausbreitung der Gräser gefährdet die Kolonien der Ameisen, da sich durch die Beschattung das spezifische Mikroklima im Nestinnern verändert. Bereits bei der Waldinventur 1991 wurde die Anzahl aktiver Ameisenkolonien auf den 500 m' großen Probeflächen bestimmt. Um Informationen über die Auswirkung der großflächigen Absterbeprozesse auf diese waldtypischen Arten zu erhalten, wurden die Aufnahmen 2000 wiederholt. Schwerpunkt der Verbreitung sind die Distrikte Enzianruck und Hochwald. Im Rachel-Bereich konnten nur jeweils zwei Nester nachgewiesen werden. Während sich die Kolonien im Bereich Enzianruck behaupten konnten, wurde im Gebiet des Distriktes Hochwald ein starker Rückgang der Bestände beobachtet. o o Karte 77 Vergleich der Ameisenvorkommen auf den Probekreisen zwischen den Jahren 7997 und o 'Kolonien 1991 o 1 o 2 Kolonien o o o Meter I 00 o ~ o 140


140 6.1 Der alte Bergfichtenwald ließ man Anfang der neunziger Jahre seinen Blick vom Lusengipfel über die Wälder schweifen, schien die Welt für den Betrachter noch in Ordnung. Es war ein beeindruckendes Bild: dunkelgrüne Fichtenwälder, die über die sanften Gipfel des Grenzkammes wogen und Bergmischwälder, die sich talwärts nahtlos anschlossen. Wald so weit das Auge blicken konnte. Nur vereinzelt war das grüne Wäldermeer von kleineren Windwürfen und Totholzinseln durchbrochen. Beim Abstieg durchwanderte man relativ gleichförmige Wälder, die fast ausschließlich durch alte Fichtenbestände geprägt waren hatte die Fichte einen Anteil von 98 % an der Gesamtbestockung. Die verbleibenden 2 % verteilten sich auf Rotbuche (1,2 0/0). Vogelbeere (0,4 0/0), Bergahorn (0,2 0/0) und sonstige Baumarten (0,2 0/0) (RALL 1995). Nur selten öffneten sich die alten Bestände und man traf auf kleinere Verjüngungsgruppen oder Dickungen, die allesamt noch aus der Zeit der forstlichen Bewirtschaftung stammten. Seit Unterschutzsteilung des Gebietes waren die Holzvorräte von Jahr zu Jahr angewachsen und die Wälder kontinuierlich älter und dadurch sowohl aus ökonomischer als auch naturschutzfachlicher Sicht stetig wertvoller geworden (HABER 1976). So lag der Holzvorrat mit 381 Festmetern je Hektar 1991 auf einem seit Beginn der Forstwirtschaft nie erreichten Niveau. Im Vergleich dazu erscheinen die Holzvorräte der bewirtschafteten Hochlagenwälder in den benachbarten Forstämtern mit im Durchschnitt 249 fm/ha (NUESSLEIN 1996) relativ gering. Auch "urwaldartige" Bestände, die Mitte des 19. Jahrhunderts aufgemessen wurden, hatten bei Weitem nicht so hohe Holzvorräte wie ZIERL (1972) gibt für sie einen Durchschnittsvorrat von nur 265 fm/ha an. Die Veränderung des Altersklassenaufbaus seit 1855 verdeutlicht diese Entwicklung. Während damals noch 41 % der Bestände unter 80 Jahre waren, hat sich deren Anteil bis 1991 auf 14 % reduziert. Im Gegenzug stieg der Anteil der Bestände mit einem Alter von über 120Jahren von 47 auf69 0/0. Auch die einzelnen Bestände waren sehr homogen und einschichtig aufgebaut, trotz zum Teil beträchtlicher Altersunterschiede. ZIERL (1972) konnte z. B. auf einem 340 m langen Trassenaufhieb eine Altersdifferenz von ca. 200 Jahren feststellen. Dies hängt vor allem mit der hohen Lebenserwartung der Fichten im Bergwald zusammen. Sie liegt bei über 300 Jahren. Während der Waldinventur 1991 wurde für die Hochlagen ein Durchschnittsalter von 280 bis 300 Jahren ermittelt. Die älteste beprobte Fichte war 380 Jahre (RALL pers. Mitteilung). Auch KORPEL (1995) berichtet von einem 350- bis 400-jährigen Lebenszyklus der Fichtenurwälder in den Westkarpaten. Da die älteren Bäume ihr Wachstum verlangsamen und die jüngeren schneller in die verbliebenen Bestandeslücken vorstoßen, wachsen sich die anfangs unterschiedlichen Baumhöhen während des langen Bestandeslebens langsam aus und es bilden sich fast einschichtige Bestände. Da im Hochwald schattenertragende Baumarten nur sporadisch vorkommen und die Fichte relativ viel Licht benötigt, können sich nur sehr Abb.53 B!ickvom Lusen zum Rache! imjahr


141 80 70 '"""',e 60 ~ ~ CI:! 50 ~ ~ 40 <... 4J "'C 30 '4j +-' s:::: <C bis 40 Abb.54 Veränderung der Flächenanteile der verschiedenen Altersstufen seit 1855 (ELLING 1987) >120 Alter in Jahren wenige Bäume im Unter- und Zwischenstand behaupten. Auch die horizontale Struktur der Bestände ist nicht besonders ausgeprägt, da die für den Bergwald typische Rottenstruktur im Laufe der Bewirtschaftung teilweise aufgelöst wurde. Erst wenn auch der Waldboden in die Betrachtung mit einbezogen wird, ergibt sich ein vielfältigeres Bild. Da das Kronendach nicht dicht geschlossen ist, dringt viel Licht in die Bestände, was am Boden einen Wechsel von Licht und Schatten erzeugt. In den lichten Bereichen breiten sich weite Grasteppiche und Heidelbeerbestände aus, in schattigeren, feuchteren Bereichen wachsen bis mannshohe Farnbestände und dort, wo die Bäume enger stehen, ist die Rohhumusdecke frei von Vegetation. An steileren Stellen findet man Blockböden mit einem Mosaik aus moosbedeckten Felsen, Humusansammlungen und Heidelbeeren. Nur vereinzelt liegen Rannen, das sind vermodernde Baumleichen, am Boden oder finden sich umgestü rzte Bäume mit aufgeklappten Wurzeltellern. Die Waldverjüngung war 1991 mit 978 Pflanzen je Hektar noch recht spärlich vertreten. Auffällig ist der hohe Anteil der Vogelbeere, die damals schon unter der geschlossenen Baumschicht wuchs. Auch KORPEL (1995) teilt mit, dass die Vogelbeere in den Urwäldern der Westkarpaten insbesondere im Verjüngungsstadium einen hohen Anteil hat, der sogar den der Fichte oft übersteigt. Die Fichtenverjüngung war relativ gleichmäßig auf die Höhenklassen verteilt, während die Vogelbeerenverjüngung mit zunehmender Größe immer weniger wurde (Verbiss). Nennenswertes Wachstum zeigte nur die Fichtenverjüngung, die in größeren Lücken wuchs. In geschlossenen Bestandesteilen konnten keine oder nur sehr kleine Gipfeltriebe beobachtet werden. Die von PRZYBILLA (1993) und GEITNER (2000) in geschlossenen Bestandesteilen ermittelten Höhenzuwächse lagen bei etwa einem Zentimeter pro Jahr. Entsprechend diesem geringen Höhenzuwachs weisen schon relativ kleine Bäume ein hohes Alter auf. Eine von ZIERL (1972) gefällte 2,5 m hohe Fichte war beispielsweise 91 Jahre alt. Verantwortlich für dieses langsame Wachstum ist vor allem die kurze Vegetationszeit, die durch das geringe Wärmeangebot im Bestandesinneren und die lange andauernde und hohe Schneebedeckung beeinflusst wird. Insbesondere das Wurzelwachstum ist unter diesen Bedingungen stark verlangsamt. Die dafür optimalen Bodentemperaturen von 16 bis 26 oe werden in den Berglagen nur selten erreicht (On et al. 1997). 142



142 Abb.55 Blick vom Lusen zum Rachel im Jahr Abb.56 Blick vom Lusen zum Rachel im Jahr


143 6.2 Sukzession - die "Erste" Abb.57 Der Mulcheffekt durch herabgefallene Rinde führt zu günstigen Wachstumsbedingungen für die Waldbäume bahnte sich eine Entwicklung an, die den Bergfichtenwald dramatisch verändern sollte. Zunächst kleinflächig, eher schrotschussartig, breitete sich der Buchdrucker über weite Bereiche der Hochlagen aus "explodierte" die Käferpopulation förmlich und innerhalb einesjahres starben in den Hochlagen 310 ha Fichtenaltbestände ab. Ihren Höhepunkt erreichte die Käferentwicklung ein Jahr später, als 583 ha Altbestände "aufgefressen" wurden. Anschließend ging der Befall in den Hochlagen parallel zum Verschwinden der Altfichten langsam zurück, so dass im Herbst 2000 dort nur noch 15 % Prozent der Fichtenbestände intakt waren (HEURICH et al ). Schlagartig änderten sich die Standortsbedingungen. Zuerst fielen die Nadeln der abgestorbenen Bäume zu Boden und bildeten dort eine mehrere Zentimeter dicke Schicht. Jetzt konnte die Sonnenstrahlung bis zum Waldboden vordringen, wodurch dieser sich erwärmte und die Mineralisation der Nadel- und Wurzelstreu und wahrscheinlich auch der dicken Rohhumusauflagen eingeleitet wurde. Vor allem Schwarzspechte lösten auf der Suche nach Nahrung große Rindenstücke von den abgestorbenen Bäumen ab. Diese konzentrieren sich dann zusammen mit den nach und nach abfallenden Ästen um den Stammfuß, was zu einer teilweisen Bodenbedeckung und zu einer starken Anreicherung von Nährstoffen, vor allem Kalzium und Phosphor, führt (KIMMINS 1987, BEUDERT 1994). Unter den herabfallenden Rindenstücken und Ästen werden zwar einige Fichten, besonders Sämlinge und Keimlinge begraben. Durch Mulcheffekt und Nährstoffeintrag entstehen günstige Bedingungen für die Entwicklung der Waldbäume, aber auch für die Entwicklung anderer Pflanzenarten, wie Weidenröschen (Epilobium angustifolium) und Himbeere (Rubus idaeus), so dass sich in der Umgebung der alten Stämme grüne Zonen mit vielen blühenden Pflanzen bilden. Diese Entwicklung wird auch durch das günstige Mikroklima im Bereich der Stammanläufe unterstützt. Beispielsweise apern die stammnahen Bereiche im Frühling sehr bald aus und es bilden sich Schmelztrichter, was eine verlängerte "Vegetationszeit" zur Folge hat. Ob diese Schmelztrichter auch als Sammelanlagen für die im Spätwinter oft über die Schneedecke verblasenen Fichtensamen fungieren, müssen weitere Studien zeigen. Das Ausmaß dieser Entwicklungen zeigen die im Rahmen der Inventur durchgeführten Aufnahmen. So nahm die Kleinstruktur Kräuter und Himbeere gegenüber der Aufnahme 1996 um fast 70 0 /0, die Kleinstruktur Reisig/Rinde gar um über 150 Ofo zu. Dass in der Kleinstruktur Kräuter und Himbeere relativ wenig Verjüngung gefunden wurde, hängt vermutlich damit zusammen, dass sich dieses Element zum Zeitpunkt der Aufnahme 1996 erst ausbildete. Zwischen Himbeere und Waldverjüngung scheint in der Hochlagenzone nur eine geringe Konkurrenz zu bestehen, was auch SIEGRIST (2000) bestätigt. Auf Reisig/Rinde verjüngte sich die Vogelbeere neunmal mehr, als dies auf Grund des Flächenanteils der Kleinstruktur zu erwarten gewesen wäre. Eine Erklärung für 144


144 diese Beobachtung ist neben den oben beschriebenen Effekten wohl auch der durch Äste gegebene Schutz vor Wildverbiss und der Eintrag von Samen, da diese Kleinstruktur bevorzugt von Vögeln genutzt wird. Es wird erwartet, dass sich die positiven Effekte dieser Kleinstruktur in naher Zukunft noch stärker auswirken. Bereits nach wenigen Jahren fangen die Gipfel der abgestorbenen Bäume an herabzubrechen. Hat sich die Verpilzung weiter ausgedehnt, bricht der ganze Stamm entzwei und stürzt zu Boden. Dies kann je nach Standort bereits nach zwei bis drei Jahren erfolgen; es sind aber auch Bereiche bekannt, in denen die ausgetrockneten, stehenden Dürrlinge, die kurz nach den Windwürfen 1983/84 vom Borkenkäfer befallen wurden, bis heute überdauerten. Übrig bleiben ca. 3 bis 7 m hohe Stümpfe und ein Gewirr aus übereinander liegendem und ineinanderverkeiltem Totholz. Liegendes Totholz hat eine besondere Bedeutung für die Verjüngung wuchs ein großer Teil der Fichtenpflanzen auf bzw. im Schutz dieser Kleinstruktur. Rannen im eigentlichen Sinn gab es damals allerdings nur wenige, da sie im Zuge der Bewirtschaftung der Wälder vor Gründung des Nationalparkes entfernt worden waren. Deshalb wuchs der überwiegende Teil der Fichten auf alten Stubben, die noch aus der Zeit der Bewirtschaftung stammten. Mittlerweile erhöht sich das Angebot von Totholz auf dem Waldboden von Jahr zu Jahr und erste Stämme werden bereits von Fichtensämlingen besiedelt. Ob diese sich bereits auf dem noch nicht stark zersetzten Totholz etablieren können, werden die nächsten Jahre zei-gen. Insbesondere der Witterungsverlauf wird hier eine große Rolle spielen. Bisher konnte erfolgreiche Rannenverjüngung nur auf liegendem Totholz, das seit mindestens 13 Jahren umgestürzt ist, nachgewiesen werden (JEHL 2001). Günstig ist aber schon jetzt, dass der Totholzverhau die Verjüngung vor Schneeschub und z. T. vor Wildverbiss schützt. Durch die Beruhigung der bodennahen Luftschichten und seiner Schattenwirkung trägt der Verhau auch zu einem ausgeglicheneren Kleinklima bei. Darüber hinaus wird der Boden vor Austrocknung geschützt. Die große Bedeutung des Moderholzes für die Waldentwicklung wird dadurch belegt, dass /0 der Pflanzen auf oder in unmittelbarer Umgebung von dieser Struktur wuchsen. Auch andere Autoren weisen immer wieder auf die große Bedeutung von Moderholz für die Fichtenverjüngung hin, wobei bis zu 65 % der Pflanzen auf dieser Kleinstruktur wachsen (ZIERL (1972), LEIBUNDGUT (1978), On et al. (1997). KORPEL (1995), SCHMIDT VOGT (1991). RALL (1995), REIF und PRZYBILLA (1998)). Die Kleinstruktur Moderholz bringt für die hier keimenden Fichten eine ganze Reihe von Vorteilen, die in JEHL (2001) beschrieben sind. Im Gegensatz zur Fichte hat das vermoderte Holz keine Bedeutung als Wuchsort für die Vogelbeere. Nur im Baumkontakt, also der unmittelbaren Umgebung stehender und liegender Bäume, findet man sie häufiger. Abb. 58 Mit Hilfe der Strategie, sich auf Moderholz zu entwickeln, kann sich die Fichte auch stark mit Farn und Gras bedeckte Bereiche erschliessen (Rannenboot im Grasmeer). 145
145 Günstige Verjüngungsstandorte sind auch Humus, Moos und die Zwergsträucher. Dies gilt gleichermaßen für die Fichte und die Vogelbeere. Die Bedeutung der Kleinstruktur Humus dürfte aktuell größer geworden sein, da sie vor dem Borkenkäferbefall vor allem in dicht geschlossenen Bestandesteilen vorkam und sich hier erst durch die Freistellung attraktive Entwicklungsbedingungen für die Verjüngung boten. Am wenigsten Verjüngung findet man in den Farn- und Gras Fazies, da hier die Entwicklung der Pflanzen gehemmt wird. Im Herbst sterben die oberirdischen Teile der Farne und Gräser und werden durch den Schnee zusammen mit den Samen zu Boden gedrückt. Dieses dichte Geflecht erschwert es den Keimlingen, ihre Wurzel bis in den Mineralboden vorzutreiben, zudem sterben viele Samen und Keimlinge in diesem Milieu durch Pilzbefall und Fäulnis. Die Begründung, dass es für die kleinen, leichten, geflügelten Fichtensamen fast unmöglich ist, den dichten Grasfilz zu durchdringen, um auf den Boden zu gelangen (FISCHER 1998) und deshalb hohe Ausfallquoten auftreten, gilt auf Grund der hohen Schneelage vermutlich nicht für Gebirgswälder. Selbst wenn den Keimlingen der Anschluss an den Mineralboden gelingt, konkurrieren sie nun mit den Wurzeln der Gräser und Farne und bekommen in der dichten Vegetation nur sehr wenig Licht und Wärme ab (PLATE 1975). Wenn dann am Ende der Vegetationszeit die oberirdischen Organe der Gräser und Farne abgestorben sind und der Schnee sie auf den Boden drückt, bedeutet das zumeist auch das Ende der bis zu diesem Zeitpunkt überlebenden Keimlinge. Mit der plötzlichen Belichtung und der raschen Mineralisierung von Stickstoff nach dem Absterben der Baumschicht beginnt dann ein Wettlauf der Verjüngung mit der Konkurrenzvegetation (OTT et ai.1997). Allerdings scheint deren Ausbreitung über Ausläufer doch nicht so rasch zu verlaufen, so dass die Verjüngung noch einige Jahre Zeit hat, sich zu etablieren. Die Ausdehnung der Gras- und Farn-Fazien ging vor allem zu Lasten der Kleinstrukturen Moos, Humus und Zwergsträucher. Auch der bisher schon geringe Anteil der Kleinstruktur Wurzelteller ging vor allem durch die Ausbreitung der Gras-Fazies in die Wurzeltellermulde weiter zurück. Der Anteil dieser Kleinstruktur ist mittlerweile so gering, dass sie für die Verjüngung keine bedeutende Rolle spielt. Die Ausdehnung der Gras- und Farn-Fazien hat aber neben den negativen Auswirkungen auf die Verjüngung auch einen sehr positiven Effekt: Der Boden wird abgedeckt und der dichte Wurzelfilz schützt ihn wirksam vor Erosion. Auffällig ist auch der starke Rückgang der Bärlappe um fast 40 0 /0. Die Gründe für diesen Rückgang sind noch unklar. Eine mögliche Ursache könnte der Mangel an Nährelementen (WAIER 1996) oder die Konkurrenzschwäche der Bärlappe sein. Für die im alten Wald schon vorhandenen Verjüngungspflanzen verbesserte sich die Situation nach dem Ausfall der Beschattung durch die alten Bäume entscheidend. Es gelangte mehr Licht und Wärme auf den Boden, wodurch die Stoffwechselvorgänge und hier vor allem das Wurzelwachstum angeregt wurden. Jetzt kam die Stunde der Vogelbeere, die jahrelang im Schatten des Altbestandes ausgeharrt hatte, sie schoss geradezu in die Höhe. Ihre Jahrestriebe betrugen auf der Freifläche im Mittel 18 cm, es wurden aber auch Zuwächse von fast einem halben Meter beobachtet. Die Reaktion der Fichten war verhaltener, sie legten im Mittel um 5 cm zu. Allerdings konnten auch hier Maximalwerte von bis zu 14 cm beobachtet werden (GEITNER 2000). Auf einer Dauerbeobachtungsfläche von JEHL (2001), über der die Bestände bereits Ende der 80er Jahre abgestorben waren, erreichte die Fichte sogar einen durchschnittlichen Höhenzuwachs von 24 cm pro Jahr. 146
")


146 147 (Abb. 59)
 wies nach, dass auch nach der Mast 1995 eine beträchtliche Stammzahlreduktion in der Verjüngung ablief, diese war aber nicht so stark ausgeprägt wie in der Schweiz: von 29.")

147 6.3 Sukzession - die "Zweite" erging es auch den Pflanzen, die aus dem Mastjahr 1958 im Bayerischen Wald hervorgingen, trotz aufwendiger Bodenbearbeitung und Düngung konnten nur sehr wenige überleben. GEITNER (2000) wies nach, dass auch nach der Mast 1995 eine beträchtliche Stammzahlreduktion in der Verjüngung ablief, diese war aber nicht so stark ausgeprägt wie in der Schweiz: von erfassten Keimlingen 1998, waren im Folgejahr nur noch vorhanden. Auch die Ergebnisse der Inventur 2000 lassen vermuten, dass die Überlebenswahrscheinlichkeit der Pflanzen relativ hoch ist, da sehr viele Fichten die Erfassungsschwelle von 10 cm erreichten. Abb.60 Obwohl die Vogelbeere recht zahlreich auf den vor längerer Zeit abgestorbenen Flächen auftritt, können sich keine geschlossenen "Vogelbeerenvorwälder" herausbilden. Trotz der positiven Reaktion der schon vor den Absterbeprozessen vorhandenen Verjüngungspflanzen auf den Ausfall der Baumschicht war die bei den Inventuren 1996 und 1998 ermittelte Verjüngungsdichte noch relativ gering. Erst bei der Inventur 2000 konnte ein starker Zuwachs bei der Verjüngungsdichte, insbesondere bei den kleinen Pflanzen festgestellt werden. Der Grund für diese Entwicklung lag allerdings schon einige Jahre zurück. Es war die starke Fichtenblüte und erfolgreiche Zapfenanlage des Jahres 1995, die im Frühsommer des Folgejahres einen Teppich von Fichtenkeimlingen auf günstigen Kleinstandorten hinterließ. Allerdings waren diese "Miniaturbäumchen" einer großen Anzahl von Gefahren ausgesetzt. Da die kleinen Wurzeln nicht in der Lage sind, die Humusschicht bis zum Mineralboden zu durchdringen, können bereits kurze Trockenperioden zum Absterben Tausender von Keimlingen führen. Auch Schneepilze und Schneeschub können zahlreiche Pflänzchen abtöten. Von Keimlingen je Hektar, die On. et al. (1997) 1982 in einem Hochstauden-Fichtenwald bei Davos ausgezählt hatte, waren Ende 1984 schon 84 0J0 abgestorbe~. Als die Aufnah-. men neun Jahre später wiederholt wurden, waren keine Keimlinge mehr übriggeblieben. Ähnlich Dass mehr Keimlinge überlebten wie im Schweizer Beispiel und in der Vergangenheit, hängt vermutlich mit dem Borkenkäferbefall zusammen. Wie die Analyse der nach Absterbejahren stratifizierten Probeflächen zeigte, konnten sich vor allem dort viele Verjüngungspflanzen etablieren, wo die Altbestände 1996 abgestorben waren. Auch wenn auf diesen Flächen viele Keimlinge durch herabfallende Nadeln überdeckt wurden und abstarben, führte das verbesserte Licht- und Wärmeangebot dazu, dass überdurchschnittlich viele Keimlinge überlebten. Doch schon auf den Flächen, über denen die Altbäume ein Jahr später abstarben, konnte im Durchschnitt nur eine halb so hohe Verjüngungsdichte nachgewiesen werden. Das Mast jahr 1995 hat also zu einem enormen Zuwachs der Pflanzenzahlen geführt, der sich auch bei der nächsten Inventur fortsetzen dürfte. Ergebnisse von Dauerbeobachtungsflächen deuten darauf hin, dass mehr als die Hälfte der Individuen die Erfassungsschwelle von 10 cm noch nicht erreicht haben (BAUER pers. Mitteilung). Im Vergleich zur Fichte verläuft die Entwicklung bei der Vogelbeere viel gemächlicher, da die Samenproduktion bei dieser Baumart mehr oder weniger kontinuierlich und auf Grund der wenigen 148
148 Altbäume auf geringerem Niveau vonstatten geht und nicht auf wenige Mast jahre mit zum Teil großem Abstand beschränkt ist. Folge war, dass der Anteil (nicht deren Anzahl) der Vogelbeere in der Verjüngung immer weiter zurückging. Auch ist sie bei Weitem nicht so homogen über die Hochlagen verteilt wie die Fichtenverjüngung, was mit den nur wenigen alten Vogelbeeren und der zoochoren Ausbreitungsstrategie, die zu einer starken Klumpung der Individuen führt, erklärt werden kann. Der Anteil der Rotbuche hat sich in den vergangenen Jahren nur geringfügig verändert. Obwohl immer wieder diskutiert wird, dass die prognostizierte Klimaveränderung zu einer Arealausbreitung der Rotbuche in die Hochlagenwälder führen könnte, wird dies doch noch ein sehr langer Prozess sein, da sie nicht mit den klimatischen Bedingungen der Freiflächen zurecht kommt. Deshalb konzentriert sich die Buchenverjüngung zur Zeit vor allem auf bergmischwaldnahe Bereiche, in denen überlebende Altbuchen die Klimaextreme abmildern. Auch die Ausbreitung der Gräser auf den Freiflächen, die meist mit hohen Mäusedichten einhergeht, ist einem Anstieg des Rotbuchenanteils abträglich, da vermutlich viele Bucheckern gefressen und auch die Rinde von Verjüngungspflanzen abgenagt werden, was zu hohen Ausfällen führen kann. Die Rotbuche als Schattenbaumart, die ein ausgeglichenes WaIdinnenklima benötigt, könnte erst dann weiter in den Bergfichtenwald vordringen, wenn sich ein Vorwald aus Vogelbeere und/oder locker stehenden Fichten bilden würde. Andere Baumarten kommen in den Hochlagen des Inneren Bayerischen Waldes mit nur wenigen Individuen vor und spielen bisher keine Rolle im Wiederbewaldungsprozess. Dies hängt auch mit der Tatsache zusammen, dass die Pionierbaumarten sich vor allem dort verjüngen, wo der mineralische Rohboden freigelegt ist (LAESSIG und SCHOENENBERGER 1993, HOMANN und ENGELS 1991). Solche Kleinstrukturen treten allerdings nach Borkenkäferbefall, im Gegensatz zur Situation nach Windwürfen, kaum auf. Die Entwicklung nach dem großflächigenabsterben des Altbestandes verläuft somit nicht gemäß dem Modell der sekundären Sukzession, d. h.dass sich zunächst ein Anfangs- odervorwaid aus Pionierbaumarten entwickelt, der dann von der Klimaxbaumart unterwandert wird (Übergangswaid) und sich anschließend zum Schlusswald entwickelt (LEIBUNDGUT 1978, KORPEL 1993, ZUKRIGL 1963). Die in der Verjüngungsschicht des Altbestandes vorhandenen Vogelbeeren wachsen den Fichten zwar voraus, stehen aber meist so weit auseinander oder fehlen auf großen Flächen, so dass man nicht von Vorwald sprechen kann. Ein eher dem Modell der sekundären Sukzession entsprechender Ablauf könnte sich eventuell auf den für die Fichte verjüngungsökologisch ungünstigen Standorten, also vor allem den vergrasten Bereichen ergeben, denn hier ist zunächst nur die Vogelbeere in der Lage zu keimen und zu wachsen. Im Laufe der Zeit wird sie vermutlich selbst großflächige Graspartien besiedeln. Der verminderte Lichteinfall unter dem Schirm der Vogelbeere, verbunden mit alljährlichem Laubabwurf, könnte allmählich zum Zurückdrängen der Gräser führen und ein Ansamen der Fichte ermöglichen. Diese Vogelbeervorwälder böten dann besonders gute Bedingungen für die Entwicklung der Fichtenverjüngung. Darüber hinaus trägt die Vogelbeere auf Grund ihrer besonders kalzium- und phosphatreichen Streu zu einer Steigerung der biologischen Aktivität im Boden und hier insbesondere im Auflagehumus bei (LEDER und PETRAK 2000). Bisher konnte die Bildung solcher Vorwälderjedoch noch nicht beobachtet werden. 149
 sind Pionierbaumarten die ersten, die eine nicht (mehr) mit Wald bedeckte Fläche besiedeln.")

149 Abb.61 Die Verteilung der günstigen Kleinstrukturen führt dazu, dass die Fichten, oft als kleine Trupps im Verband aufwachsen Nach SCHurn et al. (1992) sind Pionierbaumarten die ersten, die eine nicht (mehr) mit Wald bedeckte Fläche besiedeln. Sie produzieren eine große Menge leichter, vom Wind transportierter Samen und sind relativ unempfindlich gegen Spätfröste, starke Einstrahlung, Dürre und Wind. Sowohl Fichte als auch Vogelbeere entsprechen in weiten Teilen dieser Definition. Allerdings fehlt bei der Fichte die Dürreres.istenz und bei der Vogelbeere werden die Samen nicht vom Wind, sondern über Zoochorie verbreitet. Zudem scheint sie schattentoleranter als die Fichte (KORPEL 1995). Die beschriebenen Sukzessionsverläufe stimmen also nur bedingt mit den Modellvorstellungen der sekundären Sukzession überein, weil es im Bergfichtenwald - abgesehen von den geringen Anteilen Rotbuche - keine exakte Trennung zwischen Pionier- und Klimax-, zwischen Licht- und Schattenbaumarten gibt. Die Fichte nimmt in diesem System eine "Zwitterrolle" ein, sie ist gleichzeitig Pionier- und "Klimaxbaumart". Die Vogelbeere bildet auf dem überwiegenden Teil der Fläche keinen klassischen Vorwald aus, sondern ist nur Begleiter der Fichte, was letztendlich dazu führt, dass die klassischen Sukzessionsmodelle hier nicht greifen. Die Entwicklungsdynamik des Ökosystems wird vermutlich auch in Zukunft ganz entscheidend von der Fichte bestimmt, auch, weil die Vogelbeere nur in etwa die halbe Lebensdauer der Fichte besitzt (KORPEL 1995). Verteilung und Veränderungen der Kleinstrukturen steuern in ganz entscheidendem Maß die Verteilung der Waldverjüngung und sind Grund für ihr geklumptes Auftreten. Die Verjüngung konzentriert sich auf den oben beschriebenen günstigen Standorten. Dort wurden bis zu 16 Pflanzen größer 10 cm pro Quadratmeter gefunden, während die Grasteppiche in direkter Nachbarschaft fast verjüngungsfrei sind. Die kleinen Trupps und Rotten werden deshalb im Verband aufwachsen, wo sie sich gegenseitig stützen und eine "Kampfgemeinschaft" bilden, die innerhalb kurzer Zeit über ein eigenes Innenklima verfügen wird und die Konkurrenzvegetation ausdunkelt. Obwohl der Mensch nicht mehr direkt in die Verjüngungsprozesse eingreift, werden sie noch in erheblichem Umfang durch die Waldbehandlung vor 1970 determiniert. Bestände, die damals sehr vorratsreich waren, weisen heute eine geringere Verjüngungsdichte auf, ehemalige Verjüngungsund Jugendstadien eine höhere. Auch wenn für die Verjüngungsdichte der 1996 gekeimten Pflanzen kein Zusammenhang zu den 1991 vorgefundenen Beständen mehr besteht, bleibt ein Zusammenhang im kleineren Maßstabsbereich bestehen. "Hot Spots" der Verjüngung sind nämlich nach wie vor alte vermoderte Stöcke und die Bereiche um die abgestorbenen Bäume. Die Verteilung dieser Kleinstrukturen wurde aber zu einem erheblichen Teil durch die im Hochwald wirkenden Forstleute bestimmt. Klassische Rannenverjüngung auf liegenden Stämmen, die diese Determinierung durchbrechen könnte, kommt bisher nur in geringem Umfang vor. Es wird also zumindest noch eine Waldgeneration dauern, bis der Wald in den Hochlagen als vollkommen "wild" bezeichnet werden kann und das Erbe der Bewirtschaftung abgeschüttelt hat. 150



. damit ~btöteten. D.")
150 6.4 "Schäden" an der Verjüngung Abb.62 Verbiss durch den Rothirsch ist ein wichtiger Faktor für die Entwicklung der Vogelbeeren verjüngung. Insgesamt konnten nur sehr geringe "Schäden" an den Verjüngungspflanzen festgestellt werden. Die Befürchtungen, dass der Buchdrucker in größerem Umfang Verjüngungspflanzen zum Absterben bringen könnte, haben sich nicht bestätigt. Diese Schäden traten bisher nur 1997 v. a. im Bereich des Lusens auf, nachdem dort zuvor große, zusammenhängende Altholzbereiche abgestorben waren und sich die Käfer auf der Suche nach Nahrung selbst in junge Fichten einbohrten und her auf den Probeflächen nicht nachgewiesen werden. Ein Grund dafür könnte das raschere Abschmelzen der Schneedecke auf den Totholzflächen auf Grund der relativ hohen Frühlingstemperaturen in den vergangenen Jahren sein. Dass der Schneeschimmel auch im Bayerischen Wald eine große Rolle spielen kann, zeigen massive Ausfälle an bis zu 2 m hohen Fichten, die Anfang der 80er Jahre beobachtet wurden (SCHERZlNGER pers. Mitteilung). damit ~btöteten. D.ie starken Schneebruchereignisse der 'letzten Obwohl immer wieqer Fraßsteilen an Ästchen der beiden Jahre hatten wohl,auf Grund der noch geringen Naturverjüng~ng auffal'len, sind die Beschädigungen Größe der Pflanzen ebenfalls keine größe- du~ch Rüsselkäfer im Untersuchun,gs- ren Beschädigungen zur Folge. Auch Rindenfraß gebiet praküsch'bedeutungslos.die Befürchtung, ', durch Mäuse war bedeutungslos. dass si~h die Käfer im Wurzelsystem der abgestor- < ben~en.bäume ähnlich erfolgreich' ver,mehren könnten wiein _ ~en -Störken, frisch gefällter Bäu-', me, hat sich somit nicht bestätigt. Im Gegensat~ zu den Beobachtungen im Rathel-Lusen-Gebiet konnten im Falkenstein-Rachel-Gebiet, wo vom Buchdrucker befallene Bäume entnommen werden, deutlich mehr Schäden durch Rüsselkäfer beobachtet werden (BAUER, pers. Mitteilung). Abb.63 Durch das Zusammenbrechen der abgestorbenen Bäume bilden sich Verhaue, die die Raumnutzung des Schalenwildes beeinflussen. Auch der Schneeschimmel, der in der Literatur oft als eine der wichtigsten Ausfallursachen der Verjüngung beschrieben wird (OTT et al. 1997, SCHOENENBERGER 1988, BRANG 1996), konnte bis- 151


151 Der Leittriebverbiss bewegt sich auf einem sehr niedrigen Niveau. Nur bei der beim Wild beliebten Vogelbeere spielt er eine Rolle. Obwohl dort der Verbiss in den letzten Jahren stark zurück gegangen ist, muss er doch in den nächsten Jahren genau beobachtet werden. Ein Problem liegt nämlich darin, dass es sich bei der Vogelbeere in den Hochlagen vor allem um Sommerverbiss handelt, der sich im Laufe des Sommerhalbjahres aufsummiert. Folglich wird die Höhe des Leittriebverbisses entscheidend vom Aufnahmezeitpunkt beeinflusst: Je später im Sommer gemessen wird, desto höher dürfte der Leittriebverbiss ausfallen. Da die Inventuren meist im Frühsommer durchgeführt werden, wird der Leittriebverbiss möglicherweise unterschätzt. Es wird vermutet, dass sich durch das Zusammenbrechen der Altbäume im Laufe der Zeit ein Verhau bildet, der die Raumnutzung des Schalenwildes beeinträchtigt und damit zu einer kleinräumlich differenzierten Verbisssituation führt. Detailstudien sollen genaueren Aufschluss über diesen Prozess geben. Da sich der Begriff Schäden eigentlich auf wirtschaftliche Verluste bezieht und deshalb sehr negativ besetzt ist, sollte man ihn in Nationalparken, in denen keine wirtschaftsbedingten Nutzungen erfolgen, nicht verwenden. Denn in natürlichen Systemen gibt es keine Schäden im eigentlichen Sinn. Es handelt sich vielmehr um Interaktionen von Lebewesen mit ihrer belebten und unbelebten Umwelt, die neutral bewertet werden müssen. Als natürliche Selektionsmechanismen haben sie eine entscheidende Bedeutung im Evolutionsprozess. Zu Schäden werden sie erst, wenn menschliche Interessen, wie z. B. Minimierung von Kosten für die Waldverjüngung, beeinträchtigt werden. Leider findet sich im deutschen Sprachraum kein Begriff, mit dem man das Wort "Schäden" zufriedensteilend ersetzen könnte. Abb.64 Im Bereich des Rachelmassivs sind weite Flächen mit Gräsern bedeckt, die Waldverjüngung konzentriert sich hier v. a. auf die Kleinstrukturen Baumkontakt und Moderholz. 152
152 6.5 Muster der Verjüngungsverteilung im. Gelände Wie bereits diskutiert, sind Gras- und Farn-Fazies wichtige Steuergrößen der Verjüngungsdichte. Diese Kleinstrukturen sind nicht homogen über die Hochlagen verteilt, sondern variieren in Abhängigkeit von den Standortsbedingungen. So konnten signifikante Zusammenhänge zwischen Meereshöhe, Bodenform, Exposition, Geländeneigung und der Verteilung der Konkurrenzvegetation nachgewiesen werden. Dementsprechend ändert sich auch das Verteilungsmuster der Verjüngung in Abhängigkeit von diesen Faktoren. So wächst der Anteil der mit Gräsern und Farnen bedeckten Flächen innerhalb der Probekreise mit zunehmender Meereshöhe an, parallel dazu sinkt die Verjüngungsdichte - allerdings schneller, als auf Grund der Zunahme der Konkurrenzvegetation zu erwarten wäre. Als zusätzlicher Effekt macht sich hier das in höheren Lagen geringere Wärmeangebot bemerkbar. Für die Böden gilt ähnliches; auf den Bodenformen mit der höchsten Bedeckung durch Gräser und Farne ist die Verjüngungsdichte am geringsten. Am ungünstigsten ist die Situation auf der Bodenform mittelgründiger Lehm über Rachel verfestigtem Schutt, gefolgt von Lehm, Block Humus-Böden und Nassböden. Auch die kühlen und feuchten, nach Norden und Osten gerichteten Hänge sind stärker vergrast und weisen eine geringere Verjüngungsdichte auf als Süd- und Westhänge. Natürlich spielt hier neben der Konkurrenzvegetation wie bei zunehmender Meereshöhe auch das verminderte Wärmeangebot eine wichtige Rolle für die Entwicklungsmöglichkeiten derverjüngung. Fasst man diese Ergebnisse zusammen, können die ungünstigsten Verjüngungsbedingungen in den nach Norden bis Osten exponierten Lagen oberhalb 1250 m, auf verfestigtem Lehm über Schutt lokalisiert werden. Diese Bereiche sind auf Karte 12 dargestellt. Sie befinden sich vor allem im Bereich des Rachelmassivs, aber auch um den Plattenhausenriegel. Günstiger ist die Situation im Gebiet des Moorberges und des Steinfleckberges. Dort liegen zwar auch große Bereiche über 1250 m, da hier die Bestände unmittelbar nach dem Mast jahr 1996 abgestorben sind, konnten sich jedoch mehr Pflanzen etablieren als im Rachel-Gebiet. _ zusätzlich: Hfrlenlage über 1250 m mittelgründiger Lehm über verfestigtem Schutt Exposition Nord-West bis Ost 1000 o Meter I '''. ",~~" Platten hausen- ~riegel - ~'~ ~ {'.", ' ', '-,,.,?/ "," ~f,5;,;p " Karte 12 Bereiche, in denen aufgrund der Standortvoraussetzungen ungünstige Verjüngungsbedingungen zu erwarten sind. 153
 gehören die Gebirgsnadelwälder zu den subpolaren und hochmontanen Waldformationen.")


153 6.6 StörungsregiIne hochinontaner WaldforInationen Abb. 65 Boreale Wälder, wie hier im Norden Mihnesotas, haben eine Vielzahl von Gemeinsamkeiten mit den Nadelwäldern der Gebirge. Nach Ono (1994) gehören die Gebirgsnadelwälder zu den subpolaren und hochmontanen Waldformationen. Damit sind sie eng mit den borealen Nadelwäldern verwandt, mit denen sie eine Vielzahl von Charakteristika teilen. Dazu zählen unter anderem eine ausgeprägte Saisonalität und eine kurze Vegetationszeit mit niedrigen Durchschnittstemperaturen, stark saure Böden mit geringer Nährstoffanlieferung, eine sehr langsame Streuzersetzung und damit verbunden eine mächtige Humusauflage. Sowohl die Gebirgsnadelwälder als auch die borealen Wälder sind artenarm und zeichnen sich meist durch eine üppige Bodenvegetation aus. Unterschiede ergeben sich lediglich hinsichtlich höherer Niederschläge, einer höheren Schneedecke sowie größerer Phytomasse und Baumhöhen bei den Gebirgsnadelwäldern, was allerdings auch stark von der jeweiligen Meereshöhe abhängig ist. Insofern können einige Analogien zwischen den borealen Nadelwäldern und den Fichtenhochlagenwäldern hinsichtlich walddynamischer Prozesse hergestellt werden. Nach KORPEL (1995) neigen die Urwälder der Fichtenstufe spätestens ab der Optimalphase zum Gleichschluss. Auf Grund einer geringeren Lebenserwartung und Wuchsleistung spielt die Vogelbeere in dieser Phase der Waldentwicklung keine große Rolle mehr, sie wird durch die Fichte verdrängt. Im weiteren Wachstumsverlauf reduziert sich die Kronenlänge der Fichten, was die Labilität der Bestände erhöht. Dies führt im Allgemeinen zu einem schnellen Verlauf der Zerfallsphase (KORPEL 1995). Auch SCHMln-VOGT (1985) weist auf die physikalische Instabilität borealer Wälder hin. Deshalb neigen diese einschichtigen Bestände, wenn sie sich über große Flächen erstrecken, wie dies im Untersuchungsgebiet der Fall war, auch zu großflächigen "Schadereignissen" (On et al. 1997) in Folge von Störungen. Deshalb bezeichnet die nordamerikanische Literatur die borealen Wälder auch als Disturbance forests (Ono 1994), also durch Störungen geprägte Waldökosysteme. 154


.")
154 Wichtigster Störungstyp im borealen Wald sind Brände, die sich in den kontinental geprägten Gebieten während ausgeprägter Hochdruckwetterlagen entfachen können. Die Rotationszyklen von Flächenbränden liegen im trockenen Westen Kanadas und in Alaska zwischen 50 und 200 Jahren. In Ostkanada und in den subalpinen Wäldern der nördlichen Rocky Mountains betragen sie 400 Jahre und mehr (SCHMIDT-VOGT 1985). Die Feuer in der borealen Zone haben z. T. eine gewaltige Ausdehnung, so brennen im borealen Eurasien in jedem Jahr zwei bis drei Millionen Hektar, in Extremjahren wie 1987 sogar bis zu zehn Millionen Hektar. Für die borealen Bereiche Nordamerikas beträgt der Jahresdurchschnitt ein bis fünf Millionen Hektar, in Extremjahren bis 7,4 Millionen Hektar (ANONYMUS 1995). Die Flächengrößen können bis zu 400 km', im Extremfall bis zu km' groß sein (SCHERZlNGER 1996). Dieser Störungstyp spielt jedoch in den eher maritim geprägten Fichtenwäldern West- und Mitteleuropas keine große Rolle. Es ist allerdings belegt, dass auch im Inneren Bayerischen Wald Brände auftreten können. So wird im Heimatbuch der Waldstadt Zwiesel (FRIEDEL 1954) über "zwei große Waldbrände in den Eisensteiner Hochwaldungen, die großen Schaden anrichteten" berichtet. Auch 1782 "war ein so heißer Sommer C..)' dass die dürren Wälder von selbst in Brand gerieten". Ob die Wälder tatsächlich von selbst in Brand gerieten, oder ob hier menschliche Unachtsamkeit die Ursache war, lässt sich leider nicht mehr nachprüfen. MANN (1986) berichtet, dass in Deutschland im Durchschnitt alle zwei Jahre Stürme mit Windgeschwindigkeiten von über 100 km/h, alle drei bis fünf Jahre Orkane mit Windstärke 11 und etwa alle 30Jahre gewaltige Orkane auftreten. Lawinen- und Murenabgänge können in Gebirgsnadelwäldern erhebliche Flächen betreffen. Im Bayerischen Wald und in den borealen Nadelwäldern spielen sie aufgrund der geringen Hangneigung und dem Fehlen einer alpinen Stufe keine Rolle. In Kombination mit Windwürfen, aber auch ausgelöst durch günstige Witterungsbedingungen oder durch länger anhaltende Trockenperioden spielen Insektenmassenvermehrungen eine wichtige Rolle im Störungsregime von Wäldern. Auch sie können gewaltige Ausmaße annehmen. Insbesondere nach Windwürfen führt Buchdruckerbefall rasch zu großen "Schadholzanfällen". So wurden nach den Stürmen "Vivian" und "Wiebke" 1990 alleine in Bayern 5,1 Mio. Festmeter Borkenkäferholz eingeschlagen. Auch im Nationalparkgebiet entstanden nach Sturmereignissen ha, ha' und ha Kahl- bzw. Totholzflächen in Folge von Abb.66 Feuer spielen im Entwicklungszyklus kontinental geprägter Nadelwälder eine wichtige Rolle. Auf Grund der hohen Windgeschwindigkeiten im Gebirge sind die Fichtenwälder dort stärker von Windwurf- und -bruch betroffen als die borealen Wälder. So wurde der Innere Bayerische Wald seit Beginn des letzten Jahrhunderts 80-mal von schweren Stürmen getroffen (FISCHER et al. 1990). Die größten Windwurfereignisse traten infolge der Stürme von 1868/70 mit 810 ha, 1929 mit 900 ha' und 1983/84 mit 173 ha auf. Auch ROTI- Windwurf und Käferbefall konnten nicht differenziert werden. 155
155 Abb. 67 Val/masten bei der Fichte im Bayerischen Wald seit Buchdruckerbefall. Die Analyse der Insektenschäden zwischen den Jahren 1853 und 1908 von BAR THELMESS (1972) zeigt ebenfalls, in welchem Ausmaß Insekten den Waldaufbau verändern können: Borkenkäfer 14,0 Mio. Efm, Nonne (Lymantria monacha) 10,0 Mio. Efm, Kiefernspanner (Bupalus pinarius) 2,8 Mio. Efm, Kiefernspinner (Dendrolimus pim) 2,0 Mio. Efm. In borealen Wäldern Nordamerikas treten darüber hinaus in großem Umfang Große Lärchenblattwespe (Pristiphora erichsom). "Willow leaf botchiminer" (Micrurapteryx salicifolliela) und "Large Aspen Tortrix" (Choristoneura conflictana) auf. Bei den aufgeführten Störungstypen handelt es sich um sogenannte inhärente Störungen, "da die Schlüsselorganismen des Systems an die Wirkung der Störung angepasst sind" (BOEHMER 1997) und infolgedessen das Ökosystem eine große Elastizität besitzt. So zeigt die Entwicklung auf den vom Borkenkäfer zum Absterben gebrachten Flächen, dass sowohl Vogelbeere als auch Fichte bestens an diese Prozesse angepasst sind. Die Vogelbeere, die nicht vom Buchdrucker befallen werden kann, "lässt" ihre Samen mit Hilfe von Tieren weiter verbreiten, fruktifiziert sehr früh und ist an die extremen Witterungsbedingungen auf Freiflächen angepaßt. Gleiches gilt für die Fichte, eine typische Baumart des borealen Waldes. Sie besitzt flugfähige Samen, die ihr helfen, selbst große Störungsflächen wieder zu besiedeln, so konnten LAESSIG et al. (1995) in der Schweiz bei einer Vollmast noch in 1000 m Entfernung vom nächsten Bestand zehn Fichtensamen pro Quadratmeter nachweisen. Darüber hinaus ist die Fichte in der Lage, selbst auf dicksten Rohhumusauflagen zu keimen, ist frosthart und kommt gut mit dem Freiflächenklima zu Rande. Erfolgskritisch für die Besiedlung der Flächen ist allerdings die teilweise sehr niedrige Blühfrequenz der Bäume. So berichtet BUELOW (1964) von dem seltenen Ereignis Voll- und Sprengmast, das im Bayerischen Wald nur zweimal in einem Menschenleben eintritt. In den Gebirgswäldern der Schweiz rechnet man bei der Fichte etwa alle fünf Jahre mit einer Vollmast (OTT et al. 1997). Auch wenn in den letzten Jahren ein gehäuftes Auftreten von Masten im Bayerischen Wald beobachtet werden konnte, hat die Fruktifikationshäufigkeit einen wichtigen Einfluss auf die Geschwindigkeit der Wiederbewaldung einer Störfläche. Hätten die Bäume nicht 1995 geblüht, sondern vielleicht erst 1998, wäre die Sukzession vermutlich ganz anders verlaufen. Die in den Ausgangsbeständen vorhandene Verjüngung hätte dann zwar genauso weiter wachsen können, der Eintrag von Samen auf die Flächen wäre aber sehr viel geringer gewesen, da 1998 bereits weite Teile der l I I 2000 I i Hochlagenwälder abgestorben und samentragende Altbäume in den zentralen Bereichen kaum mehr vorhanden waren. Hätten die Bäume vor Beginn der Absterbewelle geblüht, wären die meisten Keimlinge in den dunklen und kalten Altbeständen abgestorben, wie dies nach den Masten 1988 und 1992 beobachtet wurde. Auch die geringen Verjüngungszahlen im Rachelbereich können dadurch begründet werden, dass die Bestände während der Mast 1995 bzw. dem Keimen der Jungfichten im Folgejahr noch weitestgehend geschlossen waren und deshalb den Keimlingen nur ungünstige Bedingungen boten. Zusätzlich erschwert die stärkere Vergrasung der Gneisböden die Etablierung der Waldverjüngung. 156



156 allem ein warmes Frühjahr, da die Käfer erst zum Schwärmflug ansetzen, wenn die Lufttemperatur C überschreitet. Insofern ist es nicht erstaunlich, dass die Blüte 1995 zum "richtigen" Zeitpunkt stattfand, denn gute Käferjahre sind meist auch gute Jahre für die Blüte der Fichte. Abb. 68 Ein ausreichendes Wärmeangebot ist von großer Bedeutung für die Entwicklung der Fichtenkeimlinge. Demnach ist die Entstehung und auch das spätere Erscheinungsbild des Waldes weitgehend vom Zufall abhängig. Aber ist es wirklich nur der Zufall, der die Waldentwicklung steuert? Die Blüte der Fichten und die explosionsartige Vermehrung der Buchdrucker scheinen eng miteinander verbunden. Günstige Bedingungen für die Populationsentwicklung des Buchdruckers sind gleichzeitig auch günstige Bedingungen für die Anlage und Entwicklung der Fichtenblüten. Wenn die Nährstoffdepots der Bäume in Folge von warmen Jahren mit ausreichender Wasserversorgung (ROHM EDER 1972) aufgefüllt sind, wird die Blüte der Fichte durch hohe Temperaturen und Sonneneinstrahlung Ende Juni, Anfang Juli, dem Zeitfenster, in dem die weiblichen Blüten angelegt werden, stimuliert. Im darauffolgenden Jahr blühen die Bäume. Für die weitere Entwicklung der Zapfen ist eine trocken-warme Witterung günstig (SCHMIDT VOGT 1991), denn in kalten Sommern können trotz Vollmast keine reifen Samen ausgebildet werden (OTT et al. 1997). BEUDERT (pers. Mitteilung) konnte die Bedeutung des Witterungsverlaufes im Juni/Juli des Vorjahres für die Blüte der Fichte 1988, 1992 und 1995 im Bayerischen Wald bestätigen. Auch die Käferpopulation braucht warme, trockene Jahre, um sich zu einer Massenvermehrung aufschaukeln zu können. Von Bedeutung ist vor Ein weiteres Indiz für die wechselseitige Anpassung von Buchdrucker und Fichte liegt in der Tatsache, dass die Mannbarkeit (Beginn des Samentragens) der Fichte im Alter zwischen 30 und 40 Jahren im Freistand und zwischen 50 und 60 Jahren im Bestand erreicht wird (ROHMEDER 1972) und gleichzeitig auch der Buchdrucker bis zu 70 Jahre alte Bestände nur bei sehr hohen Populationsdichten und Bestände jünger als 50 Jahre (fast) gar nicht befällt (BECKER und SCHROETER 2000). Durch diese "Abstimmung" ist es möglich, dass sich die Fichte ansamt, bevor sie durch den Buchdrucker zum Absterben gebracht werden kann. Eine fortwährende koevolutive Interaktion zwischen den gegnerischen Arten Buchdrucker und Fichte hat somit eine komplexe Assoziation zwischen beiden Arten geschaffen, die deren Fortbestand sichert (COCKBURN 1995). Abb.69 Blühende Fichte. 157


157 6.7 Was bleibt VOIn alten Wald? Mit Hilfe der Interpretation von Luftbildern und unter Berücksichtigung terrestrischer Erhebungen konnte festgestellt werden, dass praktisch alle Fichtenaltbestände im Untersuchungsgebiet (1.174 ha) abgestorben sind. Die Fichte tritt nur noch bis zu einem Alter von 55 Jahren bestandsbildend auf und selbst in diesen jungen Beständen hat der enorme Befallsdruck aus den benachbarten Altbeständen im Übergangsbereich zu Absterbeprozessen geführt. Darüber hinaus haben die meisten Laubbäume die Käferentwicklung überlebt. Geringe Abgänge gibt es bei ihnen nur durch Windwurf und mechanische Schäden nach dem Absterben und Umbrechen der benachbarten Fichten. Alte Fichten haben nur als Einzelbäume und in wenigen Fällen als Baumtrupps überlebt. Die Gründe für das Überleben dieser Bäume sind nicht genau bekannt. Jedoch reproduzieren Testläufe eines Simulationsmodells zur Buchdruckerdispersion ebenfalls dieses Muster, was vermuten lässt, dass stochastische Prozesse eine entscheidende Rolle beim Überleben der alten Fichten spielen (FAHSE und HEURICH in Vorbereitung). Allerdings könnte auch individuenspezifischen Eigenschaften eine Bedeutung zukommen. Morphologisch lassen sich zwar keine Unterschiede zwischen befallenen und nicht befallenen Bäumen erkennen, möglicherweise können aber genetische Analysen hier zu neuen Kenntnissen führen. Die in den borealen Wäldern häufig auftretenden Brände haben sehr ähnliche Muster zur Folge, wie der Buchdruckerfraß im Bayerischen Wald. Sie verursachen ebenfalls ein flächenhaftes Absterben der Altbestände. Ähnlich wie nach Buchdruckerbefall bleiben auch nach Bränden Reste des alten Waldes erhalten. Deren Anteil beträgt zwischen 3 und 15 % der gesamten Brandfläche. Je größer die Brandfläche, desto größer ist auch der Anteil dieser Bestandesreste. Ganz im Gegensatz zur Buchdruckerkalamität bleiben nach Bränden auch Altbestände erhalten, vor allem in der Nähe von Gewässern. Die Fläche dieser Bestandesreste ist immer kleiner als 10 ha, 50 % sind sogar kleiner als 2 ha. Der größte Bestand, der den Buchdruckerbefall überdauerte, hat eine Fläche von 14,3 ha, die durchschnittliche Flächengröße liegt allerdings nur bei 0,9 ha, 50 % der Flächen sind sogar kleiner als 0,3 ha. Auch Einzelbäume überleben Brände. Ihre Anzahl beträgt jedoch im Durchschnitt nur 0,75 je Hektar und ist damit etwas geringer als der im Untersuchungsgebiet beobachtete Wert von fast zwei Individuen je Hektar, die den Buchdruckerbefall überleben (DE LONG und TANNER (1996), EBERHART und WOOD WARD (1987) zit. in DELONG und KESSLER (1999)). Abb.70 Bei Großbränden, wie hier im Yello wstone Nationalpark, können auch größere Nadelbaumbestände über leben, diese befinden sich dann meist in der Nähe von Gewässern. 158


158 Nur sehr heftige Stürme wie Orkane oder Hurrikans führen zur großflächigen Zerstörung der Bestände. "Gewöhnliche" Stürme reißen nur Lücken oder Schneisen in die Walddecke, meist sind nur Altbestände auf exponierten oder instabilen Standorten betroffen. Die durchschnittliche Flächengröße der Windwürfe in Folge der Gewitterstürme von 1983/84 im Nationalparkgebiet lag bei ca. 0,5 ha, die größte Fläche hatte eine Ausdehnung von 5,3 ha. Im Vergleich dazu betrug die durchschnittliche Schadensfläche der Stürme Vivian und Wiebke 1990 in der Schweiz 1 ha und in Bayern 1,7 ha, die größte Fläche in der Schweiz maß 125 ha (KOENIG et al. 1995, HOLENSTEIN 1994). Bei Windwurfereignissen sind also nicht die Bestandesreste die Inseln, sondern die Windwurfflächen selbst. Die überlebenden Bestandesreste spielen eine wichtige Rolle als Keimzellen für die Wiederbesiedlung der Störungsflächen. Auf Grund dieser Bedeutung spricht man in diesem Zusammenhang vom "biologischen Erbe" (FRANKLIN 1994), welches in den Bestandesresten erhalten wird. Welche Rolle die den Buchdruckerbefall überlebenden Bestände für die Tierwelt haben, ist noch weitgehend unbekannt. Zufällige Beobachtungen deuten jedoch darauf hin, dass sich in den Bereichen, in denen Bestandesreste überlebt haben, mehr Waldameisenkolonien und auch mehr Hinweise auf die Anwesenheit von Auerhühnern (Tetrao urogallus) gefunden wurden. Für die Waldverjüngung kann die Bedeutung der überlebenden Bestände und Einzelbäume mit Hilfe einer GIS-Analyse unter den folgenden Annahmen abgeschätzt werden: Nach BRANG (in BAYERI SCHE LANDESANSTALT FÜR WALD UND FORSTWIRTSCHAFT 1998) erfolgt ein nennenswerter Eintrag von Fichtensamen nur bis in etwa 100 m vom Bestandesrand. Auch KOHLERMANN (zit. in ROHMEDER 1972) kommt nach Untersuchungen mit Samenfangkästen zu dem Ergebnis, dass bis in doppelter Baumlänge noch 50 % und in vierfacher Baumlänge noch 10 % der Samendichte des Bestandesinneren erreicht wird. Das entspricht nach SIEGL und MESSER (beide zit. in ROHMEDER 1972) ca bzw. 200 Samen je Quadratmeter. Da für die Samenverbreitung ausgehend von EinzeIbäumen keine Quellen vorliegen, wird unterstellt, dass hier eine ausreichende Samendichte nur bis in 50 m Entfernung erreicht wird. Legt man nun um jeden im Untersuchungsgebiet noch vorhandenen Bestand eine Zone von 100 m und um jeden noch vorhandenen Einzelbaum eine Zone von 50 m, ergibt sich eine Flächenabdeckung von fast 66 0/0. Allerdings sind die Restbestände und die Einzelbäume nicht gleichmäßig über das Unter- Abb. 71 Je heterogener die Waldstrukturen sind, desto größer ist die Wahrscheinlichkeit, dass alte Fichten Buchdruckermassenvermehrung überleben, die Freiflächen im Hintergrund entstanden durch Borkenkäferbekämpfung. 159
 Teil der Fichtensamen auch viel weiter als 100 m.")

.")


159 suchungsgebiet verteilt, so dass einige Bereiche nicht ohne weiteres von Samen erreicht werden können. Ein Beispiel dafür sind die höheren Lagen des Moorberges. Dort sind auf großer Fläche keine alten Fichten mehr vorhanden. Allerdings fliegt ein (kleiner) Teil der Fichtensamen auch viel weiter als 100 m. Insbesondere die Samen, die im Spätwinter auf die Schneedecke fallen, werden bei verharschter Oberfläche oft sehr weit vom Wind verblasen. Ob diese Art der Samenverbreitung einen nennenswerten Beitrag zur Verjüngung der Bestände leisten kann, wird zur Zeit mit Samenfallen untersucht. Darüber hinaus ist zu erwarten, dass die Einzelbäume und die Randbäume der verbleibenden Bestände häufiger fruktifizieren, wie das auch von Waldrändern und freistehenden Fichten bekannt ist (ROHMEDER 1972). Schwer abzuschätzen ist allerdings, ob und wenn ja in welchem Umfang sich der Buchdruckerbefall in den übriggebliebenen Fichtenbeständen fortsetzen und wie hoch die Überlebensrate der verbleibenden Einzelbäume sein wird. Der Eintrag von Vogelbeersamen wird zunächst in der gleichen Größenordnung ablaufen wie bisher. Die Altbäume dürften in den nächsten Jahren zwar durch das Zusammenbrechen der abgestorbenen Fichten beschädigt werden, da die Vogelbeeren jedoch schon nach wenigen Jahren fruktifizieren, dürfte bei dieser Baumart in Zukunft mit einer kontinuierlich anwachsenden Samen nachlieferung zu rechnen sein. Von einem Abreißen der Verjüngung nach dem Absterben der Altbestände, wie es GEITNER (2000) formuliert hat, kann also nicht die Rede sein. ~r,,".... :... ' ;'" I. ' '. '1/.., Karte 73 Bereiche der Hochlagen, die auch nach dem Absterben der meisten Altbestände noch ausreichend von Fichtensamen erreicht werden können.. ;~ : "..... \. '::'. :..... Überltberde Einze!bäume.. Überltberde Bestände S) mzone um die Einzelbäurre m Zone um die Bestände 1000 o Meter... :. :. ':0 '.J. ~.t)o. ~ 160
")


160 161 (Abb. 72)
 Die großen Totholzmengen in Kombination mit dem reichlichen Blütenangebot bilden günstige Lebensbedingungen für xylobionte Insekten, Die durch den Buchdrucker ausgelöste massive Veränderung der")

. Auf den entstandenen Freiflächen bildet sich ein für die Hochlagen reiches Angebot an Blütenpflanzen, aber auch eine sich immer weiter ausbreitende Grasdecke aus.")
.")
161 6.8 Wie reagieren die Tiere? Abb, 73 (u.) und 74 (r.) Die großen Totholzmengen in Kombination mit dem reichlichen Blütenangebot bilden günstige Lebensbedingungen für xylobionte Insekten, Die durch den Buchdrucker ausgelöste massive Veränderung der Waldvegetation führt natürlich auch zu einem Wandel des Lebensraumangebotes vieler Tierarten. Wichtige Charakteristika des neuen Lebensraumes sind sein Totholzreichtum, das Fehlen der alten Bäume und damit verbunden eine größere Biomasseproduktion auf dem Waldboden. In Folge verändert sich das Mikroklima auf der Fläche: die Temperaturen, im Besonderen die Bodentemperaturen steigen; es erhöhen sich Niederschlag und Einstrahlung, die relative Luftfeuchtigkeit sinkt (BRUNNSCHWEIGER 2000). Auf den entstandenen Freiflächen bildet sich ein für die Hochlagen reiches Angebot an Blütenpflanzen, aber auch eine sich immer weiter ausbreitende Grasdecke aus. Die zahlreich auf den Freiflächen wachsenden Vogelbeeren haben eine große Bedeutung für die Tierwelt. Werden ihre Beeren doch von bis zu 63 verschiedenen Vogelarten als Nahrung genutzt (TUREK 1961). Darüber hinaus verwerten 31 Säugetierarten die verschiedensten Teile dieser Baumart (TUREK 1967). Diese Faktorenkombination führt zu einem nahezu idealen Biotop für xylobionte Insekten wie Wildbienen, aculeate Wespen (Aculeata), Bockkäfer (Cerambycidae) und Schwebfliegen (Syrphidae) (BARTAK 1995, KOEHLER 1997, KUHL MANN 1999). Gleichzeitig verbessern sich auch die Lebensbedingungen für Reh und Rothirsch. Bei diesen Arten wird ein starker Anstieg der Sommerbestände erwartet. Auch die Kleinsäuger profitieren durch das Absterben der Altbestände und reagieren durch er 'höhte Reproduktion und ein Anwachsen der Populationen. Bei einer beispielhaften Erfassung konnte in noch geschlossenen Beständen eine Individuendichte von 26 Tieren je Hektar, im gleichen Jahr in abgestorbenen Beständen im Durchschnitt 70 Individuen je Hektar nachgewiesen werden. Auffällig war jedoch, dass es im Gegensatz zu Windwurfflächen nicht zum Zusammenbruch der ursprünglichen Kleinsäugerpopulation kam, sondern die Dominanz der Waldarten erhal-. ten blieb. Die Flächen wurden von Rötelmaus (Clethrionomys glareolus), Gelbhalsmaus (Apodemus flavicollis) und Waldspitzmaus (Sorex ara- 162 ~--~
, Siebenschläfer (Glis glis) und Zwergspitzmaus (Sorex minutus). Abb.")

162 neus) dominiert. In vergrasten Partien konnte im Gegensatz zum geschlossenen Wald auch die Erdmaus (Microtus agrestis) nachgewiesen werden (HUBER 2000). Das Vorkommen weiterer Arten beschränkte sich auf Einzelexemplare von Haselmaus (Muscardinus avellanarius), Siebenschläfer (Glis glis) und Zwergspitzmaus (Sorex minutus). Abb.75 Unter den Spechtarten ist der Buntspecht die einzige, die nachhaltig von den Lebensraumveränderungen profitieren konnte. Im Gegensatz dazu wirkt sich das großflächige Absterben der Altbäume eher negativ auf die vorkommenden Ameisenarten aus. So konnte im Rahmen der Inventur ein erheblicher Rückgang der Nesterzahl und Neststandorte der Waldameisen nachgewiesen werden, wofür im Wesentlichen zwei Gründe verantwortlich sind: Ein bedeutender Nahrungsbestandteil der Waldameisen sind Blattläuse (Aphidina). die vor allem in den Fichtenkronen leben. WELLENSTEIN (2000) gibt an, dass die Nahrung der Waldameisen zu 62 % aus Honigtau von Blattläusen besteht. Nach dem Absterben der Fichten fällt dieser Nahrungsbestandteil fast vollständig aus. Nur wenige Läuse, die auf Gräsern und krautigen Pflanzen leben, verbleiben dann als Honigtau- Produzenten. Die durch die zunehmende Belichtung ausgelöste Ausbreitung von Gräsern gefährdet die Neststandorte der Ameisen. Werden sie überwuchert, sind die Ameisen nicht mehr in der Lage, das spezifische Mikroklima unter der Nestkuppel zu erhalten und müssen das Nest verlassen. Zusätzlich beeinträchtigt ein dichter Pflanzenwuchs in unmittelbarer Nestumgebung das Auslaufverhalten der Arbeiterinnen (TRAVAN und SCHMIDT 1994). Auch BRUNNSCHWEIGER (2000) konnte eine Schwächung der Kolonien auf Borkenkäferflächen beobachten, was sich in einem verminderten Ein- und Auslaufverhalten vermutlich in Folge der hohen Einstrahlung dokumentierte. Insgesamt lag die Zahl der Ameisenarten auf abgestorbenen Flächen (5) weit unter der Artenzahl von intaktem Wald (9). Auch die Anzahl der gefundenen Nester war mit fünf auf den Borkenkäferflächen weit geringer als auf den noch lebenden Beständen, wo 16 Ameisennester auf m' großen Probeflächen nachgewiesen werden konnten. Im Gegensatz dazu konnte in der Hanglage weder eine deutliche Artenverarmung noch ein Rückgang der Nestanzahl beobachtet werden (BRUNNSCHWEIGER 2000). Sehr gute Informationen liegen über die Reaktionen der Vogelwelt auf die Lebensraumveränderungen vor. Allerdings stammen die Beobachtungen von einer Dauerbeobachtungsfläche am Großen Spitzberg, die zum überwiegenden Teil der oberen Hanglage zuzurechnen ist und durch einen großen Windwurf dominiert wird. Seit dem Beginn der Untersuchungen 1989 hat sich die Gesamtartenzahl von 48 auf 73 erhöht. Ganz im Gegensatz zum abrupten Absterben der WaIdbestände verlief diese Entwicklung bei den Vögeln eher kontinuierlich. Die Zunahme der Arten erfolgte in erster Linie bei den Singvögeln und hier vor allem bei den Zugvögeln. Auch die Siedlungsdichte der Vögel nahm im Laufe der Jahre zu, nur unterbrochen von Bestandseinbrüchen unmittelbar nach dem Absterben der Altbestände, was darauf zurückgeführt wird, dass die Totholzbestände ohne Bodenvegetation für viele Vogel- 163
 und Goldhähnchen (Regulus regulus), Tannen-")
, Kohlmeise (Parus major). Grasmücken (Sylvia spec.), Laubsänger (Phylloseopus spec.")
163 arten sehr unattraktiv sind. Auch hier sind es die Zugvögel, die vor allem zu diesem Anstieg beigetragen haben. Im Gegensatz dazu nahm die Abundanz der Standvögel ab (SCHERZlNGER 1999). Unter den Verlierern der Lebensraumveränderungen befinden sich die Arten des Kronenraumes und Samenfresser, wie Fichtenkreuzschnabel (Loxia curvirostra) und Goldhähnchen (Regulus regulus), Tannen- (Parus ater) und Haubenmeise (Parus cristatus), sowie Erlenzeisig (Spinus spinus) und Gimpel (Pyrrhula phyrrula). Gewinner sind die Arten der Parklandschaft und Waldsteppe wie Heckenbraunelle (PruneIla modularis), Kohlmeise (Parus major). Grasmücken (Sylvia spec.), Laubsänger (Phylloseopus spec.), Amsel (Turdus merula) und Gartenrotschwanz (Phoenicurus phoenicurus) (SCHERZlNGER 1999). Die Dichte von Dreizehenspecht (Picoides tridactylus) und Schwarzspecht (Dryocopus martius) stieg während des Buchdruckerbefalls stark an, konnte sich in Folge jedoch trotz des hohen Totholzangebotes nicht auf diesem hohen Niveau halten und sank wieder bis auf die Ausgangswerte ab. Nur der Buntspecht (Dendrocopos major) profitierte nachhaltig von der Lebensraumumgestaltung und konnte seine Siedlungsdichte gegenüber dem Referenzjahr 1982 um den Faktor 7 erhöhen (SCHERZlNGER 1998). Abb.76 Die Heckenbraunelle gehört, wie auch andere Arten der Parklandschaften zu den Gewinnern der Entwicklung nach dem Borkenkäferbefall. Ganz anders stellt sich die Situation im Winter dar, denn dann macht sich der Verlust an Wetterschutz, an Insektenbeute und Baumsamen stark bemerkbar: Im Gegensatz zum Sommerhalbjahr sinkt die Artenzahl im Winter während des ganzen Beobachtungszeitraumes kontinuierlich ab. Auch die Siedlungsdichten sinken bei allen Arten ab, im Winter gibt es keine Gewinner. Insbesondere für die Standvögel ergeben sich aus der in Sommer und Winter stark unterschiedlichen Lebensraumqualität Probleme. Während im Sommer der Tisch reichlich gedeckt ist, gibt es im Winter weder Deckung noch Nahrung. Als Beispiel für eine solche Art kann das Auerhuhn dienen. Während des Sommers sind die bodennahen Temperaturen auf den Freiflächen höher als in den kühlschattigen Wäldern, das reiche Blütenangebot und die stärker fruchtende Heidelbeere tragen zu günstigeren Bedingungen bei der Kükenaufzucht bei (SCHERZlNGER 1995). Die Ausbreitung der Gräser und der Rückgang der Waldameisen wirken diesem Trend entgegen. Im Winter müssen sich die Auerhühnervermutlich aus den abgestorbenen Bereichen zurückziehen, da sie keine Deckung vor der Witterung und vor Fressfeinden (z. B. Habicht) finden können. SCHERZlNGER (1995) unterscheidet bei den waldbewohnenden Tierarten drei Strategietypen. Da sind zum einen die hochspezialisierten Arten des Waldinnenklimas, klassische k-strategen, die an das konstante Lebensraummilieu alter Wälder angepasst sind. Zur zweiten Gruppe gehören die Arten der Störungsflächen, meist hochmobile Generalisten, die schnell auf Änderungen des Lebensraumes reagieren können, also die klassischen r-strategen. Bleiben als dritte Gruppe noch die Arten, deren Habitatansprüche die ganze Pa- 164

 und das Eichhörnchen (Sciurus vulgaris), die in den vergangenen Jahren im")
164 lette der unterschiedlichen WaIdentwicklungsstadien, also auch Störungsflächen umfassen. Als Gewinner nach der großflächigen Borkenkäferkalamität gehen vor allem die r-strategen wie die Mäuse, viele Insektenarten und Laubsängerarten hervor. Die großen Verlierer sind die k-strategen des "Klimaxwaldes", ihr Lebensraum ging großräumig verloren. Zu diesen Verlierern gehören allen voran der Weißrückenspecht (Dendrocopos leucotosl, aber auch Baummarder (Martes martes) und das Eichhörnchen (Sciurus vulgaris), die in den vergangenen Jahren im Hochlagenwald immer weniger beobachtet werden konnten. Aber auch die Arten des Waldlückensystems haben große Schwierigkeiten, die Situation zu meistern, wie am Beispiel des Auerhuhns und der Waldameisen deutlich wird. Abb.77 Die stark unterschiedliche Habitatqualität im Sommer und Winter erschwert dem Auerhuhn eine kontinuierliche Besiedelung der abgestorbenen Flächen. 165
165 6.9 Verändern sich die Störungsrnuster? Wie in den vorhergehenden Kapiteln herausgearbeitet sind Borkenkäferfraß und Sturmwürfe ein immanenter Bestandteil der Fichtenwaidökosysteme, die in mehr oder weniger regelmäßigen Zeitintervallen wiederkehren. Allerdings deutet Einiges darauf hin, dass sich die Störungsmuster in den letzten Jahren verändern. So wird aus vielen Regionen von einer Zunahme der Frequenz und Intensität von Störungen berichtet. Beispielsweise nahm die Fläche abgebrannter Wälder in den vergangenen beiden Dekaden sowohl in Kanada als auch in Rußland um J0 zu (AUCLAIR und CARTER 1993, KRANKI NA 1991). Auch die Frequenz von heftigen Stürmen stieg in Kanada im gleichen Zeitraum -ebenfalls um % an (ETKIN 1993 zit. in JARDINE 1995). Gleichzeitig erhöhte sich die Windgeschwindigkeit in den borealen Wäldern Ontarios um 50 % (schindler 1990 zit. in JARDINE 1995). Auch in Mitteleuropa scheinen sich die Intervalle, in denen heftige Stürme auftreten, verkürzt zu -haben. Zudem hat sich die Heftigkeit der Stürme erhöht (GRASSL und KLINGHOLZ 1990). Darauf deutet auch die kurze Abfolge und die Heftigkeit der Orkane "Vivian" und "Wiebke" 1990 sowie des Orkans "Lothar" im Jahr 2000 hin. Insbesondere "Lothar" erreichte eine bisher in Mitteleuropa nicht gekannte Wucht: Bei einer Windgeschwindigkeit von 212 km/h fiel das Windmessgerät am Feldberg aus. Insgesamt waren in Baden-Württemberg ca ha betroffen, ha mehr als bei den "Jahrhundertstürmen" Anfang der 90er Jahre. Für ganz Mitteleuropa wird ein Sturmholzanfall von 180 Mio. fm/ha angegeben (PRESSEMITIEILUNG DES MINISTERIUMS LÄNDLICHER RAUM 2001). Für wieder andere Teile der Erde wurden solche Beobachtungen nicht bestätigt, beispielsweise konnten LAESSIG und MOCALOV (2000) im Ural für die Periode keine Zunahme der Windgeschwindigkeit und Sturmhäufigkeit feststellen. Auch Massenvermehrungen von Insekten scheinen in den letzten Jahren ebenfalls an Heftigkeit zugenommen zu haben. So wird aus Alaska von der größten Massenvermehrung des "spruce beetles" (Dendroctonus rufipennis) in historischer Zeit berichtet: Bis 1996 starben in südwestalaska km' Wald in Folge Käferbefalls ab. Im Ber~ich des Wrangel Mountains st. Elias Nationalparks in südostalaska waren es km'. Der Höhepunkt der Entwicklung konnte, wie auch im Bayerischen Wald, 1996 beobachtet werden. Damals starben in nur einem Jahr km' Fichtenwälder ab (zum Vergleich beträgt die Fläche des Nationalparks Bayerischer Wald 250 km 2 ). Seitdem war ein stetiger Rückgang des Befalls zu beobachten, 2000 lagen die Neubefallsflächen "nur" noch bei 351 km'. Zwischen 1994 und 2000 summierte sich die betroffene Fläche auf km'. Nimmt man die übrigen in Alaska auftretenden Schadinsekten noch hinzu, beläuft sich die gesamte Schadensfläche zwischen 1994 und 2000 auf km' (USDA FORESTsERVICE 2001). Ein weiterer in Nordamerika gefürchteter Schädling ist der "spruce Budworm" (Choristoneura occidentalis). Es handelt sich dabei um eine Motte, deren Raupen an Tannen und Fichten fressen. Die Massenvermehrungen dieses Insekts zeigten im vergangenen Jahrhundert einen exponentiellen Trend, d. h. das Insekt verdoppelte mit jeder Massenvermehrung die betroffene Fläche waren es 10 Mio. Hektar, Mio. Hektar und Mio. Hektar, was in etwa der Fläche Frankreichs entspricht (BLAIS 1984 zit. in JÄRDINE 1995). Auch die aktuelle Buchdruckermassenvermehrung im Bayerisch-Böhmischen Grenzgebirge hat mittlerweile erhebliche Dimensionen angenommen. Mittlerweile sind im Rachel-Lusen-Gebiet über ha Fichtenaltbestände betroffen. Im angrenzenden Nationalpark sumava dürfte das 166

.")

166 Ausmaß abgestorbener und geräumter Flächen eine ähnliche Größenordnung erreicht haben. Damit liegt die bisher erreichte Fläche aber noch unter dem Wert, der für die Massenvermehrung angegeben wird. Damals waren im Bayerisch-Böhmischen Grenzgebirge rund ha Kahlflächen entstanden (SCHERZlNGER 1996). Betrachtet man nur das alte Nationalparkgebiet, liegt die Fläche abgestorbener Fichtenaltbestände jedoch weit über dem für angegebenen Wert von ha (Windwurf und Käferbefall). Grund für ein sich änderndes Störungsregime könnte die sich abzeichnende globale Klimaveränderung sein. So prognostizieren Klimamodelle unter anderem eine größere Sturmfrequenz und -heftigkeit, die Zunahme von Trockenperioden und den Anstieg der Durchschnittstemperaturen. Die Folge wären häufigere Windwurfereignisse, größere Windwurfflächen, die Zunahme von Brandfrequenz und deren durchschnittliche Größe sowie häufigere und länger anhaltende Insekten kalamitäten, da die Aktivitäten von Insekten entscheidend durch den Witterungsverlauf beeinflusst werden (Ipcc 2001, BAYFORKLIM 1999, HEURICH eta!. 2001). Einige Indizien deuten darauf hin, dass sich das Klima bereits verändert; so hat sich die globale Durchschnittstemperatur im 20. Jahrhundert um 0,6. Cerhöht (lpcc 2001). Die sieben heißesten Jahre seit Beginn der Wetteraufzeichnungen lagen alle in den 80er und 90er Jahren (ANONYMUS 1993). Auch die Aufzeichnungen der Wetterstationen des Nationalparks bestätigen diesen Trend (JEHL 2001). Abb.78 Stimmen die Prognosen der Klimaforscher, ist in Zukunft häufiger mit Windwürfen zu rechnen. 167
167 7. Schlussfolgerungen für das Nationalpark-Management Die Bewältigung der Borkenkäfermassenvermehrung stellt die Nationalparkverwaltung in mehrerlei Hinsicht vor große Herausforderungen. Insbesondere die verschiedenen, zum Teil konträr verlaufenden Zielvorgaben sind schwierig zu vereinen. Da ist nicht nur der klassische Konflikt zwischen der Erschließung des Gebietes mit Rad-, Wander- und Reitwegen, Loipen, Erlebnissteigen und Informationszentren für Erholung und Bildung und den Naturschutzzielen, nein - bereits innerhalb der Naturschutzziele selbst zeigt sich das Dilemma, in dem sich das Nationalparkmanagement grundsätzlich befindet. In einem Nationalpark soll sich die Natur nach ihren eigenen Gesetzmäßigkeiten entwickeln dürfen, ohne dass der Mensch bzw. die Nationalparkverwaltung immer wieder steuernd eingreift. Mit der Formel "Natur Natur sein lassen" wurde diese Zielstellung des Naturschutzes in Nationalparken treffend charakterisiert. Auf der anderen Seite stehen die Forderungen zum Erhalt der Biodiversität, also der Artenschutz. Obwohl von der Internationalen Naturschutzunion beide Ziele mehr oder weniger auf die gleiche Hierarchieebene gestellt werden, kann der Artenschutz in Nationalparken nur eine untergeordnete Rolle spielen, die sich vor allem auf die Vervollständigung der ursprünglichen Artenausstattung durch Wiederansiedlungsprojekte beschränkt. Begründet wird dies vor allem durch die Vermutung, dass der Artenschutz praktisch im Kielwasser des Prozessschutzes mit erfüllit werden kann. Dies setzt aber voraus, dass ein Schutzgebiet alle verschiedenen Entwicklungszyklen der zu schützenden Lebensgemeinschaften in geeigneten Proportionen unter Berücksichtigung des natürlichen Störungsregimes umfasst. Wie soll man aber auf einen großflächigen, nahezu kompletten Verlust der Altholzausstattung und der daran gebunden Arten in einer ganzen Waldgesellschaft reagieren, wenn die Schutzgebiete - wie oft in Mitteleuropa - relativ klein sind? Soll man die Natur sich selbst überlassen und damit Artenverluste in Kauf nehmen oder versuchen, repräsentative Teile der Hochlagen durch Borkenkäferbekämpfung mit all ihren negativen Begleiterscheinungen zu schützen? Wie viel Eingriff darf zum Schutz einer gefährdeten oder gar vom Aussterben bedrohten Art sein? Die Entscheidung darf aber nicht auf die Frage hinauslaufen, was mehr wert ist, der Erhalt von Arten oder der "Schutz der natürlichen Prozesse". Denn beides ist nur scheinbar getrennt, da die Qualität der Prozesse von der Artenvielfalt abhängt. Darüber hinaus spielen wirtschaftliche und politische Zielsetzungen eine wichtige Rolle für die Entscheidungsfindung. Zum einen gilt es, die an den Nationalpark angrenzenden Wirtschaftswälder vor dem Übergreifen des Borkenkäfers zu schützen. Ein klares Ziel, das durch die Anlage eines mindestens 500 m breiten Randbereiches, in dem vom Buchdrucker befallene Bäume entnommen werden, erreicht werden konnte (HEURICH et al. 2001). Zum anderen wurde in der Nationalparkverordnung festgelegt, dass "in einem Zeitraum bis zum Jahr 2017 (...) die Ausbreitung des Borkenkäfers auf die Wälder der Hochlagen zwischen Falkenstein und Rachel zu verhindern" ist. Was zunächst ein Kompromiss zwischen Nationalparkgegnern und -befürwortern ist, ist auf der anderen Seite auch eine Lösung des Dilemmas zwischen Prozess- und Artenschutz. Während im alten Nationalparkgebiet die nicht gelenkte Entwicklung bis auf die Einschränkung durch den Randbereich ungehindert ablaufen kann, versucht man im Erweiterungsgebiet, die alten Bergfichtenwälder mitsamt ihrer Arteriausstattung zu erhalten, was bisher sehr erfolgreich gelingt. Dieses Zonierungskonzept ist mittelfristig auch die Voraussetzung dafür, dass die Arten des Waldinnenklimas und Waldlückensystems das Rachel Lusen-Gebiet wieder besiedeln können. 168


168 Eine weitere wichtige Zielvorgabe für das Parkmanagement ist auch 14(2) der Nationalparkverordnung : "Soweit die natürliche WaIderneuerung flächig und längerfristig ausbleibt, soll die Entwicklung einer standortgerechten, natürlichen Waldzusammensetzung unterstützt werden". Auf Grund der deutlichen Zunahme der Verjüngungsdichten seit 1998 auf jetzt Pflanzen je Hektar hat sich die Situation deutlich entspannt. Insbesondere wenn man bedenkt, dass sich in der Höhenschicht zwischen cm nochmals Pflanzen je Hektar befinden, die wohl zu einem erheblichen Teil bei der nächsten Inventur die 20 cm-schwelle erreichen werden. Damit haben die Durchschnittswerte schon ein Niveau erreicht, wie es für Wirtschaftswälder im Flachland empfohlen wird. So nennen BURSCHEL und Huss (1997) hier Richtwerte von bis Fichten je Hektar. Die Soll-Werte für Hochlagenwald, die On et al. (1997) mit bis Pflanzen angeben, sind damit sogar schon überschritten. MAYER und On (1991) halten sogar nur 200 Verjüngungsansätze je Hektar für ausreichend, die Strukturnachhaltigkeit von Fichtenbeständen als gesichert ansehen zu können. Auch im Vergleich zu mitteleuropäischen Urwäldern der Fichtenstufe sind die im Jahr 2000 erreichten Verjüngungsdichten relativ hoch. So gibt KORPEL (1997) für die Urwälder der Westkarpaten im Durchschnitt Pflanzen an und auch LEIBUNDGUT (1978) berichtet, dass in den Urwäldern der Bergstufe selten mehr als Pflanzen je Hektar gefunden werden. Allerdings ist die Verjüngung sehr ungleichmäßig über die Hochlagen verteilt, so dass es auch noch einige Bereiche mit relativ wenigen Verjüngungspflanzen gibt. Ein solcher Bereich ist das Rachelmassiv. Von Bedeutung ist auch, dass der überwiegende Teil der Pflanzen noch sehr klein ist und eventuell in Zukunft auftretende "Schädlinge" wie der Schneeschimmel große Teile der Verjüngung vernichten könnten. Als gesichert können erst die Pflanzen angesehen werden, die deutlich über die durchschnittliche Schneedecke von zwei bis drei Metern hinausragen (On et al. 1997). Bis die Pflanzen diese Größe erreichen, wird noch einige Zeit vergehen, da die Verjüngungszeiträume in den Hochlagen sehr lang sind. Beispielsweise gibt KORPEL (1997) die Dauer für die Verjüngungsphase mit 90 bis 100 Jahren an, bei MAYER und On (1991) betragen sie sogar 100 bis 200 Jahre. Selbst für den Wirtschaftswald streben On et al. (1997) Verjüngungszeiträume von über 150 Jahren an. Die für Gebirgswaldökosysteme rasante Verjüngungsentwicklung der letzten Jahre legt jedoch die Vermutung nahe, dass das Verjüngungsstadium im überwiegenden Teil der Hochlagenzone des Nationalparks Bayerischer Wald wesentlich rascher abgeschlossen sein wird. Nach dem aktuellen Sachstand sind Maßnahmen zur Stützung der natürlichen Waldverjüngung nicht notwendig. Die Entwicklung des neuen Waides muss aber auch in Zukunft genau verfolgt werden, um dieses für Mitteleuropa einzigartige Ereignis möglichst umfassend wissenschaftlich dokumentieren. Allerdings ist es wichtig, sich immer wieder klar zu machen, in welchen Zeiträumen die Entwicklungsvorgänge in den Hochlagenwäldern ablaufen. Zeiträume, die unsere Geduld und unser Vorstellungsvermögen bei weitem übersteigen (On et al. 1997). Abb.79 Die aktuellen Forsch u ngsergebn isse zeichnen ein positives Bild von der Entwicklung der Waldverjüngung. Wenn sich dieser Trend fortsezt, wird es nicht notwendig sein, der Natur mit Ergänzungspflanzungen unter die Arme zu greifen. 169
169 Waldentwicklung iin Inontanen Fichtenwald nach großflächigein Buchdruckerbefall iin Nationalpark Bayerischer Wald Seit Beginn der Buchdruckermassenvermehrung im Nationalpark Bayerischer Wa~d sind in der hoch montanen Stufe des Rachel-Lusen-Gebietes fast 85 % der ca ha umfassenden Fichtenbestände abgestorben. Um deren Auswirkungen auf die natürliche Regeneration der WaIdbestände zu verfolgen, werden seit 1996 in zweijährigem Turnus Inventuren durchgeführt. Als Referenz für diese Erhebungen dienen die Ergebnisse der Waldinventur 1991, als die Fichtenwälder in den Hochlagen noch weitgehend geschlossen waren. Datenerhebung und Auswertung entsprechen dem in der Staatsforstverwaltung eingesetzten Standardverfahren für die Forsteinrichtung. Auf 572 permanenten Probeflächen, die in einem 200 mal 200 m Raster über das Gebiet verteilt sind, werden Daten über die Baumverjüngung größer 10 cm erhoben. Darüber hinaus wurden die Flächenanteile von Kleinstrukturen erfasst, die als bedeutend für die Entwicklung der WaIdverjüngung angeschätzt werden. Außerdem wurden mit Hilfe von Luftbildern in einem Teil der Hochlagen alle Bäume und Bestände kartiert, die den Buchdruckerbefall überlebt haben. Fichtenaltbestände sind dort weitestgehend verschwunden. Bestandesbildend tritt die Fichte nur noch bis zu einem Alter von ca. 55 Jahren auf und selbst diese jungen Bestände wurden von den Rändern her durch Buchdrucker befallen. Altfichten haben nur als Einzelbäume und kleine Trupps die Gradation überlebt. Trotz der geringen Dichte von ca. 2 lebenden Bäumen je Hektar können rechnerisch 66 % des Untersuchungsgebiets ausreichend mit Baumsamen versorgt werden. Die Verjüngungsdichte (> 20 cm) ist von 978 Pflanzen je Hektar im Jahr 1991 auf im Jahr 2000 angestiegen. Berücksichtigt man die Pflanzen zwischen 10 und 20 cm, die jedoch erst seit 1996 erhoben werden, findet man in den Hochlagen bereits durchschn ittlich Pflanzen je Hektar. Auch wenn ein Großteil der Waldverjüngung noch nicht als gesichert angesehen werden kann, hat die Dichte mittlerweile ein Niveau erreicht, wie es beispielsweise auch für Wirtschaftswälder im Flachland angestrebt wird. Die häufigste Baumart ist mit 77 % die Fichte, gefolgt von dervogelbeere mit 19 0 /0. Alle anderen Baumarten spielen mit zusammen 4 % nur eine untergeordnete Rolle. Die Verjüngung weist in Abhängigkeit von den Kleinstrukturen des Waldbodens eine starke Klumpung auf. Insbesondere in der Gras- und Farn-Fazies, die zusammen 56 % der Hochlagen bedecken, konnten nur wenige Pflanzen gefunden werden. Im Gegensatz dazu ist die Situation bei den Kleinstrukturen "Baumkontakt" und "Moderholz" viel günstiger. Obwohl sie nur einen Flächenanteil von 8,3 % einnehmen, wachsen hier 45 % aller erfassten Fichten. Darüber hinaus variiert die Verjüngungsdichte in Abhängigkeit von Bodenform, Meereshöhe, Exposition und dem Absterbezeitpunkt der Altbestände. Insbesondere im Bereich des Rachels ist die Verjüngungsdichte noch relativ niedrig. Dort dominiert die Bodenform mittelgründiger Lehm über verfestigtem Schutt, große Teile befinden sich über m, sind stark vergrast und starben erst einige Jahre nach der Fichtenvollmast 1995 ab. Schäden an den Verjüngungspflanzen traten bisher nur in sehr geringem Umfang auf. Von Bedeutung ist nur der Leittriebverbiss durch Schalenwild bei der Vogelbeere. Er betrug im Jahr ,7%,imJahr2000 nurnoch 14,4%. Durch Vergleich mit Forschungsergebnissen aus anderen hochmontanen Waldformationen wird der Schluss gezogen, dass Störungen ein natürlicher Impuls für die Entwicklung dieser Waidökosysteme sind, an den die Biozönose angepasst ist. 170
170 Forest deveiopinent after Iarge bark beetle outbreak in the Bavarian Forest National Park Over 85 % of the 2300 ha mountain spruce forest at high elevations have been killed since the beginning of the bark beetle outbreak in the Bavarian Forest National Park. Following the outbreak an inventory to monitor the pracesses of natural regeneration has taken place every other year since Comparative data was taken fram the 1991 forest inventory data. This latter data was taken at a time when the canopy of the mountain spruce forest was mostly closed. All data gathering and analysis was completed using Bavarian State Forest Service standardised pracedures. Data fram all trees between 5 and 10 cm in height was gathered in 572, 200 metre plots. In addition, the area percentage of small site factors considered important for the development of forest regeneration were documented, and aerial photographs were used to identify and map all spruce trees that survived the bark beetle outbreak in one part of the research area. The results showed that old spruce stands had almost disappeared fram the research area, with the remaining spruce stands consisting of young trees up to a maximum age of 55 years. However, many parts of the young stands, and in particular those adjacent the stand border, were also killed by bark beetle. The only old spruce trees to survive the outbreak stood as single trees or in small clumps. Although the number of living trees remaining is relatively small, theoretical calculations indicate that their seeds could reach 66% of the study area. In 1991 only an average of 978 trees greater then 20 cm in height were found per hectare. This contrast with a count of 1928 trees greater then 20 cm in height in the year As such, regeneration density has increased significantly since If trees between 10 and 20 cm in height are also included in the count (trees of this height have been included in the survey count since 1996) then the total number of plants is 3569 per hectare. Even if the greater part of the regeneration has not reached a level were it could be considered successful, the regeneration density has reached a level comparable with targets for tree density in lowland managed forests. Spruce and Mountain Ash account for 77% and 19% respectively of all the young plants. Other tree species are of minor significance accounting for only 4% ofthe area in total. The regeneration is spread unevenly thraugh out the area. Clumps of regeneration are mainly influenced by small structure site factors. In particular, few trees are found in areas covered with grass and fern, wh ich account for 56 % of the forest floor area. This contrasts with those areas where small structures such as "stem contact" and "decomposing wood" occur in abundance. Although these areas cover only 8,3% ofthe forest floor, 45 % of all spruce seedlings are found there. Beside site quality, regeneration density varies in relation to soil quality, elevation, exposition and the year when the old spruce stands died. This is particularly evident in the Rachel area where regeneration density is low. The main soil here consists of clay over compacted rubble. Also on large areas above 1250 m the forest graund is heavily covered with grass and old stands died after the seed year of 1995 which resulted in less seedlings in this area. To date there is little evidence of damage to seedlings. For example, evidence of brawsing of mountain ash saplings by deer and elk has reduced fram 44.7% in 1991 to just 14.4% in This is a significant reduction in browsing. Comparison with research results fram other high elevation forests concludes that disturbances are a natural factor in the pracess ofthe development of forest ecosystems. 171
171 8. Literaturverzeichnis ANONYMUS (1993): Climate News, World Meteorological Organization, 2, Jan ANONYMUS (1995): Firescan- Researching Taiga Fires as Natural Ecological Factors and Environmental Risks. Taiga-News no 13. ARBEITSKREIS STANDORTSKARTIERUNG IN DER ARBEITSGEMEINSCHAFT FORSTEINRICHTUNG (Hrsg., 1982): Forstliche Standortsaufnahme. Landwi rtschaftsverlag, Mü nster-hi Itru pp; 188 S. AUCLAIR, A. und T. CARTER (1993): Forest wildfires as arecent source of CO, at mid-high northern altitudes. Canadian Journal of Forest research 23; S BARTAK, M. (1995): Xylobiotic animals of the Bavarian Forest. Unveröffentlichter Projektbericht, Prag; 105 S. BARTHELMESS, A. (1972): Wald - Umwelt des Menschen. Alber/Freiburg-München; 332 S. BAYERISCHES STAATSMINISTERIUM FUER ERNAEHRUNG, LANDWIRTSCHAFT UND FORSTEN (1982): Richtlinien für die mittel- und langfristige Forstbetriebsplanung in der Bayerischen Staatsforstverwa Itu ng. BAYFORKLIM (1999): Klimaänderungen in Bayern und ihre Auswirkungen. Abschlussbericht des Bayerischen Klimaforschungsverbundes; 90 S. BECKER, T. und H. SCHROETER (2000): Ausbreitung von rindenbrütenden Borkenkäfern nach Sturmschäden. AFZ/DerWald, 6/2000: S BLASCHKE, M. (1999): Waldverjüngung in montanen und subalpinen Fichtenwäldern Mitteleuropas. Ergebnisse einer Literaturstudie unter besonderer Berücksichtigung der Verhältnisse in den Hochlagen des Bayerischen Waldes. Forst und Holz, 54 Jg., Nr. 15; S BOEHMER, H.-J. (1997): Zur Problematik des Mosaik-Zyklus-Begriffes. Natur und Landschaft, 72 Jg. Heft 7/8. BRANG, P. (1996): Experimentelle Untersuchungen zur Ansamungsökologie der Fichte im zwischenalpinen Gebirgswald. Beiheft Nr. 77 zur Schweizerischen Zeitschrift für Forstwesen, Zürich. BRUNNSCHWAIGER, A. (2000): Einfluss einer Borkenkäfergradation auf Artenvielfalt, Siedlungsdichte und Vitalität von Ameisen (Hymenoptera, Formicidae) im Nationalpark Bayerischer Wald. Unveröffentlichte Diplomarbeit Bayerische Julius-Maximilians Universität, Würzburg ; 113 S. BUELOW, G. v. (1964): Fichtennaturverjüngung in den Hochlagen des Bayerischen Waldes. AFZ; S BURSCHEL, P. und J. Huss (1997): Grundriss des Waldbaus. Berlin, Wien; 478 S. COCKBURN, A. (1995): Evolutionsökologie. Gustav Fischer Verlag, Stuttgart; 357 S. DELONG, C. S. und W. B. KESSLER (1999): Ecological characteristics of mature forest remnants left by wildfire. Forest Ecology and Management 131 (2000); S EICHRODT, R. (1970): Über die Bedeutung von Moderholz für die natürliche Verjüngung im subalpinen Fichtenwald.- Beiheft Nr. 45 zu: Zeits. d. Schweiz. Forstvereins; 122 S. ELLlNG, W., BAUER, E. KLEMM, G. und H. KOCH (1987): Klima und Böden - Waldstandorte. Wissenschaftliche Schriftenreihe, Bayer. Staatsministerium ELF, München; 255 S. FIRBAS, F. (1952): Spät und nacheiszeitliche Waldgeschichte Mitteleuropas nördlich der Alpen. Bd. I und 11, Jena. 172
172 FISCHER A., ABS G. und F. LENZ (1990): Natürliche Entwicklung von Waldbeständen nach Windwurf. Ansätze einer "Urwaldforschung" in der Bundesrepublik. Forstw. Cbl. 109; S FISCHER A. und H. JEHL (1999): Vegetationsentwicklung auf Sturmwurfflächen im Nationalpark Bayerischer Wald aus den Jahren In: Forstliche Forschungsberichte. Schriftenreihe der Forstwissenschaftlichen Fakultät der Technischen Universität München und der Bayerischen Landesanstalt für Wald und Forstwirtschaft; S FISCHER, A (1998): Verjüngung und Wiederbewaldung in den Hochlagen des Nationalparks Bayerischer Wald. In: Bayerische Landesanstalt für Forstwirtschaft (Hrsg): Borkenkäferproblematik im Nationalpark Bayerischer Wald; S GEITNER, K. (2000): Entwicklung der Verjüngung in den Hochlagen des Nationalparks Bayerischer Wald nach Borkenkäferbefall. Unveröffentlichte Diplomarbeit. Technische Universität München; 73 S. HABER, W. (1976): Nationalpark Bayerischer Wald. Entwicklungsplan. Gutachten des Lehrstuhls für Landschaftsökologie Weihenstephan; 62 S. HANSEN, A. J., SPIES, T. A., SWANSON, F. J. und J. F. OHMANN (1991): Conserving biodiversity in managed forests: lessons from natural forests. Bioscience 41; S HEURICH, M., REINELT, A. und FAHSE, L. (2001): Die Buchdruckermassenvermehrung im Nationalpark Bayerischer Wald. In: Nationa I pa rkverwa Itu ng Bayerischer Wa Id: Waldentwicklung im Bergwald nach Windwurf und Borkenkäferbefall; S HOHMANN, M. und F. ENGELS (1991): Was kommt nach dem Sturm? AFZ 12; S HOLENSTEIN, B. (1994): Sturmschäden 1990 in der Schweiz. BUWAL, Bern. Schriftenr. Umwelt 218; 40 S. HUBER, S. (2000): Reaktion der Kleinsäuger auf Kronendachverlichtung und Absterben alter Waldbestände in Folge einer Borkenkäferkalamität im Nationalpark Bayerischer Wald. Unveröffentlichte Diplomarbeit. Bayerische Julius Maximilians-Universität, Würzburg ; 119 S. IPCC WORKING GROUP II (2001): Climate Change 2001: Impacts, Adaptation and Vulnerability. IPCC WG2 Third Assessment Report. Summary for Policymakers; 22 S. JARDINE, K. (1995): The Carbon Bomb: Climate Change and the fate of the boreal forests. Bericht von Greenpeace International; 35 S. JEHL, H. (1995): Die Waldentwicklung auf Windwurfflächen im Nationalpark Bayerischer Wald. In: Nationalparkverwaltung Bayerischer Wald (Hrsg.): Nationalpark Bayerischer Wald - 25 Jahre auf dem Weg zum Naturwald. Grafenau; S JEHL, H. (2000): Wetterbeobachtung im Nationalpark. Unveröffentlichter Bericht der Nationa I pa rkverwa Itu ng. KIMMINS, H. (1987): Forest Ecology. Macmillan Publishing Company, New York; 531 S. KOEHLER, F. (1997): Bestandserfassung xylobionter Käfer im Nationalpark Bayerischer Wald (Insecta: Coleoptera). Beitr. z. Bayer. Entomofaunistik, Bamberg 2; S KOENIG, A., MOESSMER R. und A. BAEUMLER (1995): Waldbauliche Dokumentation der flächigen Sturmschäden des Frühjahres 1990 in Bayern und meteorologische Situation zur Schadenszeit. Berichte aus der Bayerischen Landesanstalt für Wald und Forstwirtschaft, 2; 335 S. 173
173 KORPEl, s. (1993): Vorkommen, Charakteristik und Folge der Entwicklungsstadien bzw. - phasen in den europäischen Urwäldern. symp. Urwälder/Forstl. Fak. Zvolen; s KORPEl, s. (1995): Die Urwälder der Westkarparten. G. Fischer, stuttgart; 310 s. KRANKINA, O. (1991): Forest fires in the former soviet Union: Past, present and future greenhouse gas contributions to the atmosphere. In: Ted s. Vinson and Tatyana P. Kolchugina (eds.). Proceedings of the International Workshop on Carbon Cycling in Boreal Forest an sub-arctic Ecosystems. U.5. Environmental Protection Agency, Global Change. KUHlMANN, M. (1999): Besiedlung von Windwürfen im Nationalpark Bayerischer Wald durch Wildbienen und aculeate Wespen (Hymenoptera Aculeata). Ber. Naturf. Ges., Bamberg; LAESSIG, Rund W. schoenenberger (1993): Forschung auf Sturmschadensflächen in der Schweiz. Forst und Holz 48 (9); s LAESSIG, R., EGLI, 5., ODERMAn, 0., schoenenberger W., stoeckli, B. und T. WOHlGEMUTH (1995): Beginn der Wiederbewaldung auf Windwurfflächen; Schweizer Zeitschrift Forstwesen 146, 11; s LAESSIG, R. und s. MOCAlOV (2000): Frequency and characteristics of severe storms in the Urals and their influence on the development, structure and management of the boreal forests. Forest Ecology and Management, 135; s LEDER, B. und M. PETRAK (2000): Bedeutung der Vogelbeere für Waldbau und Wild. LÖBF Mitteilungen, 1; LEIBUNDGUT, H. (1978): Europäische Urwälder der Bergstufe. Verlag Paul Haupt, Bern. LEIBUNDGUT, H. (1978): Über die Dynamik europäischer Urwälder. AFZ, München 33; MAYER, H. und E. On (2. Aufl. 1991): Gebirgswaldbau - Schutzwaldpflege. G. Fischer Verlag, stuttgart; 587 s. MINISTERIUM LAENDLICHER RAUM (2001): Ein Jahr nach "Lothar". Informationen zum Stand der Bewältigung der Sturmschäden ein Jahr nach "Lothar". Pressemitteilung ; 17 s. NUESSlEIN, s. (1996): Hochlageninventur Bayerischer Wald. Bayerische Landesanstalt für Wald und Forstwirtschaft, Freising; 16 s. NUESSlEIN, s. und G. FAISST (1998): WaIdentwicklung im Nationalpark Bayerischer Wald Totholzflächen und Waldverjüngung. Bayerische Landesanstalt für Wald und Forstwirtschaft, Freising; 24 s. NUESSlEIN, 5., FAISST, G., WEISSBACHER, A., MORITZ, K., ZIMMERMANN, L., BITIERSOHL., J., KENNEL, M., TROYCKE, A. und H. ADLER (1999): Zur Waldentwicklung im Nationalpark Bayerischer Wald Bayerische Landesanstalt für Wald und Forstwirtschaft; 47 s. OKOLOW, C. (1991): Der Einfluss der Forstwirtschaft auf die natürlichen Begrenzungsfaktoren des Buchdruckers. Seevögel 12, Sonderheft; 5; On, E., FREHNER, M., FREY, U., LUESCHER, P. (1997): Gebirgsnadelwälder. Haupt, Bern; 287 s. On, E., LUESCHER, F., FREHNER, M., BRANG, P. (1991): Verjüngungsökologische Besonderheiten im Gebirgsfichtenwald im Vergleich zur Bergwaldstufe. Schweizerische Zeitschrift Forstwesen, Zürich 142, 11; Ono, H.-J. (1994): Waldökologie. Ulmer, stuttgart; 391 s. PERRY, D. A. (1997): Disturbance, recovery and stability. Creating a forestry for the 2'" century: the science of ecosystem management. Island Press, Washtington D. c.;
174 PETERMANN, R. und P. SEIBERT (1979): Die Pflanzengesellschaften des Nationalparks Bayerischer Wald. Wiss. Schriftenr. Bayer. StMinELF, München; 142 S. PLATE, G. (1975): Ökologische Untersuchungen zur Verjüngung der Fichte. Diss. Ludwig Maximilian-Univ. München; 199 S. PRZYBILLA, M. (1993): Zur Regeneration der Fichte in den Altholzbeständen der Hochlagen des Nationalparkes Bayerischer Wald. Unveröffentlichte Diplomarbeit, Forstwissenschaftliche Fakultät der Albert Ludwigs-Universität, Freiburg ; 105 S. RALL, H. (1995): Die Wälder im Nationalpark Bayerischer Wald: Von forstwirtschaftlicher Prägung zur natürlichen Entwicklung. In: Nationalparkverwaltung Bayerischer Wald (Hrsg.): Nationalpark Bayerischer Wald - 25 Jahre auf dem Weg zum Naturwald, Grafenau; S REIF, A. und M. PRZYBILLA (1998): Regeneration der Fichte in den Hochlagen des Nationalparks Bayerischer Wald. AFZ/Der Wald, 8; S ROH MEDER, E. (1972): Das Saatgut in der Forstwirtschaft. Verlag Paul Parey, Hamburg; 273 S. ROTIMANN, M. (1986): Wind- und Sturmschäden im Wald. Sauerländer, Frankfurt; 128 S. ROMME, W. H. und D. G. DESPAIN (1989) The Yellowstone Fires. Scientifc American 261 (5); S RYHIEL, E. J., COULSON R. N., SHARPE, P. J., ALLEN T. F. H. und R. O. FLAMM (1988): Disturbance Propagation by Bark Beetles as an episodic Landscape Phenomenon. Journal of Landscape Ecology 1(3); S SCHERZlNGER, W. (1995): Artenschutzprojekt Auerhuhn im Nationalpark Bayerischer Wald. Unveröffentlichtes Manuskript; 77 S. SCHERZlNGER, W. (1996): Naturschutz im Wald. Ulmer, Stuttgart; 447 S. SCHERZlNGER, W. (1998): Sind Spechte "gute" Indikatoren der ökologischen Situation von Wäldern? Vogelwelt 119; S SCHERZlNGER, W. (1999): Natürliche WaIdentwicklung und Einnischung der Avifauna innerhalb der Langzeit-Dynamik am Beispiel des Inneren Bayerischen Waldes. Unveröffentlichtes Manuskript; 37 S. SCHMIDT-VOGT, H. (1985): Struktur und Dynamik natürlicher Fichtenwälder in der borealen Nadelwaldzone. Schweiz. Z. Forstwesen, Zürich,136/12; S SCHMIDT-VOGT, H. (1991): Die Fichte, Band 11/3. Verlag Paul Parey, Hamburg; 647 S. SCHOENENBERGER, W. (1988): Standortgemäße Aufforstung in hohen Lagen. In: Tagungsbericht zum Internationalen Arbeitstreffen "Gefährdung der Waldgrenze: Bedeutung und Zustand der Pioniergehölze. Bayerische forstliche Forschungs- und Versuchsanstalt München. SCHurn, P., SCHUCK, H. J. und B. STIMM (1992): Lexikon der Forstbotanik. Ecomed, Landsberg/Lech; 579 S. SIEGRIST, J. (2000): Natürliche Wiederbewaldung und Struktur eines ungeräumten Fichtenwindwurfes auf potentiellem Bergmischwaldstandort. In: Waldentwicklung im Nationalpark Berchtesgaden von 1983 bis Forschungsbericht; 146 S. SEYFERT, I. (1974): Zur Geschichte der Waldweidenutzung im Nationalparkbereich. Nationalpark, Grafenau, 4/75; S TRAVAN, J. und O. SCHMIDT (1994): Hinweise für den Schutz der Roten Waldameisen im bayerischen Staatswald. Bayerische Landesanstalt für Wald und Forstwirtschaft (Hrsg.). TUREK, F. J. (1961): Ökologische Beziehungen der Vögel und Gehölze. Verlag der Slowakischen Akademie der Wissenschaften, Bratislava; 330 S. 175
175 TUREK, F. J. (1967): Ökologische Beziehungen der Säugetiere und Gehölze. Verlag der Slowakischen Akademie der Wissenschaften, Bratislava; 210 S. TURNER, M. G., DALE V. H. und E. H. EVERHAM (1997): Fires, Hurricans, and Volcanoes: Comparing Large Disturbances. BioScience 47; S USOA FOREST SERVICE (H rsg., 2001): Forest I nsect and Disease Conditions in Alaska General Technical report R10-TP-86; 67 S. WAIER, C. (1996): Schädigung von Lycopodium annotinum L. im Nationalpark Bayerischer Wald - Vergleichsuntersuchungen zu Unveröffentlichte Diplomarbeit, Universität Regensburg ; 122 S. WELLENSTEIN, G. (2. Aufl., 1990): WaIdbewohnende Ameisen - ihre Bedeutung, ihre Biologie, ihre Hege und ihr Schutz. Deutsche Ameisenschutzwarte (Hrsg.), Allg. Zeitungsverlag GmbH, Kempten. WERMELINGER, B., DUELLI, P., OBRIST, M., OOERMATI, O. und M. SEIFERT (1995): Die Entwicklung der Fauna auf Windwurfflächen mit und ohne Holzräumung. Schweiz. Z. Forstwesen, Zürich, 146,11; S ZIERL, H. (1972): Der Hochwald. Forstwissenschaftliche Forschungen. Beihefte zum Forstwissenschaftlichen Centralblatt, Heft 33; 80 S. 176
176 Abbildungsverzeichnis Archiv Nationalparkverwaltung : 4, 10, 15, 53, 75, 76, 77, 78 Bäuml: 69 Heurich: 3, 6, 9,12,14,26,27,31,50,52,56,57, 58,59,61,62,63,64,65,66,70,71,72,73 Kiener: 1 Pöhlmann: 7, 60 Rall: 45 Scherzinger: 11, 13, 20, 79 Strunz: 16,55,68 Wasem: 30 Zellner: 2, 17,
177 Glossar Advektion: horizontaler Luftmassentransport. Bei Advektivfrösten wird, meist aus nördlichen Richtungen, Kaltluft zugeführt. Die Temperaturen in dieser Kaltluft sind um so niedriger, je größer die Meereshöhe des betrachteten Standortes ist. Anthropogen: durch den Menschen verursacht. Assimilate: körpereigene organische Substanzen, die aus anorganischen Substanzen synthetisiert werden (z. B. DNA, Aminosäuren, Kohlenhydrate). Ausapern: wenn sich beim Abschmelzen der Schneedecke erste schneefreie Stellen ausbilden. Autochthon: bodenständig, an Ort und Stelle entstanden. Bestand: Kollektiv von Bäumen, die ähnliche Merkmale aufweisen, eine zusammenhängende Mindestfläche bestocken und ähnliche Behandlung erfordern. Bodenform: Böden mit gleichen oder ähnlichen, für die Vegetation stabilen Merkmalen hinsichtlich ökologischer Eigenschaften. Boreal: nordisch, speziell kontinental-kühle, nördliche Nadelwaldzone. Brusthöhendurchmesser (BHD): Durchmesser eines Baumes in einer Höhe von 1.30 m. Color Infrarot (CIR) Luftbilder: Falschfarbenbilder, die Unterschiede in den Eigenschaften von Oberflächen (z. B. Vitalität von Waldbeständen) durch entsprechende Farbgebung besonders deutlich hervorheben. Deposition: Ablagerung von Stoffen, i. e. S. von Schadstoffen, auf Oberflächen. Man unterscheidet zwischen nasser und trockener Deposition. Erste erfolgt über Regen, Nebel, Schnee usw. Letzte über Transport von Luftverunreinigungen auf Oberflächen (Stäube). Diasporenbank: Gesamtheit der im Boden ruhenden, nicht gekeimten Samen und Sporen. V. a. die Diasporen vieler Pionierpflanzen sind in der Lage, lange Zeit zu überdauern, ohne ihre Keimfähigkeit zu verlieren. Dispersion: Ausbreitung, Verteilung. Diversität: quantitativer Ausdruck für strukturelle, räumliche und artenmäßige Vielfalt eines Ökosystems; Mannigfaltigkeit (Artendiversität, Stru ktu rdiversität). Edaphisch: Zum Boden gehörend; wird in Bezug auf die Organismen des Bodens und die abiotischen Bodenfaktoren gebraucht. Erntefestmeter ohne Rinde (Efm o. R.): Maßeinheit für Planung, Einschlag, Verkauf des Holzes, entspricht einem Kubikmeter. Femelschlag: Eine Form des Hochwaldbetriebs, bei dem der Bestand durch gruppen- und horst -: weise Entnahme von Altbäumen aufgelichtet wird. Die in den Bestandeslücken entstehenden Jungwuchsgruppen werden durch weitere Auflichtung so lange vergrößert, bis sie zusammenwachsen. Forstoperat: Alte Bezeichnung für Forsteinrichtungswerk. Bericht über die Ergebnisse einer Waldinventur und die daraus abgeleiteten Planungen. Fruktifikation: Ausbildung von Früchten oder Samen bei Pflanzen. Georeferenzierung: lagegenaue Einpassung von räumlichen Daten in ein Koordinatensystem. Gradation: Gesamtablauf der Massenvermehrung einer Population. Habitat: Ressourcen, Strukturen und Umweltmerkmale, die den Lebensraum eines Individuums bzw. einer Art ausmachen. Kambium: Wachstumszellen bzw. Wachstumsschicht in einer Pflanze. Bei Bäumen bildet das Kambium jedes Jahr sowohl neue Rinden - als auch Holzzellen. Dadurch entsteht ein sog. "Jahrring", eine neue Holzschicht, der Baum wächst und wird dicker. Kluppschwelle: Mindestdurchmesser in Brusthöhe, ab der ein Baum auf einer bestimmten Probefläche erfasst wird. k-stratege: Lebensraumnutzung unterhalb der Kapazitätsgrenze, bei relativer Langlebigkeit, ge- 178
178 ringer bzw. relativ konstanter Fortpflanzungsrate (meist "konservative" Arten wie Schwarzspecht, Uhu, Großraubtiere, Auerhuhn). Mast: Das Fruchten der Waldbäume. Ursprünglich nur auf Buche und Eiche bezogen, da Bucheckern und Eicheln durch Vieheintrieb genutzt wurden. Median: Der Wert, der die nach Größe sortierten Messwerte exakt in eine obere und untere Hälfte teilt. Monitoring: Langzeitbeobachtung, Dauerbeobachtung. Mycel: Als Mycel bezeichnet man die Gesamtheit der Pilzfäden, die aus langgestreckten, oft verzweigten Zellen bestehen (Hyphen). Das Mycel bildet das Pilzgeflecht oder den Thallus (=typische Gestalt niederer Pflanzen). Organische Auflage: Humusauflagerung auf Mineralboden (Laub- und Nadelstreu, abgestorbene Pflanzenteile). Pflanzengesellschaft: Durch eine charakteristische Artenkombination definierte Vegetationseinheit, die in der Synsystematik als Assoziation bezeichnet wird. Verwandte Assoziationen werden in übergeordnete Einheiten (Verband, Ordnung, Klasse) zusammengefasst. Pheromone: Stoffe, die als chemische Auslöser zwischen Individuen der gleichen Art wirken und dadurch intraspezifische Koordination bedingen. In Pheromonfallen werden buchdruckerspezifische Lockstoffe genutzt, um die Käfer anzulocken und zu fangen. Phytomasse: Biomasse der lebenden Pflanzen. Pionierarten: Erstbesiedler. Pflanzen- und Tierarten, die neu entstandene Lebensräume besiedeln und die dort vorherrschenden Extrembedingungen tolerieren können. Plenterung: Waldbehandlung, bei der als Folge einer gezielten Entnahme von einzelnen, meist starken Bäumen kleinflächig alle Baumarten eines Bestandes in den verschiedenen Schichten, Alters- und Entwicklungsstufen gemischt vorkommen. Die Entnahme der Bäume wirkt sich somit gleichzeitig als Ernte-, Verjüngungs- und Pflegemaßnahme aus. Podsolierung: Auswaschung von organischen Substanzen und Eisenhydroxiden durch Sickerwasser vom humosen Oberboden in tiefere Boden horizonte. Population: Gesamtheit aller Individuen einer Art oder Rasse in einem geografisch begrenzten Verbreitungsgebiet, wobei die Einzelindividuen unbegrenzt untereinander fortpflanzungsfähig sind. Prädation: Räubertum; System in dem eine Art (Räuber) eine andere (Beute) als Nahrung nutzt. Prozessschutz: Sicherung einer Entwicklung ohne direkte menschliche Einflussnahme. Rannen-Verjüngung: standortbegünstigtes Aufwachsen von Waldbäumen auf dem vermodernden Holz von abgestorbenen liegenden Bäumen. Reproduktion: Vermehrung, Fortpflanzung. Rohhumusartiger Moder: Organische Auflage, die sich unter ungünstigen Bedingungen ausbildet und nur eine geringe biologische Aktivität aufweist. r-stratege: Lebensraumnutzung bis an die Kapazitätsgrenze, bei relativer Kurzlebigkeit und kurzfristigem Reaktionsvermögen auf neues Lebensraum- bzw. Nahrungsangebot durch massive Steigerung der Fortpflanzungsrate (in der Regel "Pionierarten" wie Reh, Wühlmäuse, Schleiereule, Wildschwein) Schichtung: Vertikale Struktur des Waldes. Schlankheitsgrad: Quotient aus Höhe und Durchmesser eines Baumes (H/D-Verhältnis). Maß für die Stabilität von Bäumen gegenüber mechanischen Belastungen (Wind, Schnee). Fichten mit einem Stabilitätsindex < 80 (70) gelten als stabil. Schmelztrichter: unmittelbare Umgebung von Baumstämmen, in der der Schnee im Frühling zuerst abschmilzt. 179
179 Sprengmast: schüttere Samenproduktion, an der nur wenige Waldbäume beteiligt sind. Stereoskop: Optisches Gerät mit zwei Okularen, das durch Kombination sich überlappender Bilder ermöglicht, beim Betrachten einen räumlichen Eindruck zu erhalten. Stubben: Baumstümpfe, die nach Fällung oder Umbrechen der Bäume verbleiben. Synsystematik: Gliederung von floristisch definierten Vegetationseinheiten (Pflanzengesellschaften) nach Art eines hierarchischen Systems. Transpiration: Wasserabgabe von Pflanzen an die Atmosphäre. Trift: Transport von nicht miteinander verbundenen Stammabschnitten auf Bächen, die zu diesem Zweck häufig ausgebaut und aufgestaut (Klausen) wurden. Verjüngungszeitraum: Zeitraum, der zu einer vollständigen Verjüngung des Bestandes notwendig ist. Vollmast: ergiebige Samenschüttung, bei der ein Großteil der Individuen einer Baumart beteiligt sind. Waldinventur: Regelmäßige Zustandserhebung zur Gewinnung relevanter Informationen über den Aufbau und die Struktur der Wälder. Wurzelanläufe: Übergangsbereich zwischen Stamm und Wurzelsystem, der durch den Ansatz starker Seitenwurzeln gekennzeichnet ist. Xylobionte: direkt oder indirekt vom Holz lebende Organismen (betrifft vor allem Totholzbewohner unter Pilzen und Insekten). Managementzonen im Nationalpark: Zone I (Naturzone nach IUCN): Diese Zone beinhaltet sämtliche Flächen des Nationalparkes, auf denen grundsätzlich keine menschlichen Maßnahmen vorgesehen sind. Zone 11 (Entwicklungszone nach IUCN): Die Wälder in Zone II sollen innerhalb eines Übergangszeitraumes schrittweise der natürlichen Entwicklung überlassen und somit der Zone I zugeführt werden. Zone 11 a (Hochwald im Falkenstein-Rachel Gebiet): Hier sollen bis zum Jahr 2017 wirksame Borken käferbekäm pfu ngsmaßna h men ergriffen werden, um einen raschen und flächigen Zerfall der Fichtenhochlagenwälder zu verhindern. Zone 11 b (Schutzzone um den Hochlagenwald im Falkenstein-Rachel-Gebiet): Hier sollen zum Schutz der Hochlagenwälder wirksame Borkenkäferbekämpfungsmaßnahmen ergriffen werden. Zone 11 c (Gebiet zwischen der Schutzzone um den Hochlagenwald im Falkenstein-Rachel Gebiet und dem Randbereich): Die Wälder dieser Zone sollen möglichst bald einer natürlichen Entwicklung überlassen werden. Strukturreiche, naturnah aufgebaute Mischbestände sollen bereits jetzt unberührt bleiben. Zone 111, Randbereich (Naturzone mit Managementmaßnahmen nach IUCN): Hier sollen auf Dauer alle erforderlichen ordnungsgemäßen und wirksamen Waidschutzmaßnahmen ergriffen werden, um die an den Nationalpark angrenzenden Wälder vor Schäden, die auf eine unbeeinflusste Waldentwicklung im Nationalpark zurückgehen, zu bewahren. Zone IV (Erholungszone nach IUCN): In diesem Bereich soll durch Managementmaßnahmen sichergestellt werden, dass die Verkehrssicherheit und die Funktionen der Besuchereinrichtungen gewährleistet sind. Zoochorie: Samenverbreitung durch Tiere (aktive Aufnahme durch Verzehr von Früchten, passive durch Transport im Fell). 180
180 Publikationsliste Wissenschaftliche Reihen Nationalpark Bayerischer Wald Bisher erschienene Hefte: A) "Nationalpark Bayerischer Wald" Nr. 1 Klima und Böden - Waldstandorte von W. Elling, E. Bauer, G. Klemm und H. Koch, 2. Auflage , zahlreiche Tabellen, Abbildungen, 11 mehrfarbige Karten, Maßstab 1 : Nr. 2 Rauhfuß-Hühner von W. Scherzinger, , zahlreiche Abbildungen, Tabellen und Grafiken (vergriffen) Nr. 3 Fischottervon I. Hodl-Rohn und R. Becker, , zahlreiche Abbildungen, Tabellen und Grafiken Nr. 4 Pflanzengesellschaften des Nationalparks Bayer. Wald von R. Petermann und P. Seibert, , zahlreiche Tabellen, z. T. mehrfarbige Abbildungen, Vegetationskarte im Maßstab 1 : Nr. 5 Witterung und Klima im Nationalpark Bayer. Wald - dargestellt anhand der dreijährigen Datenreihe der Klimastation Waldhäuser ( ), von E.-M. Noack, Seiten, zahlreiche Abbildungen, Tabellen und Grafiken N r. 6 Eiszeitliche Formen und Ablagerungen von U. Hauner, , zahlreiche Tabellen, Grafiken und Karten, z. T. mehrfarbige Abbildungen Nr. 7 Rot- und Rehwild im Nationalpark Bayer. Waldvon U. Wotschikowsky, , zahlreiche Abbildungen, Tabellen und Grafiken, mehrfarbige Karte der Schälschäden Nr. 8 Sammeln und Langlaufen im Nationalpark Bayer. Wald von U. Ammer, A. Pledl, H. Riederer und K. Thiele, , zahlreiche Abbildungen, Tabellen, Grafiken und Karten Nr. 9 Spechte im Nationalpark Bayer. Wald von Dr. W. Scherzinger, , zahlreiche Abbildungen, Tabellen, Grafiken und Karten Nr. 10 Ökologische Wertanalyse - Gutachten zur Waldpflegeplanung im Nationalpark Bayer. Wald auf der Grundlage einer ökologischen Wertanalyse von U. Ammer und H. Utschik, , zahlreiche Abbildungen, Tabellen und Grafiken, 4 mehrfarbige Karten im Maßstab 1: (vergriffen) Nr.11 Eine Landschaft wird Nationalpark. Die Geschichte des Waldes und seiner Besiedlung von R. Strobl, Entstehungsgeschichte des Nationalparks Bayer. Wald und Entwicklung seit 1969 von M. Haug, 2. Auflage, , zahlreiche Abbildungen, Karten, umfangreiche Listen der erschienenen Literatur und der laufenden und abgeschlossenen Forschungsarbeiten Nr.12 Vögel im Urwald Die Vogelwelt der Urwaldgebiete im Inneren Bayer. Wald von Dr. W. Scherzinger, , zahlreiche Abbildungen, Tabellen und Grafiken Nr.13 Epiphytische Flechten im Nationalpark Bayer. Wald von M. Macher, , 19 Fotos, 33 Abbildungen und 32 Tabellen o. Nr. Nationalpark Bayerischer Wald - 25 Jahre auf dem Weg zum Naturwald. Berichte über die wissenschaftliche Beobachtung der Waldentwicklung. Nationalparkverwaltung Bayer. Wald (Hrsg.), , 69 Abb., 23 Tab., 51 Fotos, Karte M 1 :
181 B) Tagungsberichte 1. Wasserhaushaltssysteme naturnaher, kleiner Einzugsgebiete, Mai , zahlreiche Abbildungen, Tabellen und Grafiken (vergriffen) 2. Forschungsziele, Planung und Koordination von Forschungsprogrammen in mitteleuropäischen Nationalparken und vergleichbaren Schutzgebieten, Mai , zahlreiche Abbildungen, Tabellen und Grafiken Säure- und Stoffeintrag mit dem Niederschlag von M. Bosse, , 38 Abbildungen, 16 Tabellen Validierung, Anpassung und Modifizierung des forst-hydrologischen Modells BROOK von M. Kennel, , 23 Abbildungen, 12 Tabellen Die Entwicklung der Schneedecke im Einzugsgebiet der Großen Ohe von A. 5tang, ; 5., 31 Abbildungen, 2 Fotos, 15 Tabellen 3. Schutz der Tier- und Pflanzenwelt und 5. Niederschlags- und Abflußauswertung der ihrer natürlichen Lebensräume, Juni , 1 Tabelle, 10 Abbildungen 1 O-jährigen Meßreihe für das Einzugsgebiet der Großen Ohe von 5. Thums, , 55 Abbildungen, 18 Tabellen 4. NaturerIeben, Naturerkenntnis, naturkundliche Bildung in Nationalparken und 6. Hydrochemische Dynamik und Versaue vergleichbaren Schutzgebieten, Juni , zahlreiche Abbildungen, Tabellen und Grafiken rungsmechanismen im Quellgebiet der Großen Ohe von I. Haag, , 47 Abb.,Anh. 5.1 Wald und Wasser, Prozesse im Wasser- 7. Einzugsgebiet Große Ohe - 20 Jahre und Stoffkreislaufvon Waldgebieten, September , zahlreiche Abbildungen, Tabellen und Grafiken hydrologische Forschung im Nationalpark Bayerischer Wald von der NPV, LWF, LfW und dem Lehrstuhl f. Bioklimatologie der TU München Wald und Wasser, Prozesse im Wasserund Stoffkreislaufvon Waldgebieten, September Teil 11: Wasser- und Stoffbilanzen, , zahlreiche Abbildungen, Tabellen und Grafiken Schläfer und Bilche, Tagungsbericht 1. Intern. Bilch-Kolloquium, 5t. Oswald C) Wasserhaushalt und Stoffbilanzen im naturnahen Einzugsgebiet der Großen Ohe 1. Die Ermittlung des Gebietsniederschlags zur Lösung hydrologischer Bilanzen von U. Teichmann, , 38 Abbildungen, 3 Fotos, 10 Tabellen D) Arbeitsberichte des Bayer. Landesamtes für Wald und Forstwirtschaft und der Nationalparkverwaltung Bayer. Wald 1. Forschung im Nationalpark Bayer. Wald von R. Holzapfl und K. Thiele, , zahlreiche Abbildungen, Tabellen und Grafiken 182














182 Anlage 1 Totholzentwicklung iin Nationalpark Bayerischer Wald Bork~n käferbefa 11 _ Flächenzugang Totholz 2000 _ Fläch~nzugang Totholz 1999 FlächenzugangTotholz bis 1999 _ Ausgeräumt~ Käfernester 2000 L.J Ausgeräu mt~ Käfernester bis 1999 Höhenstufen _ Hochlage Obere Ranglage Tallage Untere Hanglage _ Nationalpark-Enklaven Erweiterungsgebiet Gewässer (Quelle : ATKIS Bayer. landesvermessungsamt) S i ~lungen (Quelle: ATKIS Bayer. landesvermessungsamt) M.Nationalparkaußengrenze ~ Gr~nze zwischen Randbereich und Naturzone,, -'..! Gr~nz~ d~r Hochlagen ~..,. ~., Wand~rwege o Meter Bearbeitung: Marco Heurich Nationalpark Bayerischer Wald
183 Anlage 2 a Die Waldentwicklung nach Windwurf in den Hochlagen des Nationalparks Bayerischer Wald Waldort: XXXIII 2a Gfeichtethöh ( m ü.nn) Koordinaten (TK 1 : , NP Bayer. Wald): 54268/46125 Aufnahmezeit: Hans Jehl Probeauadrat :1 1:1 1:1 1 :1 Entwicklungsphase qeschlossenes Altholz steh. Totholz belassener Windwurf steh. Toth. aeschlossenes Altholz Artenzah l Bl e (ohne tote Bäume) B : t B : S : 1 i K M k Deckung Bl : e (ohne tote Bäume) B i B t S : K : M : /0 5 t Bäume und Sträucber Picea abies Bl 4 2b 2b 2b 4 4 2b b b 3 59 (tot) 2b 2b 2b 3 2a 2b : 2a 3 3 2b 2a 2a 2a 31 B2 2a : 2a 2a 2b 2b 2a 2a 2b B3 : : 2a 2a 2a 1 2a (tot) : : S + r + 1 2b 2a 1 2a 2a 23 K 2m 2a a a 1 2m 2m r + r Fagus sylvatica K r 3 Sorbus aucuparia S 1 : r 1 2a 2a 1 28 K a 1 : 1 r : + + r + r + + r Betula pubescens K : : r : : 3 Betula pendula K r : 3 Salix caprea K r : : 3 Arten der Scblagfluren und VQr... aldgebüscbe (I'lIitrifizieruogmiger)' V Rubus idaeus + r r :1 : I: : 2m 2a 2a 1 2a 2a 2m : 2m + r r 1 1 r 54 0 Epilobium ang ustifolium + + : 1 2m r 31 Eeucbte- und Ilerdichtungmiger: Ca rex curta :1 2m 18 Juncus effusus + : 1 : 5 Arten mit Ilerbreitungsscbwerpuokt in Ekbten- Iannen- und Ekbtenwäldero; (0) Vaccinium myrtillus 2b 2b 2m 2a 2a 2a 2b 2b 2a 2a 1 2a 2a 2a 2m 2m 2m 2a 2m 2b 5 4 2b 2a 2a 2a 2a 2a 2b 2m 2b 2b 2a 2a 2a 2a 2a 2a 2b 100 V Calamagrostis villosa 2m 2a 2a 2a 2m 2b 2b a b 2a 2a 2m 2m 2m 3 2a 2a 2a 3 2a 97 Deschampsia flexuosa 1 2m 1 1 2m 2a 2m 2m 2m 1 2a 1 2m 2m : + 1 2m 2a 2m 2a 2a 1 2m 2m 2m 2m 1 2m 2m 2m 2m 2a 82 Athyrium distentifolium r m : 2m r 2a + 1 2m : + 1 2b 2b 2b 2a 2a 62 V Homogyne alpina : m 1 1 2m 2m 2m 41 Maianthemum bifolium m 1 + 2m V Soldanella montana 2m + 2m 1 1 2m 2m + 2m Trientalis europaea m 1 1 : : m 28 0 Vaccinium vitis-idaea 1 : : Lycopodium annotinum 1 2m 2m : 1 1 2m : : 15 V Blechnum spicant : : r Streptopus amplexifolius r r : : : 5 0 Huperzia se lago r : ; 3 Arten mit Ilerbreitungsscbwerpunkt in Buchen- und Bucben- Ianoenwäldem' : :. Dryopteris dilatata m 2m m 2a 2m 1 1 2m 1 : : Luzu la sylvatica 2b 2b 2b 2b 2b 2b 2a 2a 2a 2b 1 1 2a : 2m 1 2m m 2m 2m 1 2a 2a 90 Oxalis acetosella 2m 2a 2m 2a 2m 2m 2m 2m 2m 2m 2m 2a 2m : 2m 2m 2m 2m 2m 2m m m 2m 2m Luzula pilosa Gymnocarpium dryopteris r + + r + r Thelypteris phegopteris : : 1 21 Stellaria nemorum 1 : 2a 2a : : 1 2m 2m 15 V Prenanthes purpurea r : : + r r 10 M Petasites albus : : Dryopteris carthusiana + : : : ; 3 I: ~ Descha mpsia cespitosa : : : : 2m Senecio fuchsii : r : : : r + 8 Agrostis canina 2m : : : 2a 2m 8 Melampyrum pratense : : : Caltha palustris : : : : Viola palustris : : : : 2m 2m 5 Rumex alpestris : r Juncus filiformis + : 3 Ca rex echinata : + : : : 3 Carex ovalis + : 3 Cardamine amara : + 3
184 An lage 2 b Die Waldentwicklung nach Windwurf in den Hochlagen des Nationalparks Bayerischer Wald Waldort: XXXIII 2a Gfeichtethöh ( m ü.nn) Koordinaten (TK 1 : , NP Bayer. Wald): 54268/46125 Aufnahmezeit: August 1997 Hans Jehl Probequadrat : : :1 I: 1 I: 1 1:1 Entwicklungsp hase geschlossenes Altholz stehendes Totholz belassener Windwurf ste hendes Totholz gesch lossenes Altholz Artenza hl B e (ohne tote Bäume) B t B S i K g M k Deckung B e (ohne t ot e Bäume) B i B t K M : Ofo 5 t Bäume und St[äu~be[ Picea abies B1 5 2b b : b 3 2b 46 (tot) 2a 2a 2m : 1 : 2b 2a : 4 2a + 23 B2 : : 2a 2a 1 2a 2a 1 1 2a 2 1 (tot) : + 3 B3 : + 2a 1 1 2a 2a 1 2a 2 1 (tot) :., m 2a r r + 2a + 3 2a 2a b 1 2m 2a 2a : + 1 2a + 56 K 2b 2a m 2a 2a : + r r r r + 2a 1 2a Sorbus aucuparia B3 2a 2a 2a a 2a : m 1 2a 2a + 39 K + r r r + r r r r Betu la pendula 5 r + : : 5 K r : 3 Populu s tremula K : r : 3 Arten der Sc hla~ N it[ifizi e[u n gwgcl; V Rubus idaeus I a 2m 2m : 2a 2a 3 2b 2a 2a 2b 2b 1 2a 2a + 1 r :1 I: Epilobium angustifolium : : + 1 2m 2m m r : 41 Feuchtezeiger und Verdichtung~ Carex curta 1 2m r ; : + 10 Carex echinata : : + 3 Juncus effusus : + : : 3 Art en mit \le[breitungillb.yillpun~t in Eicbten-Iannen- und Ei~btenwäldern: (0) Vaccinium myrtillus 3 3 2b 2b 3 3 2a 2a 3 2m 2m 1 1 2m 2a 2m 1 2m 2m 2m : 2a a 2m 3 2b 2b 2a 2b 2b 2a 2a 2a 2a 2a V Calamagrostis villosa 2m 2a 2m 2m 2a 2a 2b 2b 2m a m 2b a 2a 2a 2m 2m 2a 1 2a 3 2a 2a 100 Athyrium distentifolium 2a r r a 1 2a a 2a 2m 2m : r r r b 2b 2a 1 92 Deschampsia flexuosa 2m m 2m : 1 1 2m 2b 2a 2a 3 2m 2a 2m 2a 2m 1 2m 2m 2m 77 V Homogynealpina 1 r r r + + r 2m 1 2m m 2m 2m 44 V Soldanel la montana 1 + r 1 r + r + 2m Trientalis europaea + 1 2m Lycopodium annotinum r 2m 2m m r 23 Maianthemum bifolium m + 1 : 18 0 Vaccinium vitis-idaea V Blechnum spicant r Streptopus amplexifolius r r : : 5 Arten mit \le[b[eitungss~bwerpy nkt in By ~ben - und Bu~ben-Tannenwäldero: Dryopteris dilatata Luzula sylvatica 3 2b 3 4 2b 2b 3 2a 2a 2a + 2a 2m m b 1 + 2m 1 2m + 2a 2a 2a 1 92 Oxalis acetosella 2m 2m 2m 2m 2m 2m 2m 2m 2m m m 2m a m Lu zu la pilosa r : Gymnocarpium dryopteris 1 r + : Thelypteris phegopteris r + + ; Senecio fuchsii r r r r + ; : r 1 18 Stellaria nemorum + 2m 2m 1 : V Prenanthes purpurea r : 1 13 (V) Petasites albus + 10!kgJ.citeI.; Deschampsia cespitosa : : 2m 1 5 Agrostis canina 2m : 2a 2m 1 5 Epilobium palustris : r : : 3 Melampyrum pratense : : : + 3 Caltha palustris : : 1 3 Viola palustris : 2m 2m 3 Cicerbita alpina : r 3 Zeiqerwerte (Berechnunq quantitativ, d.h. unter Berücksichtiqunq der Artmächtiqkeiten) Licht 4,1 4,2 4,9 4,1 4,0 4,2 4,6 4,1 4,4 4,6 4,8 4,7 5,0 : 4,6 5,2 5,5 5,1 4,9 5,1 5,0 5,1 4,9 5,1 4,8 5,3 5,3 5,3 5,1 5,1 4,4 4,6 5,0 5,4 5,1 4,6 4,6 4,6 4,9 4,9 Temperatur 3,9 3,9 4,1 3,9 3,9 4,0 3,8 3,8 3,8 3,8 3,9 3,7 4,2 4,1 4,0 4,0 4,0 4,1 4,1 3,8 4,0 3,8 4,0 4,0 4,0 4,0 4,0 4,0 4,0 4,0 4,0 3,8 4,2 4,1 3,8 3,8 3,9 4,0 3,7 Kontinentalität 3,5 3,4 3,6 3,4 3,5 3,6 3,4 3,1 3,4 3,6 3,8 3,4 3,6 3,8 3,9 4,1 3,9 3,6 4,0 3,6 4,0 3,8 3,6 3,7 3,4 3,5 3,5 3,3 3,7 3,4 3,5 3,1 3,7 3,8 3,5 3,6 3,5 3,5 3,1











185 Anlage 3 a Natürliche Waldentwicklung nach Windwurf in den Hochlagen des Nationalparks Bayerischer Wald Waldort: 33/2 Gfeichtethöh ( m ü. NN) Aufnahmejahr: 1992 erstellt: Hans Jehl \ ;.. 0 '., ~.,.. ": " <.,. ~., 0, 7q. ''';11, :1 o 0,. ~~~~~~~.~~~~ ~~ -L~~~L~~~~~~~,*-,~_e~~~~~_~~~,~~~ j;;ä-~~~~~l~w~~ ~~ Legende S ta m mfu ß-/Kronen ka rte - Fichte _ Vogelbeere _ Bi rke, Aspe, Weide Baum,BHD?,10cm, + Baum, BHD < 10cm, h?, l m X Baum, BHD < 10 cm, h < 1 m o stehender toter Baum o Baumstumpf ~ umgestürzter Baum mit Wurze lteller o Kronenschirmfläche =s: Quellsumpf = (Rücke)weg === Bach Legende Seitenansicht 1 11 Fichte lebend t Fichte tot, Vogelbeere Q Birke, Aspe, Weide Bäume sind erst ab einer Höhe von 1 m dargestellt. f-- 10m----j Maßstab 1 : o ~ Nationalpark ~ Bayerischer Wald ~

 Aufnahmejahr: 1997 erstellt: Hans Jehl, 07.")


weg == Bach Legende")


186 Anlage 3 b Natürliche Waldentwicklung nach Windwurf in den Hochlagen des Nationalparks Bayerischer Wald Waldort: 33/2 Gfeichtethöh ( m ü. NN) Aufnahmejahr: 1997 erstellt: Hans Jehl, Kronenkarte o o._ Legende Stammfuß-/Kronenka rte _ Fichte Vogelbeere Birke, Aspe, Weide Baum, BHD 2: 10 cm, + Baum, BHD < 10 cm, h 2: 1 m X Baum, BHD < 10 cm, h < 1 m o stehender toter Bau m o Baumstu mpf ~ umgestürzter Bau m mit Wurzelteller o Kronenschirmfläche == Quel lsumpf = (Rücke)weg == Bach Legende Seitenansicht! 11 Fichte lebend t Fichte tot, Vogelbeere o Birke, Aspe, Weide Bäume sind erst ab ei ner Höhe von 1 m dargestellt I-- 10 m--l Maßstab 1 : '_~_---il-, ,-_-,---_ o. " ". o ",. f ~ --+~_~!L~L_...l---,'i ~ _~:,--~.---'. L\"":"" " 2~~~c:::..='1\ilCiL_~_-----'L_~+;tL_~ ~ ~.~O:.L--j/Lr~ _~"---.L ---'----.J Nationalpark Bayerischer Wald
BORKENKÄFER. Gefahr in Verzug!!
 BORKENKÄFER nach Manfred Wolf, Forstdirektion Niederbayern Oberpfalz 2004 Gefahr in Verzug!! Biologie Lebensweise der Fichten-Borkenkäfer Borkenkäfer fliegen ab dem Frühjahr auf geschwächte stehende Bäume
BORKENKÄFER nach Manfred Wolf, Forstdirektion Niederbayern Oberpfalz 2004 Gefahr in Verzug!! Biologie Lebensweise der Fichten-Borkenkäfer Borkenkäfer fliegen ab dem Frühjahr auf geschwächte stehende Bäume
B orkenkäfer in K ürze
 B orkenkäfer in K ürze - Biologie, Befall, Bekämpfung - Der Borkenkäfer? Weltweit >6.000 Arten Europa rd. 150 Arten Buchdrucker Großer Buchdrucker Ips typographus Kleiner Buchdrucker Ips amitinus Absturz
B orkenkäfer in K ürze - Biologie, Befall, Bekämpfung - Der Borkenkäfer? Weltweit >6.000 Arten Europa rd. 150 Arten Buchdrucker Großer Buchdrucker Ips typographus Kleiner Buchdrucker Ips amitinus Absturz
Abb. 1 Mittleres Eintrittsdatum der maximalen Schneedeckenhöhe Zeitraum 1961/90.
 Abb. 1 Mittleres Eintrittsdatum der maximalen Schneedeckenhöhe Zeitraum 1961/90. Abb. 2 Mittlere Schneedeckendauer Zeitraum 1961/90. Den Kartendarstellungen liegen ca. 550 geprüfte, vollständige 30-jährige
Abb. 1 Mittleres Eintrittsdatum der maximalen Schneedeckenhöhe Zeitraum 1961/90. Abb. 2 Mittlere Schneedeckendauer Zeitraum 1961/90. Den Kartendarstellungen liegen ca. 550 geprüfte, vollständige 30-jährige
Witterungsbericht. - Frühjahr
 Witterungsbericht - Frühjahr 2013 - Witterungsbericht Frühjahr 2013 Erstellt: Juni 2013 Thüringer Landesanstalt für Umwelt und Geologie - Thüringer Klimaagentur - Göschwitzer Str. 41 07745 Jena Email:
Witterungsbericht - Frühjahr 2013 - Witterungsbericht Frühjahr 2013 Erstellt: Juni 2013 Thüringer Landesanstalt für Umwelt und Geologie - Thüringer Klimaagentur - Göschwitzer Str. 41 07745 Jena Email:
Newsletter. Krummzähniger Tannenborkenkäfer. 1. Beschreibung 2. Verbreitung 3. Lebensweise 4. Befallsmerkmale und Schadwirkung 5.
 Newsletter Newsletter 4/2016 WBV Bad Kötzting Krummzähniger Tannenborkenkäfer 1. Beschreibung 2. Verbreitung 3. Lebensweise 4. Befallsmerkmale und Schadwirkung 5. Bekämpfung 1 1. Beschreibung Der Krummzähniger
Newsletter Newsletter 4/2016 WBV Bad Kötzting Krummzähniger Tannenborkenkäfer 1. Beschreibung 2. Verbreitung 3. Lebensweise 4. Befallsmerkmale und Schadwirkung 5. Bekämpfung 1 1. Beschreibung Der Krummzähniger
nec praeteritum tempus umquam revertitur; nec quid sequatur sciri potest!!beobachtungszeitraum: 29. Juli bis 04. August 2015
 05.08.2015 nec praeteritum tempus umquam revertitur; nec quid sequatur sciri potest!!beobachtungszeitraum: 29. Juli bis 04. August 2015 Die aktuelle Lage und die vergangene Zeit kehrt weder je zurück noch
05.08.2015 nec praeteritum tempus umquam revertitur; nec quid sequatur sciri potest!!beobachtungszeitraum: 29. Juli bis 04. August 2015 Die aktuelle Lage und die vergangene Zeit kehrt weder je zurück noch
Waldsterben Was wurde daraus? Beobachtungen aus dem Stift Schlägl
 Waldsterben Was wurde daraus? Beobachtungen aus dem Stift Schlägl Rahmenbedingungen Höhenlage: 600m 1378m (Plöckenstein) Hauptwindrichtung: W und N (Böhmwinde) Jahresdurchschnittstemperatur: 4,5 Grad
Waldsterben Was wurde daraus? Beobachtungen aus dem Stift Schlägl Rahmenbedingungen Höhenlage: 600m 1378m (Plöckenstein) Hauptwindrichtung: W und N (Böhmwinde) Jahresdurchschnittstemperatur: 4,5 Grad
Das Ende der Käfersaison kündigt sich an!
 Fangsumme/ Falle (Mittelwert) 9.9.215 Das Ende der Käfersaison kündigt sich an! Beobachtungszeitraum: 2. bis 8. September 215 Die aktuelle Lage Die Temperaturen der letzten Woche bewegte sich in dem vorhergesagten
Fangsumme/ Falle (Mittelwert) 9.9.215 Das Ende der Käfersaison kündigt sich an! Beobachtungszeitraum: 2. bis 8. September 215 Die aktuelle Lage Die Temperaturen der letzten Woche bewegte sich in dem vorhergesagten
Klimawandel in Baden-Württemberg
 auch in M2 LUFTTEMPERATUR Die Jahresmitteltemperatur stieg seit 1901 bis heute von rund 8 C auf über 9 C an. Der größte Anstieg erfolgte dabei erst in den letzten 30 Jahren seit 1980. Ein Beispiel verdeutlicht
auch in M2 LUFTTEMPERATUR Die Jahresmitteltemperatur stieg seit 1901 bis heute von rund 8 C auf über 9 C an. Der größte Anstieg erfolgte dabei erst in den letzten 30 Jahren seit 1980. Ein Beispiel verdeutlicht
Wald Arbeitsblatt W6 Nationalpark Bayerischer Wald
 Aufgabe 1a: Der Klimawandel: Entstehung Die Abbildung unten zeigt wie der globale Klimawandel entsteht. Beschrifte die großen Kästchen in der Abbildung mit folgenden Begriffen richtig: Erde Sonne Weltraum
Aufgabe 1a: Der Klimawandel: Entstehung Die Abbildung unten zeigt wie der globale Klimawandel entsteht. Beschrifte die großen Kästchen in der Abbildung mit folgenden Begriffen richtig: Erde Sonne Weltraum
KLIMAATLAS REGION STUTTGART
 KLIMAATLAS REGION STUTTGART n Temperatur-Jahresmittelwerte (B2.1) Im Gebiet der beträgt die Jahresmitteltemperatur zwischen etwa 4 Grad Celsius auf den Höhen der Schwäbischen Alb und bis zu etwa 12 Grad
KLIMAATLAS REGION STUTTGART n Temperatur-Jahresmittelwerte (B2.1) Im Gebiet der beträgt die Jahresmitteltemperatur zwischen etwa 4 Grad Celsius auf den Höhen der Schwäbischen Alb und bis zu etwa 12 Grad
Entwicklungen im Nationalpark Bayerischer Wald
 NÜRTINGEN, 08. NOVEMBER 2013 Entwicklungen im Dr. Franz Leibl Lage des Nationalparks Bayerischer Wald 2 Waldinventur 2002/03 Fi 67 %, Bu 24,5 %, Vogelbeere 3,1 %, Tanne 2,6 %, Andere 2,8 % 3 Zonierung
NÜRTINGEN, 08. NOVEMBER 2013 Entwicklungen im Dr. Franz Leibl Lage des Nationalparks Bayerischer Wald 2 Waldinventur 2002/03 Fi 67 %, Bu 24,5 %, Vogelbeere 3,1 %, Tanne 2,6 %, Andere 2,8 % 3 Zonierung
Mal winterlich, mal mild, mal feucht - vor allem aber extrem wenig Sonne.
 Witterungsbericht Winter 2012 / 2013 Winter 2012 / 2013: Zwischen Winter und Winterling - mit insgesamt mehr Schnee als Schneeglöckchen Der meteorologische Winter 2012 / 2013 von Anfang Dezember bis Ende
Witterungsbericht Winter 2012 / 2013 Winter 2012 / 2013: Zwischen Winter und Winterling - mit insgesamt mehr Schnee als Schneeglöckchen Der meteorologische Winter 2012 / 2013 von Anfang Dezember bis Ende
Witterungsbericht. - Frühjahr
 Witterungsbericht - Frühjahr 2015 - Witterungsbericht Frühjahr 2015 Erstellt: Juni 2015 Thüringer Landesanstalt für Umwelt und Geologie - Thüringer Klimaagentur - Göschwitzer Str. 41 07745 Jena Email:
Witterungsbericht - Frühjahr 2015 - Witterungsbericht Frühjahr 2015 Erstellt: Juni 2015 Thüringer Landesanstalt für Umwelt und Geologie - Thüringer Klimaagentur - Göschwitzer Str. 41 07745 Jena Email:
Wald in Schutzgebieten ein Überblick
 Heino Polley / Landbauforschung - vti Agriculture and Forestry Research Sonderheft 327 2009: 75-82 75 Wald in Schutzgebieten ein Überblick von Heino Polley 1 1 Einleitung Wie viel Naturschutz braucht der
Heino Polley / Landbauforschung - vti Agriculture and Forestry Research Sonderheft 327 2009: 75-82 75 Wald in Schutzgebieten ein Überblick von Heino Polley 1 1 Einleitung Wie viel Naturschutz braucht der
Grün, grün, grün muss er sein
 tropenwald Grün, grün, grün muss er sein Foto: Greenpeace/Volker Oppermann 22 Nr. 115/4.12 wald Mut zum Nichtstun im Nationalpark Bayerischer Wald: Die toten, grauen Bäume bleiben stehen Nr. 115/4.12 23
tropenwald Grün, grün, grün muss er sein Foto: Greenpeace/Volker Oppermann 22 Nr. 115/4.12 wald Mut zum Nichtstun im Nationalpark Bayerischer Wald: Die toten, grauen Bäume bleiben stehen Nr. 115/4.12 23
Gibt es in Deutschland nur noch zu warme Monate?
 Gibt es in Deutschland nur noch zu warme Monate? Rolf Ullrich 1), Jörg Rapp 2) und Tobias Fuchs 1) 1) Deutscher Wetterdienst, Abteilung Klima und Umwelt, D-63004 Offenbach am Main 2) J.W.Goethe-Universität,
Gibt es in Deutschland nur noch zu warme Monate? Rolf Ullrich 1), Jörg Rapp 2) und Tobias Fuchs 1) 1) Deutscher Wetterdienst, Abteilung Klima und Umwelt, D-63004 Offenbach am Main 2) J.W.Goethe-Universität,
Das Niederschlagsgeschehen in Mitteleuropa in den ersten 12 Tagen des August 2002
 Das Niederschlagsgeschehen in Mitteleuropa in den ersten 12 Tagen des August 2002 In den ersten 12 Tagen des August 2002 kam es in Mitteleuropa zu verschiedenen Starkregenereignissen, die große Schäden
Das Niederschlagsgeschehen in Mitteleuropa in den ersten 12 Tagen des August 2002 In den ersten 12 Tagen des August 2002 kam es in Mitteleuropa zu verschiedenen Starkregenereignissen, die große Schäden
Faktensammlung zur Dritten Bundeswaldinventur (BWI 3) für Mecklenburg-Vorpommern
 für Mecklenburg-Vorpommern") Faktensammlung zur Dritten Bundeswaldinventur (BWI 3) für Mecklenburg-Vorpommern Erhebungsmethodik BWI Großrauminventur auf Stichprobenbasis. Ziel Erfassung der aktuellen Waldverhältnisse und Produktionsmöglichkeiten
Faktensammlung zur Dritten Bundeswaldinventur (BWI 3) für Mecklenburg-Vorpommern Erhebungsmethodik BWI Großrauminventur auf Stichprobenbasis. Ziel Erfassung der aktuellen Waldverhältnisse und Produktionsmöglichkeiten
Klimawandel in Deutschland: Was wissen wir und worauf fußen diese Erkenntnisse? Dr. Thomas Deutschländer Deutscher Wetterdienst.
 Klimawandel in Deutschland: Was wissen wir und worauf fußen diese Erkenntnisse? Dr. Thomas Deutschländer Deutscher Wetterdienst Seite 1 Entwicklung der Globaltemperatur 1850 bis 2015 von der Klimareferenzperiode
Klimawandel in Deutschland: Was wissen wir und worauf fußen diese Erkenntnisse? Dr. Thomas Deutschländer Deutscher Wetterdienst Seite 1 Entwicklung der Globaltemperatur 1850 bis 2015 von der Klimareferenzperiode
Lebenserwartung und Mortalität im Alter - ein Überblick
 Lebenserwartung und Mortalität im Alter - ein Überblick Lebenserwartung älterer Menschen Die Lebenserwartung kann als das allgemeinste Maß betrachtet werden, das über den Gesundheitszustand einer Bevölkerung
Lebenserwartung und Mortalität im Alter - ein Überblick Lebenserwartung älterer Menschen Die Lebenserwartung kann als das allgemeinste Maß betrachtet werden, das über den Gesundheitszustand einer Bevölkerung
Borkenkäfer: Der Herr der Rinde
 Dr. Beat Wermelinger Borkenkäfer: Der Herr der Rinde In den Lothar-geschädigten Gebieten der Schweiz läuft zurzeit eine Massenvermehrung des Buchdruckers in noch nie gesehenem Ausmass ab. Voraussetzungen
Dr. Beat Wermelinger Borkenkäfer: Der Herr der Rinde In den Lothar-geschädigten Gebieten der Schweiz läuft zurzeit eine Massenvermehrung des Buchdruckers in noch nie gesehenem Ausmass ab. Voraussetzungen
Phänologische Uhren der ausgewerteten Haupteinheitengruppen Hessens
 Uhren ausgewerteten Haupteinheitengruppen Hessens Haupteinheitengruppe 14 - Hessisch-Fränkisches Bergland Im Hessisch-Fränkischen Bergland scheint sich innere Ring Uhr gegen den Uhrzeigersinn verschoben
Uhren ausgewerteten Haupteinheitengruppen Hessens Haupteinheitengruppe 14 - Hessisch-Fränkisches Bergland Im Hessisch-Fränkischen Bergland scheint sich innere Ring Uhr gegen den Uhrzeigersinn verschoben
Willkommen im Nationalpark Schwarzwald
 Willkommen im Nationalpark Schwarzwald Willkommen im Nationalpark Schwarzwald Eine Spur wilder. In unserer dicht besiedelten, lauten Welt sind Nationalparke Inseln voller Ruhe und wilder Natur. Sie bewahren
Willkommen im Nationalpark Schwarzwald Willkommen im Nationalpark Schwarzwald Eine Spur wilder. In unserer dicht besiedelten, lauten Welt sind Nationalparke Inseln voller Ruhe und wilder Natur. Sie bewahren
Zusammenhang zwischen Jahrringen und Klima Berechnung, Darstellung, Interpretation
 Zusammenhang zwischen Jahrringen und Klima Berechnung, Darstellung, Interpretation Jasmin FIALA, Julia FLANDORFER, Monika FÖRSTER, Stephan FRANK, Carina HASITZKA, Patrick HÖSS, Katrin LOIBL, Viktor NAWRATA,
Zusammenhang zwischen Jahrringen und Klima Berechnung, Darstellung, Interpretation Jasmin FIALA, Julia FLANDORFER, Monika FÖRSTER, Stephan FRANK, Carina HASITZKA, Patrick HÖSS, Katrin LOIBL, Viktor NAWRATA,
Die Gradtagzahlen des Jahres 2008 und ihre Abweichungen zum Zeitraum P.-H. Voß
 Die Gradtagzahlen des Jahres 2008 und ihre Abweichungen zum Zeitraum 1981-2000 P.-H. Voß Gradtagzahlen bzw. Gradtage werden zur überschlägigen Berechnung des Heizenergieverbrauchs sowie zur Witterungsbereinigung
Die Gradtagzahlen des Jahres 2008 und ihre Abweichungen zum Zeitraum 1981-2000 P.-H. Voß Gradtagzahlen bzw. Gradtage werden zur überschlägigen Berechnung des Heizenergieverbrauchs sowie zur Witterungsbereinigung
Ergebnisse und Interpretation 54
 Ergebnisse und Interpretation 54 4 Ergebnisse In den Abbildungen 24/4.1 bis 29/4.1 werden die Laktat-Geschwindigkeits-Kurve und die Herzfrequenzwerte der beiden Schwimmgruppen (Männer: n=6, Frauen: n=8)
Ergebnisse und Interpretation 54 4 Ergebnisse In den Abbildungen 24/4.1 bis 29/4.1 werden die Laktat-Geschwindigkeits-Kurve und die Herzfrequenzwerte der beiden Schwimmgruppen (Männer: n=6, Frauen: n=8)
BERICHTE FREIBURGER FORSTLICHE FORSCHUNG
 BERICHTE FREIBURGER FORSTLICHE FORSCHUNG HEFT 3 Betriebswirtschaftliche Untersuchungen im bäuerlichen Privatwald in Baden-Württemberg Testbetriebsnetz Kleinprivatwald 5-200 ha Ergebnisse FWJ 1997 FORSTLICHE
BERICHTE FREIBURGER FORSTLICHE FORSCHUNG HEFT 3 Betriebswirtschaftliche Untersuchungen im bäuerlichen Privatwald in Baden-Württemberg Testbetriebsnetz Kleinprivatwald 5-200 ha Ergebnisse FWJ 1997 FORSTLICHE
1 NIEDERSCHLAGSMENGEN
 1 NIEDERSCHLAGSMENGEN Im Kanton Solothurn fallen im langjährigen Durchschnitt etwa 1240 mm Niederschläge pro Jahr. Das sind insgesamt rund 980 Mia. Liter Regen und Schnee oder ein 225000 km langer Zug,
1 NIEDERSCHLAGSMENGEN Im Kanton Solothurn fallen im langjährigen Durchschnitt etwa 1240 mm Niederschläge pro Jahr. Das sind insgesamt rund 980 Mia. Liter Regen und Schnee oder ein 225000 km langer Zug,
2 Die Niederschlagsverteilung für Deutschland im Jahr 2004 - Überblick
 2 Die Niederschlagsverteilung für Deutschland im Jahr 2004 - Überblick Das Hauptziel dieser Arbeit ist einen hochaufgelösten Niederschlagsdatensatz für Deutschland, getrennt nach konvektivem und stratiformem
2 Die Niederschlagsverteilung für Deutschland im Jahr 2004 - Überblick Das Hauptziel dieser Arbeit ist einen hochaufgelösten Niederschlagsdatensatz für Deutschland, getrennt nach konvektivem und stratiformem
Vor 30 Jahren: Klimasprung in Mittel- und Westeuropa- ein kleiner, kurzatmiger Hüpfer
 Vor 30 Jahren: Klimasprung in Mittel- und Westeuropa- ein kleiner, kurzatmiger Hüpfer Deutschland: Unterschiedlicher Temperaturverlauf im Winter und Sommer Betrachtet man das Temperaturverhalten, beginnend
Vor 30 Jahren: Klimasprung in Mittel- und Westeuropa- ein kleiner, kurzatmiger Hüpfer Deutschland: Unterschiedlicher Temperaturverlauf im Winter und Sommer Betrachtet man das Temperaturverhalten, beginnend
ZENTRALANSTALT FÜR METEOROLOGIE UND GEODYNAMIK. Meteorologische Analyse des Niederschlags von Juni 2009
 Meteorologische Analyse des Niederschlags von 22.-25. Juni 2009 Thomas Haiden 30. Juni 2009 Synopsis Im 72-stündigen Zeitraum von 22.6.09 bis 25.6.09 (jeweils 06 UTC) fielen in weiten Teilen der Bundesländer
Meteorologische Analyse des Niederschlags von 22.-25. Juni 2009 Thomas Haiden 30. Juni 2009 Synopsis Im 72-stündigen Zeitraum von 22.6.09 bis 25.6.09 (jeweils 06 UTC) fielen in weiten Teilen der Bundesländer
Theorie der Neuankömmlinge widerlegt Die älteste Fichte der Damit wären die schwedischen Fichten die ältesten Bäume der Welt. Welt
 Ältester Baum der Welt entdeckt Fichte in den schwedischen Bergen erreicht Alter von 9.550 Jahren Den ältesten Baum der Welt haben Wissenschaftler jetzt im schwedischen Dalarna entdeckt. Die 9.550 Jahre
Ältester Baum der Welt entdeckt Fichte in den schwedischen Bergen erreicht Alter von 9.550 Jahren Den ältesten Baum der Welt haben Wissenschaftler jetzt im schwedischen Dalarna entdeckt. Die 9.550 Jahre
Waldzustandsbericht 2006
 Waldzustandsbericht 2006 Ergebnisse bundesweit Aufnahme im 16 x 16 km- Netz Schadstufe 1 (Warnstufe) Schadstufe 2-4 32% (+3%- Punkte) 40% (- 2%- Punkte) 28% (- 1%- Punkt) Ursächlich für die nur schwache
Waldzustandsbericht 2006 Ergebnisse bundesweit Aufnahme im 16 x 16 km- Netz Schadstufe 1 (Warnstufe) Schadstufe 2-4 32% (+3%- Punkte) 40% (- 2%- Punkte) 28% (- 1%- Punkt) Ursächlich für die nur schwache
8. Statistik Beispiel Noten. Informationsbestände analysieren Statistik
 Informationsbestände analysieren Statistik 8. Statistik Nebst der Darstellung von Datenreihen bildet die Statistik eine weitere Domäne für die Auswertung von Datenbestände. Sie ist ein Fachgebiet der Mathematik
Informationsbestände analysieren Statistik 8. Statistik Nebst der Darstellung von Datenreihen bildet die Statistik eine weitere Domäne für die Auswertung von Datenbestände. Sie ist ein Fachgebiet der Mathematik
Witterungsbericht. - Sommer
 Witterungsbericht - Sommer 2013 - Witterungsbericht Sommer 2013 Erstellt: September 2013 Thüringer Landesanstalt für Umwelt und Geologie - Thüringer Klimaagentur - Göschwitzer Str. 41 07745 Jena Email:
Witterungsbericht - Sommer 2013 - Witterungsbericht Sommer 2013 Erstellt: September 2013 Thüringer Landesanstalt für Umwelt und Geologie - Thüringer Klimaagentur - Göschwitzer Str. 41 07745 Jena Email:
Naturwaldreservate als Datenbasis zur Einschätzung natürlicher Waldentwicklungen in einem künftigen Nationalpark
 Naturwaldreservate als Datenbasis zur Einschätzung natürlicher Waldentwicklungen Dr. Patricia Balcar Dr. Patricia Balcar AUSWEISUNGEN VON NATURWALDRESERVATEN IN RHEINLAND-PFALZ aus der Nutzung genommen
Naturwaldreservate als Datenbasis zur Einschätzung natürlicher Waldentwicklungen Dr. Patricia Balcar Dr. Patricia Balcar AUSWEISUNGEN VON NATURWALDRESERVATEN IN RHEINLAND-PFALZ aus der Nutzung genommen
HISTALP LANGZEITKLIMAREIHEN ÖSTERREICH SOMMERBERICHT 2013
 HISTALP LANGZEITKLIMAREIHEN ÖSTERREICH SOMMERBERICHT 2013 Der vergangene Sommer machte mit Lufttemperaturen von erstmals über 40 Grad Celsius Schlagzeilen, die ZAMG berichtete ausführlich dazu. Neben den
HISTALP LANGZEITKLIMAREIHEN ÖSTERREICH SOMMERBERICHT 2013 Der vergangene Sommer machte mit Lufttemperaturen von erstmals über 40 Grad Celsius Schlagzeilen, die ZAMG berichtete ausführlich dazu. Neben den
L a n d e s f o r s t Mecklenburg-Vorpommern - A n s t a l t d e s ö f f e n t l i c h e n R e c h t s - D e r V o r s t a n d
 1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 2011 2012 2013 2014 2015 2016 2017 L a n d e s f o r s t Mecklenburg-Vorpommern - A n s t a l t d e s ö f f e n t l i c h e n R e c
1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 2011 2012 2013 2014 2015 2016 2017 L a n d e s f o r s t Mecklenburg-Vorpommern - A n s t a l t d e s ö f f e n t l i c h e n R e c
Informationsveranstaltung Frankenwald potenzieller Nationalpark Stockheim Neukenroth, 2. Juni 2017
 Informationsveranstaltung Frankenwald potenzieller Nationalpark Stockheim Neukenroth, 2. Juni 2017 Wie viele Nationalparke in Europa: > 300 im Sinne der Vorgaben der IUCN Karte Deutschland und NP in Deutschland:
Informationsveranstaltung Frankenwald potenzieller Nationalpark Stockheim Neukenroth, 2. Juni 2017 Wie viele Nationalparke in Europa: > 300 im Sinne der Vorgaben der IUCN Karte Deutschland und NP in Deutschland:
Waldschäden durch Luftverunreinigungen
 Waldschäden durch Luftverunreinigungen Neuartige Waldschäden, in den Anfangszeiten auch Waldsterben genannt, bezeichnet Waldschadensbilder in Mittel- und Nordeuropa, die seit Mitte der 1970er Jahre festgestellt
Waldschäden durch Luftverunreinigungen Neuartige Waldschäden, in den Anfangszeiten auch Waldsterben genannt, bezeichnet Waldschadensbilder in Mittel- und Nordeuropa, die seit Mitte der 1970er Jahre festgestellt
Dr. Kai Pfannschmidt. Beobachteter K L I M A W A N D E L in Thüringen. Klimaprojektionen für Thüringen Regionale Auswertungen
 Beobachteter K L I M A W A N D E L in Thüringen Klimaprojektionen für Thüringen Regionale Auswertungen Dr. Kai Pfannschmidt Thüringer Landesanstalt für Umwelt und Geologie Referat 44 Klimaagentur 2. Jenaer
Beobachteter K L I M A W A N D E L in Thüringen Klimaprojektionen für Thüringen Regionale Auswertungen Dr. Kai Pfannschmidt Thüringer Landesanstalt für Umwelt und Geologie Referat 44 Klimaagentur 2. Jenaer
Hintergrundinformation Berlin, Entwicklung der Mitteltemperatur im Dezember in den letzten 45 Jahren
 WWF Deutschland Regine Günther WWF Vertretung Berlin Große Präsidentenstraße 10 10178 Berlin Tel.: 0 30/30 87 42-0 Fax: 0 30/30 87 42 50 rguenther@wwf.de www.wwf.de Hintergrundinformation Berlin, 21.12.2006
WWF Deutschland Regine Günther WWF Vertretung Berlin Große Präsidentenstraße 10 10178 Berlin Tel.: 0 30/30 87 42-0 Fax: 0 30/30 87 42 50 rguenther@wwf.de www.wwf.de Hintergrundinformation Berlin, 21.12.2006
DIE NIEDERSCHLAGSVERHÄLTNISSE IN DER STEIERMARK IN DEN LETZTEN 100 JAHREN Hydrographischer Dienst Steiermark
 1 DIE NIEDERSCHLAGSVERHÄLTNISSE IN DER STEIERMARK IN DEN LETZTEN 1 JAHREN Hydrographischer Dienst Steiermark 1. EINLEITUNG Das Jahr 21 war geprägt von extremer Trockenheit vor allem in den südlichen Teilen
1 DIE NIEDERSCHLAGSVERHÄLTNISSE IN DER STEIERMARK IN DEN LETZTEN 1 JAHREN Hydrographischer Dienst Steiermark 1. EINLEITUNG Das Jahr 21 war geprägt von extremer Trockenheit vor allem in den südlichen Teilen
Wenn der Wald in Stress gerät
 Wenn der Wald in Stress gerät Heimische Wälder im (Klima-) Wandel Name: 1 MODUL Baumwachstum Holzanatomie Strukturen der Baumscheiben erkunden Arbeitsblatt 1 hier wird Material 1 (Baumscheibenscans) benötigt
Wenn der Wald in Stress gerät Heimische Wälder im (Klima-) Wandel Name: 1 MODUL Baumwachstum Holzanatomie Strukturen der Baumscheiben erkunden Arbeitsblatt 1 hier wird Material 1 (Baumscheibenscans) benötigt
Höhendifferenzierung der Teilgebiete im LARSIM WHM für die Schneemodellierung
 Höhendifferenzierung der Teilgebiete im LARSIM WHM für die Schneemodellierung Natalie Stahl WWA Technische Umsetzung Hydron GmbH Inhalt A) Beschreibung des Problems B) Lösungsansatz C) Detailstudie Sensitivitätsanalyse
Höhendifferenzierung der Teilgebiete im LARSIM WHM für die Schneemodellierung Natalie Stahl WWA Technische Umsetzung Hydron GmbH Inhalt A) Beschreibung des Problems B) Lösungsansatz C) Detailstudie Sensitivitätsanalyse
5. Witterung im Winter 1998/99
 5. Witterung im Winter 1998/99 von Dr. Karl Gabl Zentralanstalt für Meteorologie und Geodynamik, Regionalstelle für Tirol und Vorarlberg Die Daten zur Beschreibung des überaus schneereichen Winters 1998/99
5. Witterung im Winter 1998/99 von Dr. Karl Gabl Zentralanstalt für Meteorologie und Geodynamik, Regionalstelle für Tirol und Vorarlberg Die Daten zur Beschreibung des überaus schneereichen Winters 1998/99
Sachsen im Klimawandel Sommer
 Sachsen im Klimawandel Sommer Die wichtigsten Fakten zu dieser Jahreszeit Zusammenfassung Die Sommer (Monate Juni, Juli, August) der Klimanormalperiode 1981-2010 wiesen im Vergleich zur Referenz 1961-1990
Sachsen im Klimawandel Sommer Die wichtigsten Fakten zu dieser Jahreszeit Zusammenfassung Die Sommer (Monate Juni, Juli, August) der Klimanormalperiode 1981-2010 wiesen im Vergleich zur Referenz 1961-1990
Digitale Demokratie: Chancen und Herausforderungen von sozialen Netzwerken. Bachelorarbeit
 Digitale Demokratie: Chancen und Herausforderungen von sozialen Netzwerken Bachelorarbeit zur Erlangung des akademischen Grades Bachelor of Science (B.Sc.) im Studiengang Wirtschaftswissenschaft der Wirtschaftswissenschaftlichen
Digitale Demokratie: Chancen und Herausforderungen von sozialen Netzwerken Bachelorarbeit zur Erlangung des akademischen Grades Bachelor of Science (B.Sc.) im Studiengang Wirtschaftswissenschaft der Wirtschaftswissenschaftlichen
Das Oltner Wetter im Februar 2010
 Das Oltner Wetter im Februar 2010 Winterlich mit Hauch von Frühling gegen Monatsende Auch der vergangene Februar war, wie schon der Januar 2010, mehrheitlich durch winterliches Wetter geprägt Diese Schlussfolgerung
Das Oltner Wetter im Februar 2010 Winterlich mit Hauch von Frühling gegen Monatsende Auch der vergangene Februar war, wie schon der Januar 2010, mehrheitlich durch winterliches Wetter geprägt Diese Schlussfolgerung
Lehrveranstaltung Aktuelle Probleme des Forstschutzes
 Lehrveranstaltung Aktuelle Probleme des Forstschutzes Beitrag: Zur Erfassung der Auswirkungen von Schadereignissen im Wald Schwerpunkt: Methoden der Fernerkundung Institut für Waldinventur und Waldwachstum,
Lehrveranstaltung Aktuelle Probleme des Forstschutzes Beitrag: Zur Erfassung der Auswirkungen von Schadereignissen im Wald Schwerpunkt: Methoden der Fernerkundung Institut für Waldinventur und Waldwachstum,
KLIMAATLAS REGION STUTTGART
 KLIMAATLAS REGION STUTTGART n Temperatur-Prognose 271-2 (C 1.1) Die Klimaprognosen des Weltklimarates (IPCC) gehen global von einer Temperaturerhöhung in diesem Jahrhundert von etwa 1,4 bis 4, Grad Celsius
KLIMAATLAS REGION STUTTGART n Temperatur-Prognose 271-2 (C 1.1) Die Klimaprognosen des Weltklimarates (IPCC) gehen global von einer Temperaturerhöhung in diesem Jahrhundert von etwa 1,4 bis 4, Grad Celsius
Beobachtungen zum Einfluss der einheimischen Begleitvegetation auf die Entwicklung von jungen Ginkgo biloba Pflanzen
 Beobachtungen zum Einfluss der einheimischen Begleitvegetation auf die Entwicklung von jungen Ginkgo biloba Pflanzen Beobachtet auf dem Grundstück Mollesnejta(~2800m ü. NN) im Tal von Cochabamba, Bolivien
Beobachtungen zum Einfluss der einheimischen Begleitvegetation auf die Entwicklung von jungen Ginkgo biloba Pflanzen Beobachtet auf dem Grundstück Mollesnejta(~2800m ü. NN) im Tal von Cochabamba, Bolivien
Verhaltener Flug, stetige Entwicklung
 Borkenkäfer-Newsletter Nordschwarzwald 19.05.2016 Verhaltener Flug, stetige Entwicklung Nachdem gestern vereinzelt die 16,5 C erreicht wurden, liegen die Tempearturen heute wieder darunter. Mit örtlich
Borkenkäfer-Newsletter Nordschwarzwald 19.05.2016 Verhaltener Flug, stetige Entwicklung Nachdem gestern vereinzelt die 16,5 C erreicht wurden, liegen die Tempearturen heute wieder darunter. Mit örtlich
Pressekonferenz. Thema: Vorstellung des Geburtenbarometers - Eine neue Methode zur Messung der Geburtenentwicklung
 Pressekonferenz mit Bundesministerin Ursula Haubner, Bundesministerium für soziale Sicherheit, Generationen und Konsumentenschutz und Prof. Dr. Wolfgang Lutz, Direktor des Instituts für Demographie der
Pressekonferenz mit Bundesministerin Ursula Haubner, Bundesministerium für soziale Sicherheit, Generationen und Konsumentenschutz und Prof. Dr. Wolfgang Lutz, Direktor des Instituts für Demographie der
2. Witterung im Winter 1996/97
 2. Witterung im Winter 1996/97 von Dr. Karl Gabl, Mag. Gabriele Mascher-Schlieter ZAMG-Wetterdienststelle Innsbruck Die Beobachtungen und Messungen der Klimastationen der Zentralanstalt für Meteorologie
2. Witterung im Winter 1996/97 von Dr. Karl Gabl, Mag. Gabriele Mascher-Schlieter ZAMG-Wetterdienststelle Innsbruck Die Beobachtungen und Messungen der Klimastationen der Zentralanstalt für Meteorologie
NATURWALDRESERVAT HECKE
 Amt für Ernährung, Landwirtschaft und Forsten Passau-Rotthalmünster NATURWALDRESERVAT HECKE Naturwaldreservat Hecke Gräben, Totholz und junge Bäume vermitteln den Besuchern einen urwaldartigen Eindruck.
Amt für Ernährung, Landwirtschaft und Forsten Passau-Rotthalmünster NATURWALDRESERVAT HECKE Naturwaldreservat Hecke Gräben, Totholz und junge Bäume vermitteln den Besuchern einen urwaldartigen Eindruck.
Forstdienst. Käfersituation in Salzburg
 Käfersituation in Salzburg Winterschäden Winter 2014/2015: Mild mit relativ wenig Schnee bis in höhere Lagen Regenreicher Jänner, trockener Februar Nasser, schwerer Schnee bis in höhere Lagen Relativ wenig
Käfersituation in Salzburg Winterschäden Winter 2014/2015: Mild mit relativ wenig Schnee bis in höhere Lagen Regenreicher Jänner, trockener Februar Nasser, schwerer Schnee bis in höhere Lagen Relativ wenig
HISTALP LANGZEITKLIMAREIHEN ÖSTERREICH WINTERBERICHT 2017/18
 HISTALP LANGZEITKLIMAREIHEN ÖSTERREICH WINTERBERICHT 2017/18 Über das gesamte österreichische Tiefland gerechnet war der Winter 2017/18 im Vergleich zum Mittel 1961-1990 um 1,4 C zu warm, er verfehlt damit
HISTALP LANGZEITKLIMAREIHEN ÖSTERREICH WINTERBERICHT 2017/18 Über das gesamte österreichische Tiefland gerechnet war der Winter 2017/18 im Vergleich zum Mittel 1961-1990 um 1,4 C zu warm, er verfehlt damit
7 KRONENZUSTAND 2011 ANHANG
 ANHANG 7 KRONENZUSTAND 211 a a Der Kronenzustand der Waldbäume hat sich in diesem gegenüber 21 insgesamt leicht verbessert. Der mittlere Nadel- und Blattverlust sank um,3 Prozentpunkte auf 19,8 Prozent.
ANHANG 7 KRONENZUSTAND 211 a a Der Kronenzustand der Waldbäume hat sich in diesem gegenüber 21 insgesamt leicht verbessert. Der mittlere Nadel- und Blattverlust sank um,3 Prozentpunkte auf 19,8 Prozent.
NATURWALDRESERVAT KREBSWIESE- LANGERJERGEN
 Amt für Ernährung, Landwirtschaft und Forsten Mindelheim NATURWALDRESERVAT KREBSWIESE- LANGERJERGEN Naturwaldreservat Krebswiese- Langerjergen Buchenwald mit Totholz ein Element naturnaher Wälder. ALLGEMEINES
Amt für Ernährung, Landwirtschaft und Forsten Mindelheim NATURWALDRESERVAT KREBSWIESE- LANGERJERGEN Naturwaldreservat Krebswiese- Langerjergen Buchenwald mit Totholz ein Element naturnaher Wälder. ALLGEMEINES
Beschreibung von Diagrammen, Tabellen und Grafiken zur Prüfungsvorbereitung auf das Zertifikat Deutsch
 Beschreibung von Diagrammen, Tabellen und Grafiken zur Prüfungsvorbereitung auf das Zertifikat Deutsch Einleitung: Was ist der Titel oder Inhalt der Grafik? Das Diagramm zeigt... Die Grafik stellt... dar.
Beschreibung von Diagrammen, Tabellen und Grafiken zur Prüfungsvorbereitung auf das Zertifikat Deutsch Einleitung: Was ist der Titel oder Inhalt der Grafik? Das Diagramm zeigt... Die Grafik stellt... dar.
Klimawandel im Offenland und Wald
 Klimawandel im Offenland und Wald Klimawandel Einleitung Deutschland Thüringen und Rhön Ursachen des Klimawandels Ein anthropogener Einfluss wird als gesichert angesehen, da sich der rezente Temperaturanstieg
Klimawandel im Offenland und Wald Klimawandel Einleitung Deutschland Thüringen und Rhön Ursachen des Klimawandels Ein anthropogener Einfluss wird als gesichert angesehen, da sich der rezente Temperaturanstieg
Die Bilanz der Bayerischen Staatsforsten Geschäftsjahr Dr. Rudolf Freidhager, Reinhardt Neft 19. Oktober 2012
 Die Bilanz der Bayerischen Staatsforsten Geschäftsjahr 2012 Dr. Rudolf Freidhager, Reinhardt Neft 19. Oktober 2012 Die Bilanz der Bayerischen Staatsforsten Geschäftsjahr 2012 Geschäftsjahr 2012 - Forstliche
Die Bilanz der Bayerischen Staatsforsten Geschäftsjahr 2012 Dr. Rudolf Freidhager, Reinhardt Neft 19. Oktober 2012 Die Bilanz der Bayerischen Staatsforsten Geschäftsjahr 2012 Geschäftsjahr 2012 - Forstliche
STAR: Entwicklung der Strukturen und Beschäftigtenzahlen in Rechtsanwaltskanzleien
 Quelle: BRAK-Mitteilungen 4/2000 (S. 166-169) Seite 1 STAR: Entwicklung der Strukturen und Beschäftigtenzahlen in Rechtsanwaltskanzleien Alexandra Schmucker, Institut für Freie Berufe, Nürnberg In nachstehendem
Quelle: BRAK-Mitteilungen 4/2000 (S. 166-169) Seite 1 STAR: Entwicklung der Strukturen und Beschäftigtenzahlen in Rechtsanwaltskanzleien Alexandra Schmucker, Institut für Freie Berufe, Nürnberg In nachstehendem
ADAGIO - FRAGEBOGEN (anonym)
") ADAGIO - FRAGEBOGEN (anonym) ADAGIO ist ein EU-projekt unter Leitung der Universität für Bodenkultur zum Thema : Anpassung Der LAndwirtschaft in gefährdeten Europäischen ReGIOnen an den Klimawandel Mit
ADAGIO - FRAGEBOGEN (anonym) ADAGIO ist ein EU-projekt unter Leitung der Universität für Bodenkultur zum Thema : Anpassung Der LAndwirtschaft in gefährdeten Europäischen ReGIOnen an den Klimawandel Mit
Wie lange bleiben Baumstöcke dem Ökosystem Wald erhalten?
 GErHarD NIESE Wie lange bleiben Baumstöcke dem Ökosystem Wald erhalten? Wie lange verbleiben die mit dem Boden verbundenen Reste der gefällten Bäume im Bestand? Welche Bedeutung haben Baumart und Seehöhe?
GErHarD NIESE Wie lange bleiben Baumstöcke dem Ökosystem Wald erhalten? Wie lange verbleiben die mit dem Boden verbundenen Reste der gefällten Bäume im Bestand? Welche Bedeutung haben Baumart und Seehöhe?
Erholung zwischen toten Bäumen. Dr. Klaus Wagner und Prof. Dr. Michael Suda
 Erholung zwischen toten Bäumen Dr. Klaus Wagner und Prof. Dr. Michael Suda Jährliche Zunahme der Totholz- flächen im NP Bayerischer Wald Quelle: Berichte aus dem Nationalpark Heft 2/2003: Hochlageninventur
Erholung zwischen toten Bäumen Dr. Klaus Wagner und Prof. Dr. Michael Suda Jährliche Zunahme der Totholz- flächen im NP Bayerischer Wald Quelle: Berichte aus dem Nationalpark Heft 2/2003: Hochlageninventur
4.3. Körperkondition zum Zeitpunkt der Wiederbelegung
 Tabelle 28: Vergleich der maximalen Progesteronwerte der Gruppen 4 bis 6 Maßzahl Gruppe 4 BCS - Differenz 0,00 Gruppe 5 BCS - Differenz < 0,75 Gruppe 6 BCS - Differenz 0,75 Anzahl der Tiere 33 114 52 Mittelwert
Tabelle 28: Vergleich der maximalen Progesteronwerte der Gruppen 4 bis 6 Maßzahl Gruppe 4 BCS - Differenz 0,00 Gruppe 5 BCS - Differenz < 0,75 Gruppe 6 BCS - Differenz 0,75 Anzahl der Tiere 33 114 52 Mittelwert
Wie geht man mit gefährdeten Fichtenbeständen um?
 Wie geht man mit gefährdeten Fichtenbeständen um? Thomas Ledermann und Georg Kindermann Institut für Waldwachstum und Waldbau BFW-Praxistag 2017 Wege zum klimafitten Wald Wien, Gmunden, Ossiach, Innsbruck
Wie geht man mit gefährdeten Fichtenbeständen um? Thomas Ledermann und Georg Kindermann Institut für Waldwachstum und Waldbau BFW-Praxistag 2017 Wege zum klimafitten Wald Wien, Gmunden, Ossiach, Innsbruck
WALDSCHUTZ INFO. Borkenkäfer an der Weißtanne akut 2/2017. Borkenkäfer an Weißtanne
 WALDSCHUTZ INFO 2/2017 Borkenkäfer an der Weißtanne akut Aus vielen Landesteilen Baden-Württembergs kommen aktuell Meldungen über teils gravierenden Borkenkäferbefall an der Weißtanne (Abb. 1). Vor allem
WALDSCHUTZ INFO 2/2017 Borkenkäfer an der Weißtanne akut Aus vielen Landesteilen Baden-Württembergs kommen aktuell Meldungen über teils gravierenden Borkenkäferbefall an der Weißtanne (Abb. 1). Vor allem
HISTALP LANGZEITKLIMAREIHEN ÖSTERREICH SOMMERBERICHT 2015
 HISTALP LANGZEITKLIMAREIHEN ÖSTERREICH SOMMERBERICHT 2015 Der Sommer 2015 war sowohl im Tiefland als auch auf Österreichs Bergen der zweitwärmste seit Beginn der Temperaturmessungen, lediglich im Jahr
HISTALP LANGZEITKLIMAREIHEN ÖSTERREICH SOMMERBERICHT 2015 Der Sommer 2015 war sowohl im Tiefland als auch auf Österreichs Bergen der zweitwärmste seit Beginn der Temperaturmessungen, lediglich im Jahr
/ 4. AUSGABE BORKENKÄFER, FICHTEN UND COMPUTER. Lorenz Fahse und Marco Heurich
 FORSCHEN FÜR DIE UMWELT / 4. AUSGABE BORKENKÄFER, FICHTEN UND COMPUTER Lorenz Fahse und Marco Heurich 11 BORKENKÄFER, FICHTEN UND COMPUTER Lorenz Fahse und Marco Heurich Autoren: Lorenz Fahse, Dr. rer.
FORSCHEN FÜR DIE UMWELT / 4. AUSGABE BORKENKÄFER, FICHTEN UND COMPUTER Lorenz Fahse und Marco Heurich 11 BORKENKÄFER, FICHTEN UND COMPUTER Lorenz Fahse und Marco Heurich Autoren: Lorenz Fahse, Dr. rer.
STAR: Kostenstrukturen in Anwaltskanzleien 1994 und 1998
 Quelle: BRAK-Mitteilungen 2/2001 (S. 62-65) Seite 1 STAR: Kostenstrukturen in Anwaltskanzleien 1994 und 1998 Alexandra Schmucker, Institut für Freie Berufe, Nürnberg Im Rahmen der STAR-Befragung wurden
Quelle: BRAK-Mitteilungen 2/2001 (S. 62-65) Seite 1 STAR: Kostenstrukturen in Anwaltskanzleien 1994 und 1998 Alexandra Schmucker, Institut für Freie Berufe, Nürnberg Im Rahmen der STAR-Befragung wurden
Wald und Biodiversität in der Sicht des staatlichen Naturschutzes
 Eine Zukunftsaufgabe in guten Händen NW-FVA-Symposium Forstwirtschaft im Spannungsfeld vielfältiger Ansprüche Wald und Biodiversität in der Sicht des staatlichen Naturschutzes Dr. Manfred Klein Biodiversität
Eine Zukunftsaufgabe in guten Händen NW-FVA-Symposium Forstwirtschaft im Spannungsfeld vielfältiger Ansprüche Wald und Biodiversität in der Sicht des staatlichen Naturschutzes Dr. Manfred Klein Biodiversität
Bannwald "Wilder See - Hornisgrinde"
 BERICHTE FREIBURGER FORSTLICHE FORSCHUNG HEFT 30 Bannwald "Wilder See - Hornisgrinde" Forstbezirk Schönmünzach Wuchsgebiet Schwarzwald Einzelwuchsbezirk 3/05 Hornisgrinde-Murg-Schwarzwald ERLÄUTERUNGEN
BERICHTE FREIBURGER FORSTLICHE FORSCHUNG HEFT 30 Bannwald "Wilder See - Hornisgrinde" Forstbezirk Schönmünzach Wuchsgebiet Schwarzwald Einzelwuchsbezirk 3/05 Hornisgrinde-Murg-Schwarzwald ERLÄUTERUNGEN
Waldnaturschutz. Präsident Sepp Spann Bayerischer Waldbesitzerverband e.v.
 Waldnaturschutz Position des Bayerischen Waldesitzerverbandes 10. Bayerischer Waldbesitzertag, 17.09.2015 Präsident Sepp Spann Bayerischer Waldbesitzerverband e.v. Ausgangslage Wälder sind über Jahrhunderte
Waldnaturschutz Position des Bayerischen Waldesitzerverbandes 10. Bayerischer Waldbesitzertag, 17.09.2015 Präsident Sepp Spann Bayerischer Waldbesitzerverband e.v. Ausgangslage Wälder sind über Jahrhunderte
2. Witterung im Winter 1995/96
 2. Witterung im Winter 1995/96 von Dr. Karl Gabl, ZAMG Wetterdienststelle Innsbruck Die Beobachtungen und Messungen der Klimastationen der Zentralanstalt für Meteorologie und Geodynamik, Regionalstelle
2. Witterung im Winter 1995/96 von Dr. Karl Gabl, ZAMG Wetterdienststelle Innsbruck Die Beobachtungen und Messungen der Klimastationen der Zentralanstalt für Meteorologie und Geodynamik, Regionalstelle
Eignung des europaweiten Copernicus Dienstes für die Erkennung von Waldflächen
 Eignung des europaweiten Copernicus Dienstes für die Erkennung von Waldflächen Überprüfung des Thüringer Verfahrens der Waldflächenerkennung im Einzugsgebiet des Forstamtes Finsterbergen Referent: Constantin
Eignung des europaweiten Copernicus Dienstes für die Erkennung von Waldflächen Überprüfung des Thüringer Verfahrens der Waldflächenerkennung im Einzugsgebiet des Forstamtes Finsterbergen Referent: Constantin
Nachhaltige Waldwirtschaft aus Sicht der Naturschutzverbände
 Nachhaltige Waldwirtschaft aus Sicht der Naturschutzverbände 90 % naturnaher Wirtschaftswald 10 % Urwälder (von morgen) Florian Schöne Waldverteilung heute Ca. 31 % der Landfläche Walz et al. 2013 PNV
Nachhaltige Waldwirtschaft aus Sicht der Naturschutzverbände 90 % naturnaher Wirtschaftswald 10 % Urwälder (von morgen) Florian Schöne Waldverteilung heute Ca. 31 % der Landfläche Walz et al. 2013 PNV
3 Holzvorräte in Bayerns Wäldern
 3 Holzvorräte in Bayerns Wäldern 3.1 Internationaler Vergleich Deutschland hat unter allen europäischen Ländern abgesehen von Russland die größten Holzvorräte (Abb.1). Die Voraussetzungen für die Forst-
3 Holzvorräte in Bayerns Wäldern 3.1 Internationaler Vergleich Deutschland hat unter allen europäischen Ländern abgesehen von Russland die größten Holzvorräte (Abb.1). Die Voraussetzungen für die Forst-
Die erhobenen Daten werden zunächst in einer Urliste angeschrieben. Daraus ermittelt man:
 Die erhobenen Daten werden zunächst in einer Urliste angeschrieben. Daraus ermittelt man: a) Die absoluten Häufigkeit: Sie gibt an, wie oft ein Variablenwert vorkommt b) Die relative Häufigkeit: Sie erhält
Die erhobenen Daten werden zunächst in einer Urliste angeschrieben. Daraus ermittelt man: a) Die absoluten Häufigkeit: Sie gibt an, wie oft ein Variablenwert vorkommt b) Die relative Häufigkeit: Sie erhält
+++ StMELF aktuell +++ StMELF aktuell +++
 Bayerisches Sttsministerium für Ernährung, Landwirtschaft und Forsten Sttsminister Helmut Brunner informiert Ergebnisse der Kronenzustandserhebung 12 in Bayern Dezember 12 +++ StMELF aktuell +++ StMELF
Bayerisches Sttsministerium für Ernährung, Landwirtschaft und Forsten Sttsminister Helmut Brunner informiert Ergebnisse der Kronenzustandserhebung 12 in Bayern Dezember 12 +++ StMELF aktuell +++ StMELF
Kartendienst zur Luftschadstoffbelastung in Deutschland
 Kartendienst zur Luftschadstoffbelastung in Deutschland Eine interaktive Darstellung der räumlichen Verteilung von Feinstaub, Ozon und Stickstoffdioxid Informationen zur Handhabung I. Datenaufbereitung
Kartendienst zur Luftschadstoffbelastung in Deutschland Eine interaktive Darstellung der räumlichen Verteilung von Feinstaub, Ozon und Stickstoffdioxid Informationen zur Handhabung I. Datenaufbereitung
Klimabündnisschwerpunktregion Bruck/L Hainburg - Schwechat. Klimadaten
 Klimabündnisschwerpunktregion Bruck/L Hainburg - Schwechat Klimadaten 1. Allgemeine Beschreibung des Untersuchungsgebietes Das Untersuchungsgebiet erstreckt sich von Schwechat bis Hainburg und wird nördlich
Klimabündnisschwerpunktregion Bruck/L Hainburg - Schwechat Klimadaten 1. Allgemeine Beschreibung des Untersuchungsgebietes Das Untersuchungsgebiet erstreckt sich von Schwechat bis Hainburg und wird nördlich
11. Symposium Energieinnovation. Die Wasserkraftnutzung in Österreich bei Klimaänderungen
 Wasserkraft bei Klimaänderung 1 11. Symposium Energieinnovation Die Wasserkraftnutzung in Österreich bei Klimaänderungen, P. Stanzel Institut für Wasserwirtschaft, Hydrologie und konstruktiven Wasserbau
Wasserkraft bei Klimaänderung 1 11. Symposium Energieinnovation Die Wasserkraftnutzung in Österreich bei Klimaänderungen, P. Stanzel Institut für Wasserwirtschaft, Hydrologie und konstruktiven Wasserbau
Landesbetrieb Wald und Holz Nordrhein-Westfalen Klimawandel verändert Wälder - auch im Teutoburger Wald -
 Klimawandel verändert Wälder - auch im Teutoburger Wald - Norbert Asche, LB WuH NRW Gelsenkirchen 1 Klimaänderung in vergleich der Mitteltemperaturen und mittlerer Niederschläge Periode 1931 1961 mit Periode
Klimawandel verändert Wälder - auch im Teutoburger Wald - Norbert Asche, LB WuH NRW Gelsenkirchen 1 Klimaänderung in vergleich der Mitteltemperaturen und mittlerer Niederschläge Periode 1931 1961 mit Periode
Veränderungen in ausgewählten Verhaltensweisen bei brünstigen Kühen in der Milchproduktion
 Veränderungen in ausgewählten Verhaltensweisen bei brünstigen Kühen in der Milchproduktion Berit Füllner und Heiko Scholz, Hochschule Anhalt, Fachbereich LOEL Die Fruchtbarkeit der Milchkühe wird durch
Veränderungen in ausgewählten Verhaltensweisen bei brünstigen Kühen in der Milchproduktion Berit Füllner und Heiko Scholz, Hochschule Anhalt, Fachbereich LOEL Die Fruchtbarkeit der Milchkühe wird durch
Der Orkan Lothar (26.12.1999) Zehn Jahre danach
 Zehn Jahre danach") Der Orkan Lothar (26.12.1999) Zehn Jahre danach Folie 2 Meteorologischer Ablauf Entstehung eines Sturmtiefs über dem Nordatlantik am 25. Dezember 1999 Rapider Druckabfall innerhalb weniger Stunden Zugbahn
Der Orkan Lothar (26.12.1999) Zehn Jahre danach Folie 2 Meteorologischer Ablauf Entstehung eines Sturmtiefs über dem Nordatlantik am 25. Dezember 1999 Rapider Druckabfall innerhalb weniger Stunden Zugbahn
Witterungsbericht. - Frühjahr
 Witterungsbericht - Frühjahr 2016 - Witterungsbericht Frühjahr 2016 Erstellt: Juni 2016 Thüringer Landesanstalt für Umwelt und Geologie - Thüringer Klimaagentur - Göschwitzer Str. 41 07745 Jena Email:
Witterungsbericht - Frühjahr 2016 - Witterungsbericht Frühjahr 2016 Erstellt: Juni 2016 Thüringer Landesanstalt für Umwelt und Geologie - Thüringer Klimaagentur - Göschwitzer Str. 41 07745 Jena Email:
Die Bilanz der Bayerischen Staatsforsten Geschäftsjahr Dr. Rudolf Freidhager, Reinhardt Neft 10. Oktober 2014
 Die Bilanz der Bayerischen Staatsforsten Geschäftsjahr 2014 Dr. Rudolf Freidhager, Reinhardt Neft 10. Oktober 2014 Die Bilanz der Bayerischen Staatsforsten Geschäftsjahr 2014 Geschäftsjahr 2014 Forstliche
Die Bilanz der Bayerischen Staatsforsten Geschäftsjahr 2014 Dr. Rudolf Freidhager, Reinhardt Neft 10. Oktober 2014 Die Bilanz der Bayerischen Staatsforsten Geschäftsjahr 2014 Geschäftsjahr 2014 Forstliche
Merkblatt 14. der Bayerischen Landesanstalt für Wald und Forstwirtschaft. Buchdrucker und Kupferstecher an Fichte
 Merkblatt 14 der Bayerischen Landesanstalt für Wald und Forstwirtschaft November 201 Buchdrucker und Kupferstecher an Fichte Ereignisse wie Windwürfe, Schneebruch oder Trockenheit erhöhen das Angebot an
Merkblatt 14 der Bayerischen Landesanstalt für Wald und Forstwirtschaft November 201 Buchdrucker und Kupferstecher an Fichte Ereignisse wie Windwürfe, Schneebruch oder Trockenheit erhöhen das Angebot an
Wolfgang Batrla. Münchberg, im Februar 2006
 Die Auswirkungen der EU-Osterweiterung und der Einführung der Lkw-Maut auf das Verkehrsaufkommen des schweren Güterverkehrs auf der B 303 zwischen Schirnding und der A9 Wolfgang Batrla Münchberg, im Februar
Die Auswirkungen der EU-Osterweiterung und der Einführung der Lkw-Maut auf das Verkehrsaufkommen des schweren Güterverkehrs auf der B 303 zwischen Schirnding und der A9 Wolfgang Batrla Münchberg, im Februar
Erwerbsbeteiligung und Arbeitslosigkeit im höheren Erwerbsalter ein statistischer Überblick
 Erwerbsbeteiligung und Arbeitslosigkeit im höheren Erwerbsalter ein statistischer Überblick Menschen im höheren Erwerbsalter sind europaweit ein bislang unzureichend genutztes Arbeitskräftepotenzial. Ihre
Erwerbsbeteiligung und Arbeitslosigkeit im höheren Erwerbsalter ein statistischer Überblick Menschen im höheren Erwerbsalter sind europaweit ein bislang unzureichend genutztes Arbeitskräftepotenzial. Ihre
Studie Risikobewusstsein Hamburger Bürger für Naturkatastrophen 2010
 Prof. Dr. Beate M.W. Ratter Institut für Küstenforschung Sozioökonomie des Küstenraumes (KSO) GKSS-Forschungszentrum Max-Planck-Str.1 21502 Geesthacht Studie Risikobewusstsein Hamburger Bürger für Naturkatastrophen
Prof. Dr. Beate M.W. Ratter Institut für Küstenforschung Sozioökonomie des Küstenraumes (KSO) GKSS-Forschungszentrum Max-Planck-Str.1 21502 Geesthacht Studie Risikobewusstsein Hamburger Bürger für Naturkatastrophen
Borkenkäfer-Monitoring im Umfeld des NP Bayerischer Wald
 Borkenkäfer-Monitoring im Umfeld des NP Bayerischer Wald Die Auswirkungen von Schutzgebieten auf benachbarte Wirtschaftswälder werden kontrovers diskutiert. Im Umfeld des Nationalparks Bayerischer Wald
Borkenkäfer-Monitoring im Umfeld des NP Bayerischer Wald Die Auswirkungen von Schutzgebieten auf benachbarte Wirtschaftswälder werden kontrovers diskutiert. Im Umfeld des Nationalparks Bayerischer Wald
Aktuelles zum Wettergeschehen
 Aktuelles zum Wettergeschehen 16. Juni 2008 / Stephan Bader Die Schafskälte im Juni - eine verschwundene Singularität Der Monat Juni zeigte sich in den letzten Jahren oft von seiner hochsommerlichen Seite.
Aktuelles zum Wettergeschehen 16. Juni 2008 / Stephan Bader Die Schafskälte im Juni - eine verschwundene Singularität Der Monat Juni zeigte sich in den letzten Jahren oft von seiner hochsommerlichen Seite.
Bekämpfungsmaßnahmen und Holzlagerung
 Bekämpfungsmaßnahmen und Holzlagerung DI Bernhard Perny Institut für Waldschutz BFW-Praxistage Fast Ort 28. Jänner 2010 Übersicht Bekämpfungsmaßnahmen im Wald Allgemein Schwerpunkt Borkenkäfer Holzlagerung
Bekämpfungsmaßnahmen und Holzlagerung DI Bernhard Perny Institut für Waldschutz BFW-Praxistage Fast Ort 28. Jänner 2010 Übersicht Bekämpfungsmaßnahmen im Wald Allgemein Schwerpunkt Borkenkäfer Holzlagerung
Erwartungen junger Führungskräfte und wie Selbstständige und Arbeitgeber darüber denken Zentrale Ergebnisse
 Erwartungen junger Führungskräfte und wie Selbstständige und Arbeitgeber darüber denken Zentrale Ergebnisse Steinbeis-Studie in Kooperation mit den Zentrale Ergebnisse der Steinbeis-Studie in Kooperation
Erwartungen junger Führungskräfte und wie Selbstständige und Arbeitgeber darüber denken Zentrale Ergebnisse Steinbeis-Studie in Kooperation mit den Zentrale Ergebnisse der Steinbeis-Studie in Kooperation
Wege und Hürden der nachhaltigen Intensivierung im Forstbetrieb. Forstökonomische Tagung Forstliche Ausbildungsstätte Pichl Norbert Putzgruber
 22 11 2012 Wege und Hürden der nachhaltigen Intensivierung im Forstbetrieb Forstökonomische Tagung Forstliche Ausbildungsstätte Pichl Norbert Putzgruber Inhalt 1. Grundlagen bei der ÖBf AG 2. Wege einer
22 11 2012 Wege und Hürden der nachhaltigen Intensivierung im Forstbetrieb Forstökonomische Tagung Forstliche Ausbildungsstätte Pichl Norbert Putzgruber Inhalt 1. Grundlagen bei der ÖBf AG 2. Wege einer