Pathomorphologische Charakterisierung der neuen hypotrichen Mausmutante sht/sht
|
|
|
- Carin Wagner
- vor 7 Jahren
- Abrufe
Transkript
1 Aus dem Institut für Pathologie der Tierärztlichen Hochschule Hannover Pathomorphologische Charakterisierung der neuen hypotrichen Mausmutante sht/sht I N A U G U R A L - D I S S E R T A T I O N zur Erlangung des Grades eines D O C T O R M E D I C I N A E V E T E R I N A R I A E durch die Tierärztliche Hochschule Hannover Vorgelegt von Jörg Ehrhardt aus Münster (Westfalen) Hannover 1997
2 Wissenschaftliche Betreuung: Univ.-Prof. Dr. S. Ueberschär 1. Gutachter: Univ.-Prof. Dr.S. Ueberschär 2. Gutachter: Univ.-Prof. Dr. H.-J. Hedrich Tag der mündlichen Prüfung:
3 Meiner Mutter und meinem Vater
4
5 INHALTSVERZEICHNIS Seite 1 Einleitung 9 2 Schrifttum Die Behaarung der Maus Embryologie der Haut und der Haarfollikel Anordnung der Haarfollikel in der Haut Embryologie und Morphologie der Adnexe Morphologie des Haarfollikels Ultrastruktur der Haarfollikelzellen Matrixzellen des Bulbus Haarbildende Zellen Zellen der Wurzelscheiden Keratine des Haarfollikels und des Haares Morphologie des Haares Haararten der Maus Haarzyklus Haarwechsel Regulation des Haarwachstums Systemische und intrinsische Haarzyklusregulation Aktuelle Modelle der Haarzyklusregulation Behaarungsdefekte der Maus Mausmodelle Genetisch determinierte Behaarungsdefekte der Maus Ausprägung der Defekte Ursachen von Haarwachstumsstörungen Beteiligung der Adnexe an Haarwachstumsstörungen Hypotrichosen des Menschen Alopezien 54
6 3 Eigene Untersuchungen Material und Methoden Tiermaterial Haltungsbedingungen Untersuchungsmethoden Umfang der Untersuchungen Hautproben Lichtmikroskopische Untersuchungen Elekronenmikroskopische Untersuchungen Haarproben Trichogramme Rasterelektonenmikroskopische Untersuchungen Organproben Voruntersuchungen an Tieren der Ursprunglinie Auswertung der Voruntersuchungen Ergebnisse Befunde an den Organen Statistische Auswertung der erfaßten Körpermaße Ophthalmologische Veränderungen Befunde am Haarkleid Behaarung des Rumpfes Morphologie der Haare Maße der Haare Verteilung der Haarsubtypen des Rumpfes Vibrissen Schwanzbehaarung Befunde an der Haut und den Haarfollikeln Veränderungen der Haut des sht/sht Genotyps Morphologie der Haarfollikel des sht/sht Genotyps Bestimmung des Zyklusstandes der Haarfollikel Dichte der Haarfollikel Morphologie der Adnexe 124
7 4 Diskussion Zusammenfassung / Summary Literaturverzeichnis Anhang Fixierlösungen % ige neutral gepufferte Formaldehydlösung %ige Glutaraldehydlösung in 0,1 M Cacodylatpuffer Färbungen Färbung mit Hämalaun-Eosin 160
8 Abkürzungsverzeichnis D DP E HDA 32 HE He Hu IF IRS Grp KAP M ORS R S sht/sht +/sht Dermis Dermale Papille Epidermis homozygote Mäuse der Ursprungslinie Färbung mit Hämatoxilin-Eosin Henlesche Schicht der inneren Wurzelscheide Huxleysche Schicht der inneren Wurzelscheide Intermediärfilament innere Wurzelscheide Grupppe Keratin-assoziiertes Protein Haarmark äußere Wurzelscheide Haarrinde Subkutis homozygoter, minderbehaarter Merkmalsträger heterozygoter, normal behaarter Merkmalsträger männlich weiblich zunehmend abnehmend
9 Einleitung 9 1 EINLEITUNG Bei Mensch und Tier sind zahlreiche Erkrankungen bekannt, die mit Hypotrichose bzw. Alopezie einhergehen. Ihre Ätiologie und Pathogenese sind bis heute zumeist weitgehend ungeklärt, obwohl vor allem in der Humanmedizin ein großes Interesse an der Therapie dieser Erkrankungen besteht. Desweiteren sind Haarfollikel die einzigen Organstrukturen bei Menschen und Tieren, die sich während des ganzen Lebens regelmäßig zyklisch wiederkehrend regenerieren. Daher sind die Physiologie und Anatomie der Haarfollikel sowohl für die Erforschung von epithelialmesenchymalen Interaktionen als auch von zyklischen Gewebedifferenzierungen von großer Bedeutung. Für die Untersuchung zahlreicher mit Minderbehaarungen einhergehender erblicher und erworbener Erkrankungen bei Menschen und Tieren bieten sich Mäuse aus verschiedenen Gründen besonders als Modelltiere an: Ihre geringe Größe und die einfache Haltung läßt eine leichte und mit nur geringen Kosten verbundene Reproduktion zu. Mäuse-Inzuchtlinien bieten zudem den Vorteil eines homogenen Erbgutes, so daß eine geringere Anzahl von Versuchstieren für reproduzierbare Untersuchungsergebnisse benötigt wird. Zum anderen ist das Mausgenom weitgehend bekannt und weist eine große Übereinstimmung mit dem des Menschen auf. Mausmodelle sind daher sowohl zur Erforschung der physiologischen Regulationsmechanismen des Haarfollikels als auch der Pathologie von Hypotrichosen besonders geeignet. Ziel dieser Arbeit ist es, eine neue Mutation als Tiermodell für die Erforschung von Erkrankungen vorzustellen, die mit Minderbehaarungen einhergehen. Mit Hilfe makroskopischer sowie licht- und elektronenmikroskopischer Befunddarstellungen werden die Defekte des betroffenen minderbehaarten Mausstammes erstmalig beschrieben.
10 10 Schrifttum 2 Schrifttum 2.1 Behaarung der Maus Embryologie der Haut und der Haarfollikel Von den drei embryonalen Keimblättern beteiligen sich das Ektoderm und das Mesoderm an der Bildung der Haut und ihrer Adnexe. Aus dem Ektoderm entwickelt sich in der frühen Ontogenese zunächst die Keimschicht in Form einer einfachen Lage kubischer Zellen. Aufgrund starker mitotischer Aktivitäten entsteht daraus eine weitere Lage flacher Zellen als Deckschicht zur äußeren Oberfläche hin. Durch das Auftreten einer weiteren Zell-Lage zwischen ihnen (Intermediärschicht) entsteht ein zunächst dreischichtiges Epithel, aus dem durch fortgesetzte Differenzierungvorgänge das spätere mehrschichtige Plattenepithel mit dem zugrundeliegenden Stratum basale und dem oberflächlichen Stratum corneum hervorgeht. Diese Vorgänge sind bei der Maus bis zum 16. Ontogenesetag abgeschlossen (MICHEL 1972). Die ersten Haaranlagen zeigen sich im Stadium der Dreischichtigkeit der Epidermis als herdförmige Verdichtungen der Basalzellen. Die Epidermis erhält dabei zunächst von der Dermis das Signal, ein Hautanhangsorgan zu bilden. Die Signalübertragung erfolgt durch einen Austausch von Botenstoffen, die auch als Morphogene bezeichnet werden (HOLBROOK et al. 1993). Dieses erste Signal ist unspezifisch und somit bei allen Vertebraten gleich. Der Epidermiszapfen, der sich bei der Maus zwischen dem 14. und 17. Ontogenesetag zur Dermis hin vorwölbt, induziert durch ein klassenspezifisches Signal die Bildung der dermalen Papille und des bei der Maus nur schwach ausgebildeten bindegewebigen Haarbalgs, der den späteren Haarfollikel umgibt (REYNOLDS u. JAHODA 1996). Die so entstandene funktionelle Einheit wird als Haarkeim bezeichnet. Dieser formt sich aufgrund eines zweiten, von der Dermis ausgehenden Signals in ein spezielles, arttypisches Anhangsgebilde um. Der Haarkeim dringt dabei von der Epidermis ausgehend schräg in die Tiefe der Dermis vor. Das freie Ende verdickt sich und wird von der dermalen Papille handschuhfingerartig nach innen eingestülpt. Es entsteht der Bulbus mit der Haarzwiebel (HARDY 1992).
11 Schrifttum 11 Auf der dermalen Papille bilden sich aus den epithelialen Keimzellen im Inneren des Bulbus die Matrixkeratinozyten, welche zunächst der Papille als Kappe aufliegen. Die Haarmatrixzellen beginnen jetzt mit einer starken mitotischen Aktivität, differenzieren sich aus und schieben sich als sogenannter Haarkegel in Richtung Hautoberfläche vor (MILITZER 1982). Auf ihrem Weg an die epidermale Oberfläche verhornen diese Zellen und bilden so das Haar Anordnung der Haarfollikel in der Haut In der Ontogenese ist die Entwicklung der Sinushaarfollikel, die die Schnurrhaare im Bereich des Kopfes bilden, zuerst abgeschlossen. Sinushaare sind bei Mäusen schon am 12. Ontogenesetag auf der epidermalen Oberfläche des Oberkiefers vorhanden. Erste Follikel der Rumpfbehaarung sind frühestens ab dem 14. Ontogenesetag histologisch erkennbar (MICHEL 1972) und werden Primärfollikel genannt. Bis zum 18. Tag der Ontogenese werden dann die Sekundärfollikel in der direkten Nachbarschaft der Primärfollikel angelegt. Diese produzieren wesentlich kleinere und feinere Haare. Durch die Entwicklung von zwei Sekundärfollikeln lateral des Primärfollikels kommt es zur Bildung von charakteristischen Dreiergruppen. Diese liegen dann in parallelen Reihen quer zur Längsachse des Tieres. Da jedoch in der Entwicklung des Tieres noch weitere Sekundärfollikel wiederum neben den schon bestehenden Sekundärfollikeln angelegt werden, verliert sich diese strenge Gruppierung mit zunehmendem Alter der Maus (CLAXTON 1966). Lediglich in der Schwanzhaut ist zeitlebens eine deutliche Anordnung in Dreiergruppen zu beobachten. Auch diese Gruppen bestehen aus zwei Sekundärfollikeln und einem in der Mitte liegenden Primärfollikel. Sie sind ebenfalls quer zur Haarrichtung bzw. der Längsachse des Tieres ausgerichtet. Im Gegensatz zu den meisten anderen Säugetieren kommen bei der Maus und beim Menschen nach DUNSTAN und LINDER (1995) nur Einzelfollikel vor. Bei diesen dringt nur ein einzelnes Haar aus einem Follikel an die epidermale Oberfläche. Verbundfollikel, bei denen mehrere Haare von verschiedenen Haarfollikeln durch eine gemeinsame Öffnung nach außen wachsen, sind vor allem für Karnivoren typisch.
12 12 Schrifttum Die Haarfollikel der Maus liegen immer streng parallel ausgerichtet in der Haut. Durch einen stumpfen Winkel zwischen den Haarfollikeln und der epidermalen Oberfläche kommt es bei der Maus, wie bei fast allen Säugern, zur Ausbildung des sogenannten Haarstrichs. Die Haare des Rumpfes zeigen dabei nach kaudal Embryologie und Morphologie der Adnexe Im Rahmen der weiteren Entwicklung der Haaranlagen entstehen bei den Säugetieren an der Hinterseite des Epithelzapfens zwei epitheliale Knospen, von denen die obere zur apokrinen Schlauchdrüse wird. Mäuse besitzen diese Drüsen in der Haut des Rumpfes nicht. Lediglich in der unbehaarten Sohlenhaut der Extremitätenenden sind derartige Schlauchdrüsen ausgebildet. Die proximal folgende Auftreibung bildet die Talgdrüsenanlage (Glandula sebacea) (SUNDBERG 1994; ACKERMANN 1997). Ihr Ausführungsgang mündet am Beginn des Infundibulums, im distalen Drittel des Haarfollikels. Sie wird durch eine Basalmembran und dermales Bindegewebe begrenzt und zeigt eine holokrine Sekretion. Ihre maximale Größe erreicht sie erst postnatal, frühestens nach Entwicklung der zweiten Haargeneration (CHASE 1954). Der Haarbalgmuskel (Musculus arrector pili) entwickelt sich dagegen auf der Hinterseite des Haarfollikels. Er erstreckt sich etwa von der Mitte des Haarfollikels bis zur interfollikulären Epidermis und besteht aus einem schmalen Band glatter Muskelzellen, die das Haar aufrichten können. Haarbalgmuskeln fehlen an manchen spezialisierten, besonders kräftig ausgebildeten Haaren, wie den Wimpern oder den Sinushaaren (PINKUS 1958) Morphologie des Haarfollikels Ein aktiver haarbildender Follikel ist ein kompliziert aufgebautes, aus mehreren konzentrischen Schichten bestehendes Hautanhangsgebilde. Er wird von dem ihn mitversorgenden bindegewebigen Haarbalg und der darüberliegenden hyalinen Membran (auch Basalmembran oder Glashaut ) umgeben. Während der aktiven Wachstumsphase des Haarfollikels erstrecken sich die Haarfollikel der Maus durch die gesamte Subkutis bis direkt an die Grenze zu dem darunterliegenden Hautmuskel.
13 Schrifttum 13 Bulbus und Papille können dabei gegenüber dem distalen Teil des Haarfollikels deutlich abgewinkelt sein (ZAUN 1968; MILITZER 1982). Im Längsschnitt kann man dabei einen oberen und einen unteren Abschnitt unterscheiden. Der obere Abschnitt besteht aus dem Infundibulum, der die trichterförmige Öffnung des Haarfollikels an die epidermale Oberfläche darstellt und dem darunterliegenden Isthmus. Der Isthmus schließt sich als kurzer, verengter Abschnitt des Follikelkanals proximal an das Infundibulum und den daruntergelegenen Talgdrüsenausführungsgang an. Der untere Abschnitt des Haarfollikels besteht aus dem langen und geraden Follikelschaft und seiner proximalen Auftreibung, dem Bulbus (ACKERMANN 1997) (siehe Abbildung 2.1). Die äußerste Schicht des eigentlichen Haarfollikels ist die äußere Wurzelscheide (ORS, engl.: Outer Root Sheath ). Sie geht kontinuierlich in das Stratum basale der Epidermis über und stellt die Fortsetzung dieser Schicht in den zylindrischen Haarfollikel dar. Sie ist bei der Maus lediglich eine bis maximal zwei Zell-Lagen dick. Aufgrund von häufigen Überlappungen der Zellen erscheint die äußere Wurzelscheide gelegentlich mehrschichtig. Auch die unter der äußeren Wurzelscheide liegende hyaline Membran geht kontinuierlich in die Basalmembran der Epidermis über (ROTH u. HELWIG 1964a; ITO 1988). Darauf folgt nach innen die röhrenförmige innere Wurzelscheide (IRS, engl.: Inner Root Sheath ). Sie ist im Gegensatz zu der äußeren Wurzelscheide ebenso wie das eigentliche Haar eine Bildung der Matrixzellen des Bulbus. Sie besteht ihrerseits aus drei morphologisch verschiedenen, konzentrischen Schichten (SUNDBERG 1994; ROOK u. DAWBER 1995). Von außen nach innen sind dies die Henlesche Schicht, die Huxleysche Schicht und die Wurzelscheidenkutikula. Die Wurzelscheidenkutikula stellt die Grenze der inneren Wurzelscheide zum eigentlichen Haar dar. Die Zellen der einzelnen Schichten überlappen dabei einander nicht (SERRI u. CERIMELE 1991).

14 14 Schrifttum Abbildung 2.1: Schematischer Längsschnitt durch einen aktiven murinen Haarfollikel * * modifiziert nach DAWBER 1995 Abbildung 2.2: Schematischer Querschnitt durch die suprabulbäre Region eines aktiven Haarfollikels
15 Schrifttum 15 Das Haar selbst liegt innerhalb der inneren Wurzelscheide und weist mit Kutikula, Rinde (Kortex) und Mark (Medulla) - von außen nach innen - ebenfalls drei verschiedene Schichten auf, wie in Abbildung 2.2 dargestellt. Außer durch die im Bulbus liegende mesenchymale dermale Papille, wird die innere Wurzelscheide eines aktiven haarbildenden Follikels durch die sie umgebende äußere Wurzelscheide ernährt und vermutlich auch reguliert (PAUS et al. 1994). Im Gegensatz zu anderen Säugetieren finden sich in der murinen dermalen Papille keine Blutgefäße. Aufgrund der geringen Größe kann die Versorgung der epithelialen Zellen alleine durch Diffusionsvorgänge gesichert werden. Die Größe der dermalen Papille bestimmt dabei die Größe des von ihr versorgten Follikels (VAN SCOTT u. EKEL 1958). Bei pigmentierten Tieren finden sich an der Spitze der dermalen Papille zwischen den germinativen Zellen der Haarmatrix interdigitierende Melanozyten. Diese haben ihren Ursprung in der Neuralleiste und lagern ihre Pigmentgranula in der Haarrinde und in der äußeren Wurzelscheide ab. Ihre follikuläre Melanogenese ist streng an die aktive Wachstumsphase des Follikels gekoppelt (PAUS et al ). Im Bulbus liegen die stark mitotisch aktiven Matrixkeratinozyten der dermalen Papille kappenartig auf. Durch fortlaufende Teilungsvorgänge bilden sie den sich nach distal, zur Hautoberfläche schiebenden Haarkegel. Aus den morphologisch identischen Matrixzellen entstehen dabei sowohl die drei verschiedenen Schichten des Haares als auch die drei Schichten der inneren Wurzelscheide. Man nimmt heute an, daß die Differenzierung der einheitlichen Matrixzellen in die jeweiligen terminalen Zellen in den verschiedenen Schichten der inneren Wurzelscheide und des Haares wiederum durch die dermale Papille gesteuert wird (JAHODA u. REYNOLDS 1993). Der genaue Mechanismus ist dabei jedoch noch ungeklärt (HARDY 1992).
16 16 Schrifttum Abbildung 2.3: Matrixzellpopulation im Bulbus eines aktiven Haarfollikels (A) und deren Wachstumsrichtung (B) * * modifiziert nach HARDY 1992 Das Einsetzen der Verhornung erfolgt in den unterschiedlichen Schichten des Follikels zu verschiedenen Zeiten. Erste verhornte Zellen sind in der Henleschen Schicht der inneren Wurzelscheide schon etwa in der mittleren Höhe des Bulbus zu beobachten. Die Kutikula der inneren Wurzelscheide ist die zweite Schicht des Haarfollikels mit erkennbarer Keratinisierung. Erst danach verhornen die Rinde und dann das Mark des Haares selbst. Zuletzt verhornen die Huxleysche Schicht der inneren Wurzelscheide und schließlich auch die Kutikula des Haares auf ihrem Weg zur epidermalen Oberfläche (FORSLIND 1990). Durch eine dem Haar vorangehende Verhornung dient also die innere Wurzelscheide dem sich entwickelnden Haar als stabiler, formgebender Hohlzylinder. Auf der Höhe des Ausführungsganges der Talgdrüsen ist die innere Wurzelscheide bereits vollständig verhornt. Sie zerfällt bei ihrer Wanderung zur epidermalen Oberfläche und ihre Reste werden mit dem wachsenden Haar über die Öffnung des Haarfollikels (Infundibulum) nach außen befördert. Die innere Wurzelscheide ist zudem nur in der aktiven Wachstumsphase des Haarfollikels ausgebildet. Die äußere Wurzelscheide
17 Schrifttum 17 ist dagegen während aller Aktivitätsstadien des Follikels relativ unverändert vorhanden (ITO 1988). Auf der Höhe der Ansatzstelle des Musculus arrector pili liegt in der äußeren Wurzelscheide der sogenannte Wulst. Bei Primaten und Mäusen wird in ihm der Sitz der langsam proliferierenden Stammzellen vermutet aus denen bei späteren Haarzyklen die Zellen der inneren Wurzelscheide und des Haares hervorgehen (LAVKER et al. 1993; WILSON et al. 1994; REYNOLDS u. JAHODA 1994; AKIYAMA et al. 1995; LAVKER u. SUN 1995). Bei den anderen Säugern ist die Lokalisation dieser Stammzellen noch umstritten (DUNSTAN u. LINDER 1995). Neben den normalen Haarfollikeln der Rumpfbehaarung existieren noch spezielle Haarfollikel, die der taktilen Reizwahrnehmung dienen. Diese weisen zusätzlich folgende anatomische Besonderheiten auf: Die Follikel der Sinushaare (auch Vibrissen oder Schnurrhaare ) unterscheiden sich von dem normalen Deckhaarfollikel durch ihre Einbettung in einen zweigeteilten, blutgefüllten Hohlraum, dem Ringsinus. Dieser ist von einer großen Menge sensibler Rezeptoren durchzogen, die über den Nervus trigeminus mit definierten Hirnrindenfeldern in Verbindung stehen (PARAKKAL 1969; YAMAKADO u. YOHRO 1979). Desweiteren weisen die besonderen taktilen Haare des Rumpfes (Leithaare, siehe unten) als spezielle Einrichtungen Haarscheiben und Annularringe auf. Haarscheiben sind Mechanorezeptoren am Infundibulum, Annularringe stellen Blutsinus dar, die oberhalb des Bulbus liegen (STRAILE u. MANN 1973).
18 18 Schrifttum Ultrastruktur der Haarfollikelzellen Matrixzellen des Bulbus Die germinativen Zellen des Bulbus besitzen einen großen Zellkern, der den größten Teil ihres Zellvolumens einnimmt. In ihrem spärlichen Zytoplasma finden sich viele Ribosomen und einige Mitochondrien. Der Rest des Zytoplasmas setzt sich aus rauhem endoplasmatischem Retikulum, einem kompakten Golgi-Apparat in der Nähe des Nukleus und einer großen Anzahl von Zisternen und Vesikeln zusammen. Die Matrixzellen sind stark mitotisch aktiv, weshalb in vielen Zellen Zentrosomen ausgebildet sind. Zwischen den benachbarten Zellen finden sich desmosomale Verbindungen und Gap junctions. Im Gegensatz zu den Desmosomen der epidermalen Keratinozyten strahlen jedoch von diesen keine Filamente in das Zytoplasma aus. Es bestehen wesentlich weniger Anheftungsstellen zwischen den Matrixzellen als in der Basalschicht der Epidermis. Dies erleichtert möglicherweise die Wanderung der Zellen von der Matrix in die suprabulbären Bereiche (ROTH u. HELWIG 1964a; POWELL u. ROGERS 1997). Die Differenzierung der im Haarfollikel aufsteigenden Matrixkeratinozyten wird zuerst durch deren unterschiedliche Zellformen sichtbar. Die Zellen des Haarmarkes bleiben relativ groß und sphärisch, während die Zellen der späteren Rindenschicht eine spindelige Form annehmen. Die Längsachse dieser spindeligen Haarrindenzellen ist dabei parallel zu der Wachstumsrichtung des Haares ausgerichtet Haarbildende Zellen Die Matrixzellen, aus denen das Mark des Haares entsteht, liegen zentral über der dermalen Papille. Sie sind die ersten Zellen, die mikroskopische Veränderungen erkennen lassen. Ausgehend vom Golgi-Apparat entwickeln sich unregelmäßig große, runde, elektronendichte Granula. Diese entsprechen den Keratohyalingranula der Epidermis und werden Trichohyalingranula genannt. Sie sind nicht durch eine Membran begrenzt und konfluieren mit zunehmender Differenzierung der Zelle. Die Granula nehmen dadurch an Größe zu; ihr Durchmesser beträgt dann maximal ein bis zwei Mikrometer (ROTH und HELWIG 1964b). Mit fortschreitender Differenzierung treten insbesondere in der Nähe des Nukleus reichlich
19 Schrifttum 19 Glykogengranula auf, während andere zytoplasmatische Organellen zerfallen. Die mit Christae versehenen Mitochondrien schwellen an und vakuolisieren. Das Trichohyalin fusioniert schließlich zu einer soliden Masse und die Zellen dehydrieren und kollabieren. Bis auf Reste der Zellkerne sind keine weiteren Strukturen von Zellorganellen mehr zu finden. Während des gesamten Ausreifungprozesses der Matrixkeratinozyten, die das Haarmark bilden, werden keine Filamente in ihrem Zytoplasma ausgebildet. Die Zellen des Haarmarkes werden nach Beendigung ihrer Differenzierung durch luftgefüllte Hohlräume voneinander getrennt. Dadurch wird die Isolationswirkung des Haarkleides wesentlich verbessert (ROOK u. DAWBER 1995) (siehe Abbildung 2.6). In der Zone ihrer Keratinisierung distal des Bulbus zeigen die Zellen, die die Rindenschicht der Haare bilden, eine intensive Proteinsynthese. Sie besitzen deshalb eine große Anzahl von Polysomen und eine starke RNS- Anfärbbarkeit von Nukleus und Zytoplasma. Im Zytoplasma finden sich große Mengen von Keratinen und assoziierten Proteinen. Diese aggregieren zu Mikrofibrillen, die sich wiederum zu Makrofibrillen zusammenlagern (siehe Kapitel 2.1.6). Sie zeigen mit zunehmender Differenzierung der Rindenzellen (und damit Entfernung vom Bulbus) eine deutliche Ausrichtung parallel zur Wachstumsrichtung des Haares. Diejenigen Zellen der Haarrinde, die eine Anordnung der Keratine zu parallelen Makro- und Mikrofibrillen erkennen lassen, werden dem sogenannten Orthokortex des Haares zugeordnet. Liegen die Keratine eher in einer sogenannten quasikristallinen, hexagonalen Form angeordnet, zählt man diese Zellen zum Parakortex der Haarrinde (POWELL u. ROGERS 1997). Es ist nur selten möglich, beide Arten der Keratinanordnungen innerhalb einer einzigen Zelle vorzufinden. Innerhalb eines Haares bilden Zellen mit der gleichen Keratinanordnung verschiedene Bereiche der Rinde bilateral-symmetrisch aus. So findet man beim gewellten Haar der Schafe in dem konkaven Bereich des Haares die parakortikalen Zellen, auf der gegenüberliegenden Seite dagegen die orthokortikalen Zellen. Die Regulationsmechanismen für die Bildung dieser unterschiedlichen Muster der Keratinanordnung in der Haarrinde sind jedoch bis heute nicht vollständig aufgeklärt (GILLESPIE 1991; SWIFT 1997).
20 20 Schrifttum Im Gegensatz zu den Zellen des Haarmarkes sind die Zellen der Haarrinde frei von Trichohyalingranula, wie schon ROTH und HELWIG (1964b) festgestellt haben. In den weiter distal gelegenen Bereichen finden sich vermehrte Anzeichen der Zytolyse. Lysosomen degenerieren, die Zellkerne werden pyknotisch und schließlich degradieren die Ribosomen (PARAKKAL 1969; ROGERS u. POWELL 1994). Die Zellen, aus denen die Haarkutikula hervorgeht, flachen mit zunehmender Differenzierung stark ab. Ihre distalen Enden überlappen dabei einander. Es finden sich reichlich Desmosomen und Tonofilamente an den Kontaktflächen zu den benachbarten Kutikulazellen. Im Gegensatz zu den Zellen von Haarrinde oder Haarmark werden keine Filamente und keine Trichohyalingranula in den Zellen der Haarkutikula gebildet (ROTH u. HELWIG 1964b). Man unterscheidet bei den ausdifferenzierten Kutikulazellen drei verschiedene Zonen. Die innerste Lage ist dabei die sogenannte Endokutikula. Darauf folgt nach außen die Exokutikula, welche an ihrer Oberfläche von der sogenannten A-Schicht bedeckt wird. Die Schicht der Endokutikula entsteht aus den Resten des Zytoplasmas und der darin enthaltenen Organellen der Kutikulazelle. Distal des Bulbus sind in den Zellen der Haarkutikula zahlreiche Proteingranula zu sehen. Diese aggregieren im Verlauf der Differenzierung der Zellen und bilden die Exokutikula sowie die stark elektronendichte A- Schicht. Die Exokutikula nimmt etwa 50% der Fläche einer quergeschnittenen Kutikulazelle ein (POWELL u. ROGERS 1997). Die Melanozyten des Bulbus lagern bei pigmentierten Tieren ihre Pigmentgranula nur in Mark und Rinde ab, nicht jedoch in der Kutikula oder der inneren Wurzelscheide (PAUS et al. 1994).
21 Schrifttum Zellen der Wurzelscheiden Die Zellen aller drei Schichten der inneren Wurzelscheide (Kutikula, Huxleysche und Henlesche Schicht) zeigen eine gleichartige Differenzierung, die allerdings mit unterschiedlicher Geschwindigkeit abläuft. Zuerst differenzieren sich die Zellen der Henleschen Schicht, dann die der Kutikula und schließlich die Zellen der Huxleyschen Schicht (ROTH u. HELWIG 1964a). Die Keratinisierung der Zellen geht mit einer Abflachung und der Bildung zunehmender Mengen elektronendichter Granula einher. Die Granula sind nicht von einer Membran umgeben und enthalten wie die Zellen des Haarkmarkes Trichohyalin. Im Gegensatz zu den Markzellen sind die Trichohyalingranula dabei meist Filamenten angelagert. Warum die Trichohyalingranula nur in der inneren Wurzelscheide filamentassoziiert vorliegen ist nicht bekannt (ROGERS u. POWELL 1994). In den letzten Stadien der Differenzierung lösen sich der Kern und alle noch vorhandenen Zellorganellen auf. Die Trichohyalingranula dissoziieren und das Trichohyalin verteilt sich diffus zwischen den Filamenten. Die Zellmembranen verdicken sich, die Zellen dehydrieren und schrumpfen mit zunehmender Keratinisierung. In den unteren Anteilen des Follikels wird der Kontakt zwischen den einzelnen Schichten der inneren Wurzelscheide und zur äußeren Wurzelscheide durch Desmosomen und Gap junctions aufrecht erhalten. In Höhe des Infundibulums lösen sich diese Verbindungen und die Zellen werden einzeln oder in Gruppen in das Lumen des Follikels abgestoßen (ROTH u. HELWIG 1964a). Die Zellen der äußeren Wurzelscheide entsprechen in ihrem ultrastrukturellen Aufbau den epidermalen Keratinozyten (BLUMENBERG u. TOMIC-CANIC 1997). Zwischen der epidermalen Schicht des Haarfollikels und der Epidermis der Haut bestehen nur quantitative Unterschiede: Die Ribosomen der Zellen der äußeren Wurzelscheide sind zu Polysomen aggregiert und liegen frei im zytoplasmatischen Raum. Das endoplasmatische Retikulum ist nur schwach entwickelt. Auch die Golgi- Vesikel haben nur eine geringe Größe. Die Keratohyalingranula der Zellen der äußere Wurzelscheide sind ebenfalls klein.
22 22 Schrifttum Es überwiegen membrangebundene Granula im Zytoplasma der Zellen der äußeren Wurzelscheide (DAWBER 1995). Die wenigen, in den Zellen der äußeren Wurzelscheide gebildeten Filamente, liegen in relativ ungeordneten Bündeln diffus verteilt im Zytoplasma. Zu den benachbarten Zellen der äußere Wurzelscheide sind zahlreiche Desmosomen zu finden. Die der Basalmembran anliegende Seite der Zellen läßt eine große Zahl von Hemidesmosomen erkennen (ROTH u. HELWIG 1964a). Die äußere Wurzelscheide wird als verhältnismäßig statische Region angesehen. Ihre Zellen bewegen sich kaum mit den Zellen der inneren Wurzelscheide zusammen nach distal. Eine Keratinisierung der Zellen der äußeren Wurzelscheide findet lediglich im Bereich des Infundibulums statt. Sie verläuft in zentripetaler Richtung und wird als trichilemmale Keratinisierung bezeichnet (ITO 1988) Keratine des Haarfollikels und des Haares Keratin ist ebenso wie in der Hornschicht der Epidermis und in der Nagelplatte auch im Haar das wichtigste Strukturprotein. Es kann als Filament- oder als Matrixtyp im Haar und im Haarfollikel vorkommen. Keratine sind hochdifferenzierte, widerstandsfähige und stark unlösliche Proteine in den Epithelien der Vertebraten. Die meisten Bestandteile der Keratinfilamente kommen bereits frühzeitig in den Keratinozyten vor, aus denen später die Hornzellen in der Epidermis, dem Haar und dem Nagel hervorgehen (LANE et al. 1985; BLUMENBERG u. TOMIC-CANIC 1997). Im Haarfollikel ist die Differenzierung der Keratinozyten ein komplexer Vorgang, da er mehrere unterschiedliche, konzentrische Ringe von Keratinozyten mit unterschiedlichen Besonderheiten der Verhornung einschließt. Die Hauptzone der Verhornung (die sogenannte Keratinisierungszone) liegt etwa in der Mitte des Haarfollikels und kann besonders gut an den Zellen beobachtet werden, die später die Rinde des Haares bilden.
23 Schrifttum 23 Bei der Aushärtung und Verhornung der sich differenzierenden und im Follikel aufwärts wandernden Matrixkeratinozyten werden sogenannte Makrofibrillen gebildet. Diese sind in Längsrichtung des Haares angeordnet und in eine intermakrofibrilläre Matrix aus verschiedenen Proteinen eingebettet (siehe Abbildung 2.6). Die Makrofibrillen sind aus Mikrofibrillen aufgebaut, die wiederum aus Keratin-Intermediärfilamenten (IF) bestehen. Diese sind ebenfalls in eine intermikrofibrilläre Matrix eingebettet. Die Keratin- Intermediärfilamente besitzen eine stabförmige, α-helikale Struktur mit einem Durchmesser von acht bis zehn Nanometern. Sie sind ebenfalls in Längsrichtung des Haares angeordnet (ROGERS u. POWELL 1994). Die für die Intermediärfilamente momentan gebräuchliche Nomenklatur nach POWELL und ROGERS (1997) - modifiziert nach MOLL et al. (1993) - benennt 18 saure Typ-I Keratine von epidermalen Intermediärfilamenten als K 1.1 bis K 1.5 und K 1.9 bis K Von ihnen werden 18 basische Typ-II Keratine K 2.1 bis K 2.17 (K 2.6 a und b) unterschieden. Oftmals werden die Keratine paarweise gebildet. Ein Paar besteht dann aus einem sauren Keratin und seinem basischen Partner. Der Verhornungsprozeß der Haarfollikel-Keratinozyten, die auch als Trichozyten bezeichnet werden, entspricht dem der epidermalen Keratinozyten. Analog zu den Beobachtungen an epidermalen Keratinozyten werden bei den Trichozyten im Verlauf der Verhornung unterschiedliche Keratin-Intermediärfilamente gefunden. In den basalen Lagen der Epidermis können beispielsweise die Keratine K 2.5 und K 1.14 nachgewiesen werden. In sich differenzierenden epidermalen Keratinozyten findet sich dagegen das Keratinpaar K 2.1 und K Diese Keratine sind ebenfalls in den sich differenzierenden Zellen der inneren Wurzelscheide und der Haarkutikula vorhanden (ROGERS u. POWELL 1994). Unterschiede in der Expression der Keratin-Intermediärfilamente können aber nicht nur im zeitlichen Verlauf der Differenzierung der Keratinozyten und Trichozyten beobachtet werden. Auch innerhalb der verschiedenen Lokalisationen des Haarfollikels bzw. des Haares, werden unterschiedliche Keratin-Intermediärfilamente nachgewiesen. Die unterschiedlichen Zelltypen der Wurzelscheiden bilden verschiedene charakteristische Keratine aus.
24 24 Schrifttum So wurden in der äußeren Wurzelscheide die für epidermale Basalzellen typischen weichen, sogenannten Soft-Keratins nachgewiesen. Es handelt sich dabei um die Keratine K 2.5, K 2.6, K 1.14 und K Das Keratinpaar K 2.1 und K 1.10, welches für die innere Wurzelscheide kennzeichnend ist, kann dagegen in den Zellen der äußeren Wurzelscheide nicht nachgewiesen werden. Die Ausbildung von K 2.1 und K 1.10 bleibt ausschließlich auf die inneren Wurzelscheiden beschränkt (GOLDSMITH u. LOWELL 1991; LIMAT et al. 1991). Neuere Studien konnten zeigen, daß die Zellen der äußeren Wurzelscheide die einzigen Zellen der Haut sind, in denen physiologischerweise K 1.17 während aller HaarzyIusphasen hindurch gebildet wird (PANTELEYEV et al. 1997). Aus der Rinde des Haares sind vor allem die sauren Keratin-Intermediärfilamente K 1.1 bis K 1.5 und die basischen Keratine K 2.9 bis K 2.12 und K 2.16 isoliert worden (LANE et al. 1985; POWELL u. ROGERS 1997). Die Keratin-Intermediärfilamente des Haares besitzen an ihren Enddomänen charakteristische Aminosäuren. Im Gegensatz zu den Intermediärfilamenten der Epidermis sind ihre Enden nicht mit der Aminosäure Glycin versehen. Die Keratin-Intermediärfilamente des Haares besitzen ausschließlich schwefelreiche Cystin-Enden. Dies ermöglicht, daß im Verlauf des Keratinisierungsprozesses durch oxidative Vorgänge stabile Disulfidbrückenbindungen entstehen. Vor allem die Rindenschicht und die Kutikula des Haares enthalten große Mengen von Cystinsulfhydrilgruppen, die in den keratinisierten Trichozyten zu beinahe 100 Prozent in Cystindisulfidbrückenbindungen übergehen. Durch diese Quervernetzung der Intermediärfilamente erhält der Haarschaft seine große Festigkeit und Stabilität. In den Wurzelscheiden, besonders in der Henle-Schicht, finden sich dagegen nur geringe Mengen dieser Brückenbindungen (SWIFT 1997).
25 Schrifttum 25 Abbildung 2.4: Aus den Haarfollikelstammzellen hervorgehende Zelltypen und ihre charakteristischen Proteine Haarfollikelstammzellen ORS-Zellen IRS-Zellen Haarzellen Markzellen Henle Huxley Kutikula Kutikula Rinde Epidermale IF Trichohyalin KAP 5 IF Trichohyalin IF IF KAP 1-4 KAP 6-8 Abkürzungen: IF: Intermediärfilamente; IRS: innere Wurzelscheide; KAP: Keratin-assoziierte Proteine; ORS: äußere Wurzelscheide Die Keratin-Intermediärfilamente werden innerhalb des Trichozyten der Haarinde durch eine spezielle Gruppe von Proteinen voneinander getrennt. Es handelt sich dabei um die sogenannten Keratin-assoziierten Proteine (KAP). Diese bilden die intermakro- oder intermikrofibrilläre Matrix zwischen den Fibrillen und variieren speziesabhängig in ihrer relativen Verteilung. Aufgrund ihres unterschiedlichen Gehaltes verschiedener, schwefelhaltiger Aminosäuren können 11 Gruppen von KAPs mit einer jeweils variablen Anzahl von Mitgliedern differenziert werden (POWELL u. ROGERS 1997). Im Laufe des Keratinisierungsprozesses werden die verschiedenen Keratin-assoziierten Proteine zu verschiedenen Zeitpunkten gebildet. Es werden immer zuerst die Keratin-Intermediärfilamente produziert, worauf dann erst die Bildung der KAPs einsetzt.
26 26 Schrifttum Wie in Abbildung 2.5 dargestellt, können zunächst die KAPs der Gruppe 6 bis 8 und dann die KAPs der Gruppe 1 bis 4 in den Trichozyten der Haarinde im unteren Haarfollikel durch Analyse der KAP-Transkripte nachgewiesen werden. Das KAP 5 erscheint mit dem KAP 10 als letztes in der Kutikula des sich entwickelnden Haares (MOLL et al. 1993; ROGERS u. POWELL 1994). Bei der Maus werden zudem die Gene für die Bildung der KAPs 9 und 11 in allen Zellen der Haarrinde im proximalen Haarschaft exprimiert (POWELL u. ROGERS 1997).

27 Schrifttum 27 Abbildung 2.5: Die Expression der Keratin-Intermediärfilament- und KAP-Gene während der Differenzierung der Trichozyten im Haarfollikel * Abkürzungen: IRS: innere Wurzelscheide; KAP: Keratin-assoziiertes-Protein; Keratin-IF: Keratin-Intermediärfilament; ORS: äußere Wurzelscheide * modifiziert nach ROGERS u. POWELL 1997 Erläuterung: Die Exprimierung der Gene für die verschiedenen KAPs findet zu unterschiedlichen Zeitpunkten während der Differenzierung der Trichozyten des Haarfollikels statt. Die KAPs werden dabei immer zeitlich nach den Keratin-Intermediärfilamenten gebildet. Die gemusterten Flächen innerhalb des Haarfollikels sollen die zeitlich verschiedene Exprimierung der KAPs verdeutlichen. Innerhalb der verschiedenen Zellschichten des Haarfollikels werden unterschiedliche KAPs bilateral-symmetrisch produziert.
28 28 Schrifttum Ein weiteres wichtiges Strukturprotein des Haarfollikels ist das Trichohyalin. Wie bereits beschrieben, findet man Trichohyalin vor allem in der inneren Wurzelscheide und im Haarmark in Form großer Granula. Diese Trichohyalingranula werden nicht von einer Membran begrenzt und entsprechen den in den Keratinozyten der Haut vorzufindenden Keratohyalingranula. Innerhalb der Trichozyten der inneren Wurzelscheide liegen die Trichohyalingranula intermediärfilament-assoziiert vor. Man nahm früher an, daß sich die Intermediärfilamente aus den Granula entwickeln würden, was jedoch inzwischen widerlegt wurde (PARAKKAL 1969; POWELL u. ROGERS 1997). Das Trichohyalin verteilt sich mit zunehmender Differenzierung der Zellen zwischen den Intermediärfilamenten und vernetzt diese zusätzlich. Durch eine biochemische Reaktion (Desaminierung von Arginintrichohyalin und Bildung von Citrullin durch Peptidylarginin-deiminase, PAD) kommt es zu einer Verhärtung dieser Schichten des Haarfollikels (ROGERS et al. 1997). Dadurch kann eine Einsparung großer Mengen schwefelreicher Aminosäuren erreicht werden. Möglicherweise ist dies der physiologische Sinn dieses Vorganges, denn diese Verbindungen können hierdurch ausschließlich für die Keratinisierung von Kutikula und Rinde des Haares verwendet werden (DAWBER 1995; ROGERS u. POWELL 1994). In der Umgebung des Trichohyalins der Markzellen sind im Gegensatz zu den Zellen der inneren Wurzelscheide keine Keratin-Intermediärfilamente vorzufinden. Die Trichohyalingranula des Haarmarkes fusionieren während der Ausreifung der Zellen zu einer soliden Masse, ohne daß eine Vernetzung zwischen Filamenten, wie in der inneren Wurzelscheide, stattfindet. Die ersten Kennzeichen der beginnenden Aushärtung der Trichozyten ist die Bildung von mrna für Trichohyalin schon in Höhe des Bulbus. Erst danach ist es möglich, mrna für die Bildung von Keratin-Intermediärfilamenten im Rahmen der Entwicklung des Haarkortex nachzuweisen. Zuletzt werden mrna-sequenzen für die KAPs gebildet.

29 Schrifttum Morphologie des Haares Schon 1926 beschreib DRY als Erster ausführlich und umfassend die Morphologie der Haare der Maus. An jedem Haar lassen sich in Längsrichtung drei Abschnitte unterscheiden. Von distal nach proximal sind dies die dünn auslaufende Haarspitze, die mittlere, dickere Zone der Granne und der sich wieder verjüngende Haarschaft. Im Querschnitt unterscheidet man - von außen nach innen - die Kutikula, die Rinde (Kortex) und das Haarmark (Medulla), sofern es ausgebildet ist. Abbildung 2.6: Schematische Darstellung der bedeutenden zellulären Strukturen eines Haares * * modifiziert nach ROGERS u. POWELL 1997
30 30 Schrifttum Die Kutikula des Haares besteht aus einer dünnen, einschichtigen Zell-Lage. Die einzelnen Zellen der Kutikula überlappen einander, wobei die freien Ränder nach distal zeigen. Durch die entgegengesetzte Schichtung der Zellen der Wurzelscheidenkutikula ergibt dies eine Verankerung des Haares im Follikel. Aufgrund ihrer fischschuppenartigen Überlappung werden die Zellen der Kutikula auch Kutikulaschuppen genannt. Das Aussehen der Kutikulaschuppen hängt in erster Linie vom Durchmesser des Haares ab. Wenn sie den ganzen Umfang des Haares umkleiden werden sie kranzförmig genannt. Dachziegelartige Schuppen bedecken dagegen nur einen Teil des Haarumfangs (FORSLIND 1990). Die Kutikulaschuppen zeigen im Verlauf des Haares, bedingt durch den zu- und wieder abnehmenden Durchmesser, ein wechselndes Muster. Die Rinde stellt eine Röhre aus langgestreckten und verhornten Zellen dar. Diese Zellen sind vollständig von regelmäßig angeordneten Keratinmassen ausgefüllt. Eine Gruppe sogenannter Mikrofibrillen aggregiert dabei zu den bereits beschriebenen Makrofibrillen. Die Fibrillen werden durch die interfibrillären Matrixproteine (KAPs) voneinander getrennt. Beide Arten der Fibrillen sind in Längsrichtung des Haares ausgerichtet, wie in Abbildung 2.6 dargestellt. Meist wird die Haarrinde durch das Mark ausgefüllt, welches im Laufe der Verhornung Luft aufnimmt. Durch nach innen gerichtete Projektionen der Rindenzellen und deren Vereinigung erscheint das Mark im Längsschnitt leitersprossenartig unterbrochen. Diese Fortsätze werden auch als Trabekel bezeichtet. Fünf Arten der Ausbildung des Haarmarkes werden nach SUNDBERG u. HOGAN (1994) unterschieden: es kann vollständig abwesend sein, einzelne Luftkammern können vorhanden sein (diskontinuierlich) oder wenige, vorhandene Luftkammern können regelmäßig (intermediär) oder unregelmäßig (fragmentiert) gruppiert sein. Das Haarmark kann auch vollständig durchgehend, also kontinuierlich, ausgebildet sein. Das Vorhandensein eines Haarmarkes ist in erster Linie vom jeweiligen Durchmesser des Haares abhängig. So haben feine Haare nur ein dünnes oder gar kein Mark. Auch Spitze und Schaft können marklos sein, während die Granne des selben Haares ein Mark aufweist.
31 Schrifttum 31 Dicke Haare, wie z. B. die Ahlenhaare, weisen dagegen im Bereich der Granne ein bis zu vier-reihiges Mark auf. Der Querschnitt der Haare ist zumeist nicht rund, sondern weist eine konkave Einziehung auf, eine sogenannte Rinne (SUNDBERG u. HOGAN 1994) Haararten der Maus Anhand der anatomischen Lage und der besonderen morphologischen Ausstattung lassen sich bei der Maus nach SUNDBERG und HOGAN (1994) bis zu acht verschiedene Haararten unterscheiden. Eine Differenzierung nach Haararten kann anhand der Länge und Dicke des Haares, der Gestaltung des Haarmarks, des Verhältnisses von Rinde zu Mark, der Ausbildung der Kutikulaschuppen und des Vorhandenseins von charakteristischen Abknickungen erfolgen. Neben den Haaren des Rumpfes gibt es spezielle Haare am Schwanz, an der Schnauze (Vibrissen), den Füßen, den Augen (Wimpern), den Ohren, den Zitzen und an der Perianal-/ Genitalregion. Die Vibrissen (Schnurrhaare) sind, wie bei den meisten Säugern, die größten Haare der Maus. Sie sind an der Schnauze, in der Umgebung des Auges und an den Extremitäten zu finden. Von besonderer Bedeutung für Untersuchungen des Haarkleides ist die Rumpfbehaarung mit den bei der Maus ausgebildeten vier Subtypen: Man unterscheidet zwischen den Leithaaren, den Ahlenhaare, den Knickhaaren und den Zick-Zack-Haaren. Es existierten in der Vergangenheit mehrere verschiedene Bezeichnungen für die Haartypen des Rumpfes der Maus und anderer Säugetiere. Man hat sich heute auf die folgende Nomenklatur geeinigt (SUNDBERG u. HOGAN 1994): Die Leit-, Ahlen- und Knickhaare gehören der Hauptgruppe der Deckhaare an. Zu diesen werden außerdem bei anderen Säugetieren noch die Stacheln (z. B. des Igels) gezählt. Die Zick-Zack-Haare zählt man zu der Gruppe der Flaumhaare. Diese Gruppe umfaßt außerdem die Lanugohaare des Menschen (siehe Tabelle 2.1) und die Wollhaare (z. B. des Schafes). Bei der Maus sind jedoch nur die Zick-Zack-Haare ausgebildet.
32 32 Schrifttum Tabelle 2.1: Haartypen des Rumpfes Hauptgruppen von Haaren Subtypen der Haare bei Anteil an Behaarung anderen Säugetieren der Maus % * 1. Deck- oder Fellhaare 1.1 Stacheln Leit- oder Borstenhaare 1.2. Leithaare 2% 1.3. Grannenhaare Ahlenhaare 28% Knickhaare 2. Flaum- oder Wollhaare 2.1. Lanugohaare Flaumhaare 2.2. Zick-Zack-Haare 70% 2.3. Wollhaare [* Prozentangaben bei der Maus nach SUNDBERG u. HOGAN (1994)] ad 1.2. Die Leithaare sind gerade und besonders lange Deckhaare, die vorwiegend im Rückenbereich der Tiere zu finden sind. Sie sind im Querschnitt überwiegend rund und laufen in einer besonders langen, fein ausgezogenen Spitze aus. Diese marklose Spitze nimmt mindestens ein Fünftel der Länge des gesamten Haares ein und ragt über die Spitzen der anderen Haare des Felles hinaus. Leithaare besitzen ein ein- bis zweireihiges Mark und eine im Vergleich zu den Zick-Zack-Haaren relativ dicke Rinde. Ihre Kutikularschuppen liegen glatt an, sind aber verhältnismäßig lang ausgezogen (siehe Abbildung 3.8). ad Die Hauptmasse der Deckhaare bilden die Ahlenhaare. Sie besitzen ebenfalls eine gerade Form, sind aber deutlich dicker als die Leithaare. Ihr Haarmark weist im Bereich der Granne immer mindestens zwei bis vier nebeneinanderliegende Reihen von Zellen bzw. Luftkammern auf. Dieses immer mehrreihig ausgebildete Haarmark ist das kennzeichnende Merkmal der Ahlenhaare.
33 Schrifttum 33 Die Kutikularschuppen der Ahlenhaare überlappen im Bereich der Granne sehr stark, so daß sie im Vergleich zu denen der Leithaare recht kurz erscheinen. Zudem erhalten die Ahlenhaare durch eine besonders deutliche, parallel zur Längsachse des Haares verlaufende rinnenartige Einziehung einen bohnenförmigen Querschnitt. ad Die Knickhaare sind den Ahlenhaaren morphologisch sehr ähnlich. Sie werden daher von einigen Autoren nicht von den Ahlenhaaren unterschieden und mit diesen gemeinsam als Grannenhaare bezeichnet. Meist enthalten die Knickhaare nur ein bis zwei Stränge von Haarmarkzellen. Sie lassen bei der Maus aber eine charakteristische Einschnürung des Haarschaftes zwischen dem dritten und vierten distalen Fünftel des Haares erkennen. Proximal und distal dieses Knickes verjüngt bzw. verdickt sich der Haarschaft schnell. Distal des Knickes erreicht er nicht mehr die maximale Breite des Haarschaftes. Im Bereich der Abknickungen ist kein Haarmark ausgebildet. ad 2.2. Die Zick-Zack-Haare gehören zu der Gruppe der Flaum- oder Wollhaare. Sie bilden den dichten Unterpelz der Mäuse und sind wesentlich feiner und kürzer als die Deckhaare. Zick-Zack-Haare weisen ein nur einreihiges Haarmark auf. Ihr Hauptcharakteristikum ist aber die variable Anzahl von Abknickungen in ihrem Verlauf. Man findet immer mindestens zwei Knicke, es können aber auch bis zu vier Knicke ausgebildet sein. In den Bereichen der Abknickungen flacht sich der Haarschaft deutlich ab und es ist in diesen Bereichen, wie auch bei den Knickhaaren, kein Haarmark vorhanden. Ausführliche morphometrische Untersuchungen der Haarsubtypen der Rumpfbehaarung der Maus wurden bereits von DRY (1926) und UHR (1984) durchgeführt. Ihre übereinstimmenden Ergebnisse werden in Kapitel dargestellt.
34 34 Schrifttum Abbildung 2.7: Schematische Darstellung der vier charakteristischen Subtypen der Rumpfbehaarung der Maus ~ 5 mm Leithaar Ahlenhaar Knickhaar Zick-Zack-Haar Schon 1926 beobachtete DRY, daß die verschiedenen Haartypen entsprechend ihrer unterschiedlichen Größe zu verschiedenen Zeitpunkten entstehen. Die Leithaare werden als die längsten Haare zeitlich vor den Ahlen- und den Knickhaaren gebildet. Die Zick-Zack-Haare werden dagegen erst als letzter Haartyp produziert (MICHEL 1972). Dies legt die Annahme nahe, daß die verschiedenen Haartypen von verschiedenen Haarfollikeltypen gebildet werden, die ebenfalls zeitlich gestaffelt angelegt werden. So sollen die zuerst entstehenden, großen Primärfollikel die Deckhaare produzieren und die feineren Zick-Zack-Haare aus dem Sekundärfollikeln hervorgehen (CLAXTON 1966). Es ist jedoch aufgrund der mit zunehmendem Alter immer geringer werdenden morphologischen Unterschiede nicht möglich, alleine anhand des histologischen Bildes der Haut einer adulten Maus zwischen Primär- und Sekundärfollikeln zu unterscheiden. Eine Bestimmung des von dem jeweiligen Haarfollikel produzierten Haartyps ist im HE- gefärbten Schnitt ebenfalls bei der Maus nicht möglich (SUNDBERG u. HOGAN 1994).
35 Schrifttum 35 Der Mensch besitzt desweiteren während seiner intrauterinen Entwicklung das sogenannte Lanugohaarkleid. Dies besteht aus sehr feinen, marklosen Haaren, welche jedoch schon präpartal wieder abgestoßen werden. Außerdem wird beim Menschen zwischen Vellus- und Terminalhaaren unterschieden. Mit dem Beginn der Pubertät wird in bestimmten Körperregionen (vor allem der Scham, den Achseln und dem Gesicht bei Männern) von der Bildung von feinem Vellushaar zur Bildung der wesentlich größeren und stärker pigmentierten Terminalbehaarung umgeschaltet (STENN et al. 1996) Haarzyklus Das Haarwachstum ist bei den Säugetieren kein kontinuierlicher Vorgang, sondern verläuft in Aktivitätsschüben, die sich immer wiederkehrend und in vollständigen Zyklen wiederholen. Dies beschrieb DRY schon 1926 erstmalig für die Maus. Jeder Haarzyklus besteht aus einer Anagenphase, in der das eigentliche Haarwachstum stattfindet, dem Katagen, der Übergangszeit, in der die Follikelrückbildung erfolgt, und dem Telogen, dem Ruhestadium. Im Anagen, das nach CHASE (1954) beim Menschen in sechs Stadien unterteilt werden kann (I-VI), vergrößert sich der Haarfollikel zunächst durch fortgesetzte Zellteilungen und wächst analog zu seiner embryonalen Entwicklung in die Tiefe. Es wird angenommen, daß dies durch ein Signal der dermalen Papille induziert wird. Die bis dahin ruhenden epithelialen Zellen des Wulstes in der äußeren Wurzelscheide zeigen dabei erneut eine starke Proliferation. In dem Wulst wird daher der Sitz der epithelialen Stammzellen des Haarfollikels vermutet (LAVKER u. SUN 1995). Die dermale Papille wird wieder vom Bulbus umschlossen, die Matrixkeratinozyten proliferieren und differenzieren sich. Es bilden sich, analog zu den Vorgängen bei der embryonalen Haarfollikelentwicklung, wieder eine neue Haarzwiebel und die dazugehörigen inneren Wurzelscheiden. Das aus der neuen Haarzwiebel hervorgehende Haar schiebt sich mit fortschreitender Entwicklung nach distal und verdrängt schließlich das alte Kolbenhaar.
36 36 Schrifttum Abbildung 2.8: Ausgewählte Substadien des murinen Haarzyklus und die charakteristischen morphologischen Veränderungen des Haarfollikels Abkürzungen: BM: verdickte Basalmembran im späten Katagen; DP: dermale Papille; IRS: innere Wurzelscheide; ORS: äußere Wurzelscheide; TD: Talgdrüse * modifiziert nach PAUS (1996)
37 Schrifttum 37 Die vollständige Anagenphase dauert bei der Maus durchschnittlich 17 Tage (RANDALL u. DUSHOFF 1956; SUNDBERG u. KING 1996). Es werden während des späten Anagens durchschnittliche Haarbildungsraten von bis zu einem Millimeter pro Tag bei der Maus erreicht (CHASE 1954). Im Katagen stellen die Matrixkeratinozyten ihre Proliferation ein und differenzieren sich entweder terminal oder fallen der Apoptose anheim (SEIBERG et al. 1995). Dies führt zu einer deutlichen Volumenreduktion des Bulbus und einer erneuten Verkürzung des gesamten Haarfollikels. Der Bulbus wandert wieder in Richtung Hautoberfläche. Bulbus und dermale Papille verlieren dabei kurzzeitig den direkten Kontakt, bleiben aber durch einen schmalen Epithelstrang, der von einer nun verdickten Basalmembran umgeben ist, verbunden. Die gegebenenfalls vorhandene Pigmentproduktion der Melanozyten des Bulbus sistiert und es wird kein Haarmark mehr gebildet. Der Durchmesser des Haares wird dadurch wesentlich verringert. Nach STRAILE et al. (1961) kann die Katagenphase der Maus in acht Substadien unterteilt werden. Das gesamte Katagen dauert bei der Maus nur etwa zwei Tage (PARAKKAL 1970; SUNDBERG u. KING 1996). Während dieser Phase beseitigen Makrophagen die Reste der untergegangenen Zellen des Haarfollikels. Hierauf folgt die Telogenphase, in welcher der Haarfollikel stark verkleinert und vollständig inaktiv ist. Das untere Ende des Ruhefollikels liegt dann auf Höhe des Wulstes. Das Haar selbst nimmt an der Basis im Telogenstadium eine etwas verdickte, kolbige Form mit ausgefranstem Ende an. Es wird zum sogenannten Kolbenhaar, in dem Markstrukturen bei der Maus nur noch supraepidermal erkennbar sind (STRAILE et al. 1961). Da die aufgelöste innere Wurzelscheide nun nicht mehr als Verankerung dienen kann, ist das Haar lediglich durch sein verdicktes Ende, den Kolben, im Follikel verankert (CHASE 1954).
38 38 Schrifttum Ist der erste Haarzyklus nach der embryonalen Entwicklung des Haarfollikels durchlaufen, schließt sich die Ruhephase an. Ist auch diese beendet, erfolgen in bestimmten zeitlichen Abständen mit erneuten Anagen-, Katagen- und Telogenphasen immer wieder die gleichen Auf- und Abbauprozesse an demselben Haarfollikel. Der Zyklus eines Haarfollikels ist daher konzeptionell mit der Ziffer sechs vergleichbar. Ihr freier Arm stellt den Beginn der embryonalen Entwicklung des Follikels dar, während der Bauch die immer wiederkehrenden Zyklen veranschaulicht. Der Follikel unterliegt dabei wahrscheinlich der Regulation durch die selben Wachstumsfaktoren und Steuerungsmechanismen, welche lediglich in geringem Umfang in ihrem zeitlichen Verlauf variieren können (STENN u. EILERTSEN 1996) (siehe Abbildung 2.9).
39 Schrifttum 39 Abbildung 2.9: Konzeptionelles Modell des Haarwachstumszyklus * * modifiziert nach STENN et al. (1996)
40 40 Schrifttum Haarwechsel Bei der Geburt ist der Körper der Maus nackt. Lediglich die Sinushaare sind bereits im Bereich der Schnauze deutlich erkennbar. Andere Haare erscheinen am zweiten bis dritten Lebenstag auf der Hautoberfläche. Zuerst werden sie an Kopf, Nacken, Rücken, Ohren sowie an der Schwanzbasis sichtbar (BORUM 1954). Bis zum 14. postnatalen Tag hat das Jugendfell der Maus seine endgültige Länge erreicht. Zwischen dem 12. und 30. Tag setzt mit dem Beginn der Bildung der zweiten Haargeneration der Haarwechsel ein (SLEE 1962; SUNDBERG u. KING 1996). Hierbei treten die Ruhefollikel wieder in die Phase der Aktivität (Anagen) ein. Im Gegensatz zum Menschen, Meerschweinchen, Hund und Katze, ist bei der Maus ein hochgradig synchronisierter bzw. wellenförmiger Haarwachstumszyklus vorherrschend. Bei ihm treten Gruppen benachbarter Follikel von kranial nach kaudal laufend, gleichzeitig in den Haarwechsel ein (DRY 1926). Weitergehend werden von KRYLTZOV (1964) bei der Maus noch fünf individuelle Haarwachstumstypen unterschieden (Typ I - V). Von diesen findet sich der sogenannte sublaterale Wechseltyp bei der Maus am häufigsten. Bei ihm wird das Fell von den Schultern ausgehend in kraniokaudaler und dorso-ventraler Richtung gewechselt. Mit dem Durchlaufen einer Anagenwelle ist zu beobachten, wie Epidermis, Dermis und Subkutis deutlich dicker und im Katagen wieder dünner werden. Auch Größen und Enzymaktivitäten der Talgdrüsen schwanken Haarzyklusabhängig (PAUS et al. 1994). Bei der Maus besitzen die Talgdrüsen ihre größte Ausdehnung während des Telogens.
41 Schrifttum Regulation des Haarwachstums Systemische und intrinsische Haarzyklusregulation Es wird heute angenommen, daß jeder Haarfollikel einen eigenen Wachstumsrhythmus besitzt, der jedoch durch exogene Stimuli modifiziert werden kann. Solch ein Stimulus kann zum Beispiel die wechselnde Tageslichtlänge sein. Auch endogene, systemische Faktoren, wie schwankende Sexualhormonkonzentrationen, beeinflussen den Haarzyklus. Durch diese systemischen Faktoren wird in erster Linie der Haarzyklus benachbarter Haarfollikel synchronisiert (MESSENGER 1993). Neben den systemischen Faktoren ist eine Vielzahl lokaler Faktoren bekannt, die ebenfalls an der Regulation des Haarwachstumsrhythmus beteiligt sind. Der die Entwicklung der Haarfollikel von neugeborenen Mäusen verzögernde Epidermal growth factor (EGF) war der erste Wachstumsfaktor, der mit der Haarentwicklung in Zusammenhang gebracht wurde (MOORE et al. 1981). Weiterhin wird eine Beteiligung der Mitglieder der Transforming growth factor - β-familie (TGFβ), sowie TGFα, dem Fibroblast growth factor (FGF), dem Keratinocyte growth factor (KGF), dem Insulin-like growth factor (IGF), dem Nerve growth factor (NGF), dem Platelet derived growth factor (PDGF) und dem Bone morphogenic proteine (BMP) angenommen. Auch verschiedene Zytokine, Interferone, der Tumor necrosis factor (TNF), der Colony stimulating factor (CSF), Zell-Adhäsions-Moleküle (Cadherine, Tenascin) und extrazelluläre Matrixkomponenten (Epimorphin, Notch und Retinoide) spielen eine bedeutende, wenn auch noch nicht exakt zu definierende Rolle bei der Haarwachstumsregulation (MOORE et al. 1991; HARDY 1992; HOLBROOK et al. 1993; MESSENGER 1993; DuCROS 1995; PAUS 1996; STENN et al. 1996; TAKAKURA et al. 1996). Obwohl das eigentliche induktive Signal für den Übergang ins Anagen noch nicht bekannt ist, ist ein deutlich verändertes Profil der Wachstumsfaktoren in der gesamten Haut während des späten Anagens und des frühen Katagens nachweisbar (STENN et al. 1994; SEIBERG et al. 1995).
42 42 Schrifttum Tabelle 2.2: An der Kontrolle der Haarentwicklung und des Haarzyklus beteiligte Moleküle Molekül Lokalisation Referenz bcl2 A-Bulbus A-T DP Stenn et al 1994 c-fos Kutikula, Rinde, Henle Fisher et al EGF ORS DuCros, 1995 EGF-R ORS, Matrix Green u. Couchman, 1984 FGF-R1 A-DP Peters et al FGF-R2 A-Matrix Peters et al. 1992; Danilenko et al FGF-R3 Kutikula, A-Matrix Rosenquist u. Martin, 1996 FGF-R4 Bulbus, IRS, ORS Rosenquist u. Martin, 1996 FGF-5 A-ORS Hebert et al FGF-7 (KGF) DP Rosenquist u. Martin, 1996 HGF DP Shimaoka et al IGF-I DP, Follikelepithel Itami et al. 1995; Hembree et al IGF BP 2, 3 DP Hembree et al IL 1 α, IL 1 β A-Epithel, ORS Harmon et al. 1993; Boehm et al M-Notch Rinde, IRS, Kutikula Kopan u. Weintraub, 1993 NGF-R DP, ORS Holbrook et al Neu gene product ORS Maguire et al N-myc A-Matrix Mugrauer et al N-CAM DP Chuong et al Nexin-1 DP Yu et al PDGF α/β-rezeptor DP Ponten et al PDGF-A/B Matrix Ponten et al PTHrp Follikelepithel, IRS Hayman et al Substanz P-Rezeptor DP Pincelli et al TGFα ORS, IRS, keratogene Zone Luetteke et al TGFβ DP, ORS, IRS, Matrix Little et al TGFβ R Typ II IRS Higley et al TIMP A-Henle Kawabe et al TNF α A-Matrix Boehm et al Versican DP DuCros et al VEGF DP Lachgar et al Vitamin-D-Rezeptor ORS und A-DP Reichrath et al. 1994
43 Schrifttum 43 Abkürzungen Tabelle 2.2 (vorige Seite): A-: Anagen; DP: Dermale Papille; EGF: Epidermal growth factor; FGF: Fibroblast growth factor; KGF: Keratinocyte growth factor; HGF: Hepatocyte growth factor; IGF: Insulin-like growth factor; IL: Interleukin; IRS: Innere Wurzelscheide; K: Katagen; N-cam: Nerve-cell adhesion molecule ; NGF: Nerve growth factor; ORS: Äußere Wurzelscheide; PDGF: Platelet derived growth factor; PTHrp: Parathyroid hormone-related protein; -R: Rezeptor; TGF: Transforming growth factor; TIMP: Tissue inhibitor of metalloproteinase; TNF: Tumor necrosis factor; VEGF: Vascular endothelial growth factor; Tabelle 2.3: Endogene Haarwachstumsmodulatoren Hormone / Neuropeptide Androgene Östrogene Thyroxin Prolaktin Melatonin Retinoide Vitamin D3 Glukokortikoide ACTH (Adreno corticotropic hormone) α-msh (Melanin-stimulierendes Hormon) Substance P bekannte Effekte / beteiligt an Ausbildung der Körperbehaarung Katageninhibition Katageninduktion, Anageninduktion Induktion von Fellwechsel Körperbehaarung Anageninduktion und/oder Katageninhibition Hirsutismus und Glatzenbildung bei Frauen Induktion von Fellwechsel Follikelentwicklung Anageninduktion und/oder Katageninhibition Haarzyklusregulation Hemmung zytostatika-induzierter Follikeldystrophie Inhibition der Anagenentwicklung Katageninduktion, Hypertrichose Hypertrichose, Anageninduktion Stimulation der Follikelmelanogenese Anageninduktion
44 44 Schrifttum Der Informationsaustausch zwischen den Zellen des Haarfollikels und der dermalen Papille zur Regulation der Follikelentwicklung und des Haarzyklus kann dabei auf drei verschiedene Arten stattfinden: durch Zell-Zell-Kontakte, durch Zell-Matrix-Interaktionen und über Interaktionen von Geweben mit Nerven. Der genaue Mechanismus der Informationsübertragung bei der Entstehung des Haarfollikels ist jedoch noch weitgehend unbekannt (HOLBROOK et al. 1993). Die Bedeutung der Morphogene für die Haarentwicklung wird durch die Verwendung von Mausmutanten im Tierversuch deutlich. TGFα-Knockout- Mäuse zeigen beispielsweise einen abnormen Hautaufbau, ein dünnes, gewelltes Haarkleid (einschließlich der Vibrissen) und leiden unter entzündlichen Augenveränderungen. Sie gleichen somit der seit langem bekannten Waved-Mutante (Tgfα wa-1 ) (MANN et al. 1993; LUETTEKE et al. 1994). Andererseits kann durch Injektionen von Wachstumsfaktoren ein Haarwachstum bei minderbehaarten Mausmutanten (HNF3/forkhead homolog 11, Hfh11 nu ) erzeugt werden (DANILENKO et al. 1995). Bei der Maus sind zumindest zwei Phasen der Anagenentwicklung bekannt. Man unterscheidet zwischen einer nicht Kortikosteroid-sensitiven und eine darauffolgenden, hoch Kortikosteroid-sensitiven Phase. Dies kann der Erforschung der ersten anageninduzierenden Signale und ihrer Gene dienen (STENN et al. 1993; PAUS et al. 1994).
45 Schrifttum Aktuelle Modelle der Haarzyklusregulation Zur Zeit existieren zwei Modelle für die Zykluskontrolle des Haarfollikels. Die sogenannte Chalonhypothese postuliert, daß es während des Anagens zu einer langsamen Akkumulation von endogenen Mitoseinhibitoren (Chalonen) kommt, die ab einem gewissen Schwellenwert das Anagen beenden. Ein langsamer Aktivitätsverlust dieser Chalone während des Telogens desinhibiert demnach die Matrixzellen des Follikels, so daß er erneut ins Anagen eintritt (MESSENGER 1993). Die derzeit favorisierte Wulstaktivationshypothese erklärt den Eintritt in die verschiedenen Phasen dagegen über die wechselnde Aktivität der Follikelstammzellen in der Wulstregion. Auf ein unbekanntes Signal hin schicken diese schnell proliferierende Tochterzellgenerationen zum Aufbau des Anagenbulbus in Richtung dermaler Papille. Wenn die genetisch determinierte, endliche Zahl von Mitosen, die diese transient amplifiying cells ( sich vorübergehend teilende Zellen ) durchlaufen können, stattgefunden haben, führt dies zur Regression des proximalen Bulbus (COTSARELIS et al. 1990; SUN et al. 1991). Beide Hypothesen lassen viele Aspekte ungeklärt. So läßt die Wulstaktivationshypothese beispielsweise offen, woher die Fibroblasten der dermalen Papille wissen, wann die Anagen-induzierenden Substanzen in Richtung Wulst zu sezernieren sind, welche dies überhaupt sind und warum dies rhythmisch geschieht (PAUS 1996). Was können die Gründe für Mitoseinhibitoren sein, die Keratinozyten in die terminale Differenzierung bzw. Apoptose zu treiben, wenn man der Chalonhypothese folgt? Außerdem bleiben auch die Abschaltung der Melanogenese und die Aktivitäten der Makrophagen unerklärt (PAUS et al. 1994).
46 46 Schrifttum 2.3 Behaarungsdefekte der Maus Erste Beschreibungen haarloser Mäuse finden sich bereits im 19. Jahrhundert (GASKOIN 1856). Auch bei zahlreichen anderen Säugetierspezies sind Minderbehaarungen bekannt, darunter finden sich das Rind (OLSON et al. 1985; MEYER et al. 1992; STÖBER et al. 1995), der Hund (MARKS et al. 1992; IHRKE et al. 1993), die Katze (CASAL et al. 1994), das Schaf (MACKIE u. McINTYRE 1993), der Affe (RATTERREE u. BASKIN 1993) und die Ratte (MARIT et al. 1995). Auch beim Geflügel treten ähnliche Veränderungen in graduell verschiedener Ausprägung bis hin zu einer allgemeinen Federlosigkeit auf (PECH-WAFFENSCHMIDT et al. 1995) Mausmodelle Mäuse mit erblichen Haut- und Haardefekten sind wichtige Mittel zur Erforschung humaner und veterinärmedizinischer Erkrankungen. Sie tragen wesentlich zum besseren Verständnis der Haarentwicklung (SUNDBERG 1994; PAUS et al. 1994), der Pathogenese von mit Alopezien einhergehenden Hauterkrankungen (SUNDBERG et al. 1990; SUNDBERG u. SHULTZ 1991; SUNDBERG et al. 1995; MEISLER 1996) sowie zur Erprobung neuer Therapien bei (SUNDBERG u. KING 1996). Abgesehen von Mausmodellen stehen der Forschung auch Organ- oder Zellkulturen zur Verfügung. Es ist bisher nicht möglich, mit Hilfe von In-vitro- Modellen alle Transformationen, die der Haarfollikel während eines Zyklus durchläuft, nachzuvollziehen. Daher stellen In-vitro-Modelle eine nur sehr begrenzte Hilfe bei der Erforschung der zyklischen Vorgänge des Haarfollikels und ihrer auslösenden Signale dar (PAUS 1996). Außerdem liegen bis heute keine Beweise dafür vor, daß bedeutende Unterschiede in der Regulation der sogenannten Haarzyklus-Uhr zwischen den verschiedenen Säugetier- Spezies existieren (PAUS 1996). Forschungsergebnisse, die durch die Verwendung von Mausmodellen entstehen, sind daher bedingt auch auf andere Tierarten und den Menschen übertragbar.
47 Schrifttum 47 Da das Mausgenom im Unterschied zu den Erbanlagen der Haustiere weitestgehend bekannt ist, eignet sich diese Spezies besonders zur Klärung der genetischen Ursache solcher Defekte. Zudem weist das Mausgenom weitreichende Übereinstimmung mit dem menschlichen Genom auf (MEISLER 1996). Daher kann die Verwendung von Mausmodellen, die genetische Defekte aufweisen, der Analyse des murinen und somit auch des humanen Genoms dienen. Das Fernziel stellt dabei die vollständige Kartierung des menschlichen Genoms im Rahmen des sogenannten Human Genome Project dar (KOPALA 1997). Neben ihrer geringen Größe und der damit verbundenen einfachen und kostengünstigen Haltung und Reproduktion weisen bestimmte Mausstämme zudem interessante Merkmale auf. Beispielsweise lassen pigmentierte Mausstämme aufgrund ihrer Hautfärbung eine Diagnose des momentanen Haarzyklusstandes alleine durch makroskopische Betrachtung zu. Eine einfache Überprüfung des Einflusses von Stoffen auf den Haarzyklus ist damit möglich. So färbt sich die Haut des C57BL/6-Stammes bei Anagenbeginn dunkel und verblaßt zu Beginn der Katagenphase (STENN et al. 1996) Genetisch determinierte Behaarungsdefekte der Maus Im Schriftum wird über eine große Zahl verschiedener Mausstämme berichtet, die Defekte ihrer Behaarung in unterschiedlichster Ausprägung aufweisen (SUNDBERG 1994, HOGAN et al. 1995; LYON et al. 1996; SUNDBERG u. KING 1996). Tiere mit Behaarungsdefekten können durch spontane Mutationen entstehen. Die Träger des Gens können dann durch selektive Nachzucht vermehrt werden. Das Auftreten von Mutationen kann aber auch gezielt durch die Anwendung von Mutagenen oder durch Kreuzungen von bereits bekannten Mutationen untereinander provoziert werden. Von besonderer Bedeutung ist dabei der genetische Hintergrund der Anlagenträger, da die Mutation des gleichen Gens bei Tieren verschiedener Stämme aufgrund von Interaktionen unterschiedliche Auswirkungen haben kann. Vor allem transgene Tiere tragen heute erheblich zum Verständnis der Haarentwicklung bei (STENN et al. 1996).
48 48 Schrifttum Bei transgenen Tieren wird die Expression eines Gens gezielt hochreguliert. Im Gegensatz dazu wird bei Knock-out-Mäusen ein Gen durch Mutation vollständig inhibiert. Dadurch konnte beispielsweise gezeigt werden, daß die angora-mutation (go) identisch mit der Null-Mutation des FGF5-Gens (Fgf5) ist (HEBERT et al. 1994; MEISLER 1996) Ausprägung der Defekte Die jeweiligen Veränderungen können in unterschiedlich starker Ausprägung vorhanden sein. Sie betreffen bei dominant-rezessiven Erbgängen nur die homozygoten Merkmalsträger eines rezessiven Allels, oder die heterozygoten Träger eines dominanten Allels. Bei intermediären Erbgängen kann es zu einer geringeren Ausprägung des Merkmals bei den heterozygoten Tieren kommen, wie z. B. bei der Mutante naked. [Im Folgenden werden beispielhaft entsprechende Mausmutanten in Klammern genannt. Die Referenzen aller Mutationen sind in der Tabelle 2.4 aufgeführt.]. Gleiches gilt für das zeitliche Auftreten von Defekten. Sie können von Geburt an präsent sein oder aber erst im fortgeschrittenen Lebensalter auftreten. Möglich ist zum Beispiel, daß erste Haarzyklen ohne Besonderheiten durchlaufen werden, die Haare dann aber ausfallen und nicht wieder nachwachsen (atrichosis, bare skin, depilated). Ebenso ist aber auch der umgekehrte Fall möglich. Nachfolgende Haarzyklen können normal ablaufen, obwohl während des ersten Zyklus noch Defekte existent waren. Bei den meisten Mutationen betreffen die Veränderungen das gesamte Tier. Es sind aber auch Defekte bekannt, die lokal begrenzt sind (hairy ears, hair patches). Sie können für das Tier nahezu bedeutungslos sein (silver) und so dezent ausfallen, daß sie makroskopisch nicht sichtbar sind (Markmißbildungen bei hair interior defect). Die Veränderungen können aber auch so schwerwiegend sein, daß sie zum Tod des Individuums führen. Eine eingeschränkte Vitalität der Tiere wird häufig beobachtet, wenn nicht nur die Haare und/oder die Haut, sondern auch innere Organe des Tieres verändert sind (Herz und Nieren bei hair patches, Herz bei lanceolate).
49 Schrifttum 49 Bekanntestes Beispiel dürfte hierfür die Thymuslosigkeit und die daraus resultierende Immundefizienz der Nacktmaus sein (HNF-3/forkhead homolog 11, ehemals nude ). Meist betrifft die Vergesellschaftung mit anderen Organsystemen aber lediglich verwandte Strukturen, wie Nägel oder Zähne (naked, shaven, tabby) mit weniger schwerwiegenden Folgen. Häufig wird bei Mausmutanten mit Behaarungsdefekten eine reduzierte Fruchtbarkeit gefunden (naked, crinkled). In manchen Fällen geht dies auch mit Defekten des äußeren Genitale einher (ichthyosis). Bei der Mutation atrichosis liegen zum Beispiel hypoplastische Gonaden vor, bei hairless ist die Mamma nur rudimentär ausgebildet. Diese Mißbildungen können in der Folge zu einer stark erschwerten Nachzucht der Tiere führen. Die Erzeugung homozygoter Nachkommen kann aber auch ohne die Ausbildung makroskopisch sichtbarer Defekte erschwert oder unmöglich sein. Beispielsweise müssen dann heterozygote Tiere angepaart werden, da die homozygoten Merkmalsträger nicht in der Lage sind, ihre Nachkommen aufzuziehen (HNF-3/forkhead homolog 11). Dabei müssen dann für den eigentlichen Versuch eventuell unerwünschte heterozygote Nachkommen in Kauf genommen werden. Auch der Schwanz ist bei verschiedenen Mutanten mitbetroffen. Entweder liegen Wirbelkörpermißbildungen vor (crinkled) oder es kann zur Autoamputation nach Ringbildungen kommen (ichthyosis). Der Schwanz kann sowohl fehlerhaft behaart als auch nackt sein (tabby, crinkled) Ursachen von Haarwachstumsstörungen Zumeist liegen den Alopezien primäre Haarbildungsstörungen zugrunde (HNF-3/forkhead homolog 11). Es ist aber auch möglich, daß normal gebildete Haare aufgrund einer strukturellen Besonderheit erst nach ihrer Eruption an die Oberfläche durch mechanische Belastungen abbrechen (matted, lanceolate) und dadurch hypotriche Bereiche entstehen. Es können anomale Haare aus morphologisch intakten Follikeln hervorgehen, defekte Haarfollikel können aber auch relativ unauffällige (ragged) oder schwer mißgebildete Haare hervorbringen (harlequin).
50 50 Schrifttum Durch das Fehlen der inneren Wurzelscheide bei der Nacktmaus (HNF-3/forkhead homolog 11) ist nur noch rudimentäres Haarwachstum festzustellen (KOPF-MAIER et al. 1990). Häufig verhindert auch eine Desorientierung der Follikel in der Dermis einen Durchbruch eines gebildeten Haares an die Hautoberfläche (depilated, shaven, ragged). Auch Verhornungsstörungen wie eine Akanthose oder eine Follikelkeratose können dies bewirken (harlequin, hairless). Andererseits können primäre entzündliche Alterationen der Haut aber auch in der Folge alopezische Areale nach sich ziehen (flaky skin). Bei wenigen Mutationen betreffen die Veränderungen auch nur einen Teil der vier Haartypen des Rumpfes, eventuell einschließlich der Vibrissen (desmoglein 3, fuzzy). Bei der Mutation frizzy sind beispielsweise vorwiegend Zick-Zack-Haare ausgebildet, die bei crinkled und tabby völlig fehlen. Den tabby-mäusen fehlen außerdem die Leithaare, die Mutationen shaven und caracul besitzen darüberhinaus keine Knickhaare. Es überwiegen im allgemeinen bei der Maus Haarbildungsstörungen, die eine verminderte Behaarung zur Folge haben. Wie bei anderen Tierarten existieren aber ebenso Mutationen, die eine Hypertrichose zeigen (angora, hairy ears) Beteiligung der Adnexe an Haarwachstumsstörungen Oft sind bei Haarfollikelmißbildungen die adnexalen Strukturen mitbetroffen, so daß daraus sekundäre Veränderungen der Haut resultieren (asebia, bare skin, hairless). Betreffen die Mißbildungen auch die adnexalen Strukturen der Haarfollikel der Augenlider, finden sich häufig Schädigungen der Augen selbst in Form kornealer Läsionen (Agenesie der Meibomschen Drüsen bei tabby, fehlende Hardersche Drüsen bei ichthyosis). Eine Übersicht über die große Variabilität bekannter Haardefekte von Mäusen soll die folgende Tabelle geben.
51 Schrifttum 51 Tabelle 2.4: Auswahl von Mausmutationen mit Behaarungsdefekten* Name der Mutante Gensymbol Chromosom Organmißbildungen Haarlängenabweichung Zelldebris/Haut tordierte Haare/Follikel alopecia areata?? + + Sundberg et al angora Fgf5 5 + (+) Pennycuik et al asebia ab Pennycuik et al atrichosis at Sundberg 1994 bare skin Bsk Sundberg 1994 caracul Ca Sundberg 1994 crinkled cr Mayer et al depilated dep 4 + Mayer et al desmoglein 3 Dsg Sundberg 1994 flaky skin fsn Sundberg 1994 frizzy fr Falconer u. Snell 1954 fuzzy fz 1 + Mayer et al hairpatches Hpt Shultz et al hairy ears Eh 15 + Davisson et al hair interior defect hid? Trigg 1972 hairless hr Mann 1971 harlequin Hq X + + Sundberg 1994 ichthyosis ic Sundberg 1994 lanceolate lah Montagutelli et al matted ma Sundberg 1994 naked N Pennycuik et al HNF-3 / forkhead homolog 11 Hfh Flanagan 1966 ragged Ra Sundberg 1994 shaven Sha 15 (+) Sundberg 1994 silver si 10 + Sundberg 1994 tabby Ta X + + Pennycuik et al Abkürzungen: X: Gonosom; +: vorhanden; (+): teilweise vorhanden;?: unbekannt * erweitert und modifiziert nach HOGAN et al und SUNDBERG u. KING 1996 Kutikuladefekte Brüche der Haare fehlende Haartypen Verlust der Haare Referenz
52 52 Schrifttum 2.4 Hypotrichosen des Menschen Wie bei den Tieren ist auch beim Menschen eine Vielzahl unterschiedlicher Behaarungsdefekte bekannt. Neben Hypomelanosen und Hypertrichosen (Hirsutismus) überwiegen auch in der Humanmedizin zahlenmäßig Störungen, die mit einer Hypotrichose einhergehen. Hierbei werden wenige bis gar keine Haare an Rumpf und/oder Kopf ausgebildet (ROOK 1973; ORFANOS 1991; ROOK u. DAWBER 1995). Dies kann durch lokale oder systemische Infektionen, durch Aufnahme von Chemikalien oder durch Störungen des Hormonhaushaltes oder des Stoffwechsels hervorgerufen werden. Der Mangel an bestimmten Spurenelementen, wie zum Beispiel Kupfer, kann ebenfalls Minderbehaarungen verursachen. Angeborene genetische Defekte spielen jedoch beim Menschen ätiologisch ebenso eine bedeutsame Rolle. Kongenitale Mißbildungen können entweder idiopathisch auftreten, oder aber als Nebenbefund bei den verschiedensten Syndromen beobachtet werden. Entsprechend der Vielzahl der mit Behaarungsstörungen vergesellschafteten Syndrome existiert auch eine sehr große Variabilität der bei diesen Erkrankungen vorliegenden pathologischen Veränderungen. Die Defekte, die dabei an den Haaren selber beobachtet werden können, unterteilt man nach ihren klinischen Auswirkungen, wie in Tabelle 2.5 dargestellt. Abkürzungen Tabelle 2.5: + monogener Erbfehler; (folgende Seite) (+) wahrscheinlich erblich disponiert;? erbliche Disposition unklar; - nach bisherigen Erkenntnissen nicht erblich disponiert; zunehmend abnehmend
53 Schrifttum 53 Tabelle 2.5: Veränderungen des Haarschaftes des Menschen 1) mit erhöhter Brüchigkeit Vererbung Symptomatik Monilethrix + spindelförmige Internodien Pseudomonilethrix + Verdickungen, Torsionen, Brüche Pili torti + Torsionen, Abflachungen, Brüche Kinky-hair-syndrome (Menkes Syndrom) + Brüche Bambushaare (Netherton syndrome) + invaginierte Knoten u. Brüche Trichorrhexis nodosa? Verdickungen, Brüche Trichothiodystrophie + Verwitterung, Brüche, Abflachung, Furchungen, Fältelungen 2) ohne erhöhte Brüchigkeit Pili anulati (Ringelhaare) + Bänderung halbierte Wachstumsrate Wollhaare + stark gekrümmte Haare, Durchmesser, Pili torti/anulati, Trichorrhexis nodosa APK (Aquired progressive kinking) (+) Abknickungen, Drehungen, Durchmesser, Verkürzung der Anagenphase Spun-glass-hair (unkämmbare Haare) (+) dreieckige Schaftquerschnitte Loose-anagen-hair-syndrome (+) Haare verbleiben im Anagenstadium 3) andere Veränderungen und sind leicht ausziehbar Trichoklasie - Grünholzfraktur Trichoptilosis - Haarspliss Rollhaare (Pili incarnati) - in die Haut einwachsende Haare Trichomalazie - weiche, deformierte, verdickte Haare mit tordierten Wurzeln Trichoschisis - Querbrüche Pohl-Pinkus-Mark - Durchmesser, diskontinuierl. Mark Ausrufungszeichen-Haare - Durchmesser (Hantelform) Trichonodosis - Knoten, Risse, Brüche Trichostasis spinulosa - Vellushaare in Hornpfropfen Pili multigemini + multiple Matrizes und Papillen in einem Follikel Verwitterung - Zerfall des gesamten Schaftes
54 54 Schrifttum Man unterscheidet dabei Veränderungen, die mit einer erhöhten Fragilität des Haarschaftes einhergehen (Gruppe 1) von Erkrankungen, die dieses Phänomen nicht aufweisen (ROOK u. DAWBER 1995). Ursache für eine erhöhte Brüchigkeit sind dabei meist Zu- oder Abnahmen des Haarschaftdurchmessers. Diese können sogar pathognomonisch sein, wie zum Beispiel die bei dem Bambushaar des sogenannten Netherton Syndromes vorliegenden invaginierten Knoten, oder die spindelförmigen Internodien bei Monilethrix (siehe Tabelle 2.5). Die mit erhöhter Brüchigkeit der Haarfaser einhergehenden Erkrankungen sind zumeist hereditär bedingt (STROUD 1987). Die Erkrankungen der zweiten Gruppen zeigen anstelle der Brüche zumeist eine Aufrollung oder Verdrehung der Haare und sind meist nicht (nur) genetisch disponiert. In der dritten Gruppe finden sich ebenfalls zumeist nicht erbliche Defekte des Haares, die keiner der anderen Gruppen zuzuordnen sind und die mit verschiedensten Syndromen vergesellschaftet auftreten können (ORFANOS 1991). Neben den aufgeführten physischen Veränderungen können beim Menschen auch Verhaltensstörungen die Ursache für eine Minderbehaarung sein. Das Ausreißen der eigenen Kopfbehaarung, die nicht ursächlich verändert war, wird humanmedizinisch als Trichotillomanie bezeichnet (ROOK 1973) Alopezien Von größerem humanmedizinischem Interesse sind außer den vorangehend beschriebenen Behaarungsdefekten die verschiedenen Arten der Alopezien. Bei diesen kommt es im wesentlichen nicht zu einer Veränderung des Haarschaftes, sondern zu einer Verminderung der Anzahl der Haare. Ebenso wie bei den Hypotrichosen können von den Alopezien sowohl Männer als auch Frauen betroffen sein. Die Pathogenesen der meisten Alopezien sind bis heute immer noch nicht vollständig geklärt. Man unterscheidet vier Gruppen von Alopezien:
55 Schrifttum 55 Bei der sogenannten androgenetischen Alopezie kommt es zu einer fortschreitenden Degeneration des bindegewebigen Haarbalgs und in dessen Folge zu einer Rückbildung und Miniaturisierung der Follikel. Die ersten nachweisbaren Veränderungen sind eine fokale perivaskuläre basophile Degeneration der bindegewebigen Wurzelscheide der ansonsten normal erscheinenden anagenen Follikel. Die betroffenen Haarfollikel werden im Laufe weiterer Haarzyklen immer kleiner. Es werden keine Terminalhaare, sondern nur noch Vellushaare gebildet. Es sind häufig mehrkernige Riesenzellen in der Umgebung von Haarfragmenten zu finden. Einzelne ruhende Terminalhaarfollikel können bestehen bleiben und selten ist es möglich, diese zu neuem Wachstum zu stimulieren, was die falsche Hoffnung auf eine Heilung der Alopezie mit sich bringen kann. Da die Prävalenz der androgenetischen Alopezie bei männlichen Mitteleuropäern nahezu 100 Prozent beträgt und nur der Grad der Ausprägung variiert, muß der Ersatz von Terminalhaarfollikeln durch solche des Vellustyps als normaler Vorgang angesehen werden (ROOK u. DAWBER 1995). Aufgrund der großen Variabilität der androgenetischen Alopezie und ihrer schwerwiegenden kosmetischen Bedeutung für den Patienten, existiert ein großes wissenschaftliches und wirtschaftliches Interesse an der Aufklärung ihrer Pathogenese (SUNDBERG et al. 1995). Die diffuse Alopezie ist durch einen fokal begrenzten oder diffus verteilten Eintritt von Haarfollikeln in die Katagenphase gekennzeichnet. Dies hat in zwei bis drei Monaten, also nach Beendigung des folgenden anagenen Stadiums, einen über die Norm gesteigerten Haarausfall der nun telogenen Follikel zur Folge. Diese Art von Alopezie stellt die Reaktion der Haarfollikel auf einen unbekannten Reiz dar. Es sind bis heute viele mögliche Ursachen bekannt, die Bandbreite reicht von psychischem Stress über systemische Erkrankungen oder Mangelzustände bis zu physischen Traumata. Auch durch die Geburt eines Kindes kann es zum sogenannten telogenen Effluvium kommen. Zumeist sind diese Alopezieformen jedoch reversibel.
56 56 Schrifttum Bei der Alopezia areata des Menschen kommt es nach ECKERT et al. (1968) zu einem vorzeitigen, wellenförmig fortschreitenden Übergang der Follikel in die Telogenphase, der entweder fokal begrenzt oder aber diffus ausgebreitet ist. Es können alle Haarfollikeltypen des gesamten Körpers betroffen sein. Stets kann peribulbär und intrafollikulär ein lymphozytäres Infiltrat histologisch nachgewiesen werden. Eine Narbenbildung ist meist jedoch nicht vorhanden. Es werden Autoantikörper gegen die betroffenen dystrophischen Haarfollikel gebildet. Pathognomonisch hierfür sind die sogenannten Ausrufungszeichenhaare, die allerdings nicht immer vorhanden sein müssen. Sie werden als Kolbenhaare von normalem Kaliber und Pigmentierung mit etwa drei Millimetern Länge angesehen. Ihre distalen Enden sind gegebenenfalls abgebrochen und unregelmäßig ausgefranst. Nach proximal nimmt ihr Durchmesser nur sehr langsam ab, um in einem kleinen, aber normalen Kolben zu enden (ROOK u. DAWBER 1995; SUNDBERG et al. 1995). Unter dem Begriff vernarbende Alopezien werden diejenigen Alopezieformen zusammengefaßt, die mit einer Zerstörung von Haarfollikeln einhergehen bzw. diesen folgen. Die Zuordnung zu dieser Gruppe erfolgt unabhängig davon, ob die Follikel selbst betroffen sind, oder ob der Krankheitsprozeß extrafollikulär abläuft. Die Follikel können als Folge eines entwicklungsbedingten Prozesses fehlen oder durch ein Trauma oder eine spezifische Infektion (Favus, Tuberkulose, Syphilis) irreparabel zerstört sein. Eine große Zahl verschiedener Ätiologien führen zu der Diagnose einer vernarbenden Alopezie, wobei die Pathogenese in vielen Fällen ungeklärt ist. Einer Infiltration mit Entzündungszellen folgt dabei die Zerstörung der mit Hornpfröpfen angefüllten Follikel und die Ausbildung einer dünnen Epidermis über einer sklerotischen Dermis.
57 Eigene Untersuchungen 57 3 Eigene Untersuchungen 3.1 Material und Methoden Tiermaterial Seit mehr als 20 Jahren werden im Institut für Grundlagen der Nutztierwissenschaften der Berliner Humboldt Universität in Reinzucht verschiedene Mäuse-Inzuchtlinien gehalten traten spontan in zwei Würfen des seit mehr als 20 Generationen ingezüchteten Albino-Stammes HDA32/Thb getrennt voneinander hypotriche Tiere auf. Neben diesen minderbehaarten Tieren waren in diesen beiden Würfen aber auch normal behaarte Vollgeschwister vorhanden. Durch weitere Anpaarungen konnte gezeigt werden, daß es sich bei dem Defekt um einen einfachen, autosomalrezessiven Erbgang handelt. Insgesamt wurden etwa 250 Anpaarungen mit hypotrichen Tieren und Tieren des Ursprungsstammes HDA32/Thb vorgenommen (SCHMIDT u. SCHLOTE 1995). Das Gen der hypotrichen Tiere wurde vorläufig als sht, für short -haired, registriert. Die Gensymbole werden allgemein kursiv geschrieben. Da es sich um einen rezessiven Erbgang handelt, wird der Anfangsbuchstabe des Gensymbols in diesem Fall klein geschrieben. Der Genotyp ist hierbei anhand des Phänotyps eindeutig definierbar. Homozygote Tiere sind von der Geburt an phänotypisch anhand der feinen und gewellten Schnurrhaare und der gleichmäßig über den ganzen Rumpf ausgebildeten Hypotrichose zu identifizieren. Tiere des sht/sht Genotyps werden im folgenden synonym auch als hypotrich oder minderbehaart bezeichnet. Den heterozygoten Genotyp kennzeichnet der international üblichen Nomenklatur folgend (LYON et al. 1996; SUNDBERG u. KING 1996) ein + ( Wildtyp ). Dieser Genotyp wird im folgenden als normal behaart oder +/sht bezeichnet. Der heterozygote Anlageträger besitzt ein normal ausgebildetes Fell und ist phänotypisch von den Tieren des Ursprungsstammes HDA32/Thb ( +/+ ) nicht zu unterscheiden.
58 58 Eigene Untersuchungen Es erfolgte eine selektive Nachzucht der minderbehaarten Tiere durch reine Bruder-Schwester-Anpaarungen. Zunächst fand dies an der Berliner Humboldt Universität statt. Im April 1995 wurde ein Zuchtkern, bestehend aus drei gemischten Würfen aus reziproken Anpaarungen (sht/sht X +/sht), von Berlin in die Tierärztliche Hochschule Hannover verbracht. Zusätzlich fand eine weitere Vermehrung von zwei Würfen der Tiere der Ursprungslinie HDA32/Thb zur Kontrolle in Hannover statt Haltungsbedingungen Die Haltung der Tiere erfolgte unter den für Labormäuse konventionellen Bedingungen im Institut für Tierzucht der Tierärztlichen Hochschule Hannover. Es wurden Macrolonkäfige der Größe II (360 cm 2 ) mit Drahtdeckeln verwendet, in denen jeweils bis zu drei Tiere auf Holzgranulat (Altromin animal bedding granular) gehalten wurden. Die Raumtemperatur betrug C bei einer relativen Luftfeuchte von 40 bis 65%. Die Mäuse hatten 12 Stunden Licht pro Tag (Deckenbeleuchtung mit Neonlampen durch eine Zeitschaltuhr gesteuert) und freien Zugang zu Wasser (Macrolonflaschen) sowie Futterpellets (Altromin 1324) ad libitum. Ab dem Alter von 6 Wochen wurden Anpaarungen vorgenommen, wobei jeweils einem Männchen nur ein Weibchen zugesetzt wurde. Bei erkennbarer Trächtigkeit wurden die Paare getrennt. Um mögliche Wurfeffekte zu minimieren fand bei größeren Würfen gegebenenfallls eine Reduzierung auf acht Nachkommen pro Wurf statt. Mit 21 Tagen wurden die Jungtiere von ihren Müttern getrennt. Es erfolgte zudem eine Auftrennung nach dem Geschlecht der Tiere. Durch Anpaarung von homozygot-minderbehaarten Mäusen (sht/sht) an heterozygot-normalbehaarte Tiere (+/sht) wurde eine Aufteilung von etwa 1:1 auf beide Genotypen in den Würfen erreicht. Es erfolgte also eine Erzeugung beider Genotypen innerhalb eines Wurfes von einer Mutter. Aufgrund der daraus resultierenden identischen Wurfumwelt können sowohl Umwelt-, als auch maternale Einflüsse, die zu Unterschieden zwischen den beiden Genotypen führen könnten, vernachlässigt werden. Diese Nachkommen standen für die im folgenden beschriebenen Untersuchungen zur Verfügung.
59 Eigene Untersuchungen Untersuchungsmethoden Umfang der Untersuchungen Die Planung der Sektionen und der Probennahmen von Haut und Organen richtete sich in ihrem Umfang und dem Zeitrahmen nach der Vorgehensweise des Jackson Laboratory (Maine, USA) (HOGAN 1995). Es wurden während des ersten Haarzyklus (0-21 Tage) alle drei Tage von jeweils zwei männlichen und zwei weiblichen sht/sht Tieren Hautproben entnommen. Ebenso wurden jeweils zwei männlichen und zwei weiblichen +/sht Tieren die gleiche Anzahl von Hautproben zu den selben Altersstufen entnommen. Außer der Hautprobenentnahme wurde bei den selben Tieren der Altersstufen 0, 21, 42 und 182 Lebenstagen zusätzlich eine vollständige Sektion durchgeführt. Pro Lebenstag wurden also immer acht Tiere untersucht. Bei insgesamt 10 Untersuchungszeitpunkten (Lebenstagen) ergibt dies eine Gesamtsumme von 80 untersuchten Mäusen (siehe Abbildung 3.1). Nach Möglichkeit waren die Tiere der verschiedenen Genotypen an einem Untersuchungstag gleich alte Vollgeschwister. Maximal kamen je Untersuchungstag Tiere aus zwei verschiedenen Würfen zur Untersuchung. Bei den Tieren der letzten Altersstufe (Lebenstag 182) wurde aus praktischen Gründen eine Toleranz von +/- fünf Tagen akzeptiert. Die Tiere wurden zuvor durch Genickbruch getötet, da diese Methode wegen der damit einhergehenden Schmerzfreiheit für die Tiere und die schnelle Durchführbarkeit speziell bei Mäusen empfohlen wird (FLECKNELL 1995; CLOSE et al. 1997).
60 60 Eigene Untersuchungen Abbildung 3.1: Zeitplan der Probennahme und Anzahl der verwendeten Tiere Lebensalter in Tagen: Genotyp Anzahl Sex sht/sht 2 S H H H H H H H/S H/S H/S sht/sht 2 S H H H H H H H/S H/S H/S +/sht 2 S H H H H H H H/S H/S H/S +/sht 2 S H H H H H H H/S H/S H/S Summe der Tiere (n): Abkürzungen: H: Hautprobennahme; H/S: Hautprobennahme und Sektion des gleichen Tieres; S: Sektion Zunächst wurden dann mit einem Lineal Rumpf- und Schwanzlängen erfaßt. Anschließend sind die Körpergewichte der einzelnen Mäuse und die Leberund Nierengewichte nach ihrer Entnahme ermittelt worden (Mettler PB 3000, Fa. Mettler Waagen GmbH, Giessen). Für zukünftig durchzuführende Genomanalysen erfolgte eine Asservation von Proben der Leber, der Milz und des Schwanzes durch Schockgefrieren in flüssigem Stickstoff. Im Anschluß daran wurden Hautproben entnommen sowie Proben der Haare zur Anfertigung von Trichogrammen gezupft.
61 Eigene Untersuchungen Hautproben Lichtmikroskopische Untersuchungen Für die lichtmikroskopischen Untersuchungen wurden an den verschiedenen Lebenstagen von den jeweils acht Tieren beider Genotypen Hautproben von Bauch, Rücken, Hand, Fuß, Schwanz, Nase, Auge und Ohr entnommen. Vom kranialen Bauch und Rücken der Tiere ist jeweils ein etwa 1 cm 2 großes Stück Haut abpräpariert und auf ein etwa 2 cm 2 großes Korkplättchen plan anliegend verbracht worden. Aufgrund der Haftung der Unterhaut an dem Korkplättchen kann dabei auf eine weitere mechanische Befestigung verzichtet werden. Nach der im folgenden beschriebenen Fixation wurde ein jeweils 1 mm breiter Streifen aus der Mitte geschnitten und eingebettet. Die Lagerung des Hautstreifens erfolgte bei der Einbettung auf der medianen Schnittfläche, so daß im histologischen Schnittpräparat Längsschnitte der Haarfollikel zu sehen sind. Die Pfoten (Hände und Füße) der rechten Körperseite wurden abgetrennt und durch einen Vertikal- und einen Horizontalschnitt zerteilt. Aus dem proximalen Drittel des Schwanzes wurde eine zirka 1 mm dicke Scheibe quer abgetrennt und zudem ein etwa 10 mm langes Stück des darauffolgenden Schwanzes in Längsrichtung zweigeteilt. Dann wurde die Nase durch einen Horizontalschnitt in der Mitte der Sinushaarleiste abgetrennt und in der Medianen geteilt. Anschließend erfolgte die Abpräparation eines jeweils 2 mm breiten Hautstreifens vom linken Auge und vom linken Ohreingang. Alle Proben kamen im Anschluß an die unten aufgeführte Fixierung in Formalin (siehe Anhang) zur Einbettung. Zusätzlich zu der bereits beschriebenen Hautprobennahme wurden an Tag 21 zudem mit einer Hautstanze (Durchmesser: 4 mm) Hautproben von Rücken und Bauch gewonnen und auf Korkplättchen aufgezogen. Nach der Fixierung und Einbettung der ausgestanzten Hautstücke wurden daraus 3 µm dicke Transversalschnitte angefertigt. Der Paraplastblock wurde dabei zugetrimmt, bis die Ebene der Haarfollikelinfundibula distal der Talgdrüsenausführungsgänge erreicht war. Nun erfolgte eine Färbung der auf Objekträger gezogenen Schnitte mit Hämalaun-Eosin (siehe Anhang).
62 62 Eigene Untersuchungen Die Schnittebene der Infundibula wurde dann lichtmikroskopisch untersucht. Die Anzahl der Haarfollikelinfundibula pro Fläche (mm 2 ) ist dann morphometrisch mit dem halbautomatischen Bildanalysesystem Leitz ASM 68 K (Fa. Ernst Leitz Wetzlar GmbH, Wetzlar) erfaßt worden. Unter Verwendung des SAS-Programms (Fa. SAS Institute Inc., Cary, NC, USA 1988) wurden die Ergebnisse statistisch ausgewertet. Alle Proben wurden für 24 Stunden in 10%igem, gepufferten Formalin (siehe Anhang) fixiert. Die Entwässerung und Einbettung der Präparate erfolgte mit Hilfe eines Gewebeeinbettungsautomaten (Hypercenter II, Fa. Shandon, Frankfurt), wobei die Proben in Paraplast Plus (Fa. Shandon, Frankfurt) eingebettet wurden. Hiervon wurden dann mit einem Schlittenmikrotom (Modell HN40, Fa. Reichert und Jung, Nußloch) etwa 3 µm dicke Paraplastschnitte angefertigt und auf mit Chromalaun-Gelatine beschichtete Objekträger aufgezogen. Es folgte eine Färbung mit Hämalaun-Eosin nach dem im Anhang dargestellten Protokoll und die lichtmikroskopische Auswertung der Schnitte. Die Proben der Extremitäten und des Schwanzes wurden nach der Fixierung und vor ihrer Einbettung in 5%iger Salpetersäure für 24 Stunden entkalkt und anschließend durch mehrmalige Spülungen in Wasser neutralisiert. Die Dokumentation der lichtmikrokopisch erhobenen Befunde erfolgte unter Verwendung eines Photomikroskops Typ Axiophot (Fa. Zeiss, Oberkochem) mit Schwarzweißfilmen (Agfaplan APX 25 Professional, Fa Agfa-Gaevert AG, Leverkusen).
63 Eigene Untersuchungen Elekronenmikroskopische Untersuchungen Für die transmissionselektronenmikroskopischen Untersuchungen sind Proben der Haut von Bauch und Rücken der sechs, neun und zwölf Tage alten Tiere genommen worden. Bei diesen Altersstufen konnten anagene Wachstumsstadien der Haarfollikel angenommen werden. Es wurde jeweils ein etwa 1 cm 2 großes Stück Haut abpräpariert (kaudal der in Kapitel beschriebenen Lokalisation) und auf ein etwa 2 cm 2 großes Korkplättchen verbracht. Es wurde in Abständen von einem Millimeter kammartig parallel eingeschnitten, so daß ein Verbindungsrand bestehen blieb und 24 Stunden lang in 2,5%igem Glutaraldehyd in 0,1 M Cacodylatpuffer (siehe Anhang) immersionsfixiert. Es folgten weitere Einschnitte im Abstand von einem Millimeter im rechten Winkel zu den bereits vorgenommenen, so daß Gewebestücke von 1 mm 2 entstanden. Diese Stücke wurden mehrfach in 0,1M Cacodylatpuffer gewaschen und für 2 Stunden in 1%iger Osmiumtetroxydlösung (siehe Anhang) nachfixiert. Nach weiteren Waschschritten (je 30 Minuten) in Cacodylatpuffer wurden die Blöcke in einer aufsteigenden Alkoholreihe dehydriert (je 30 Minuten). Es folgte eine halbstündige Infiltration in Propylenoxid (Fa. Serva, Heidelberg, Nr.: 33715). Weiterhin wurden die Proben mit einem Propylenoxid-Epon-812-Gemisch (1:1) infiltriert, um schließlich in reinem Epon-812 (Fa. Serva, Heidelberg, Nr.: 21045) eingebettet zu werden. Die Polymerisation erfolgte in einem Polymerisationsschrank bei 45 C für einen Tag und bei 60 C für drei Tage. Die Kunststoffblöckchen wurden mit einem Pyramitom (TM 60, Fa. Reichert und Jung, Nußloch) auf die Probengröße zugetrimmt. Anschließend wurden an einem Ultramikrotom (Ultramikrotom OM U3, Fa. Reichert und Jung, Nußloch) 0,5-1,0 µm dicke Schnitte angefertigt und mit 0,25%igem Toluidinblau bei 80 C flotierend gefärbt. Zur Vororientierung für die transmissionselektronenmikroskopischen Untersuchungen wurden die Semidünnschnitte lichtmikroskopisch beurteilt. Danach wurden die Schnittlokalisationen für die Zielpräparation der Ultradünnschnitte herangezogen.
64 64 Eigene Untersuchungen Die Herstellung der etwa 70 nm dicken Ultradünnschnitte erfolgte mit einem Ultramikrotom (Ultracut E, Fa. Reichert und Jung, Nußloch). Sie wurden auf Kupfernetze (Typ G 200 mesh, Fa. Science Service, München) verbracht und mit einem Kontrastierungsgerät (Ultrostainer, Fa. LKB Schweden) zunächst für 30 Minuten bei 40 C in einer gesättigten Uranylacetatlösung (Ultrostain I) kontrastiert. Anschließend wurde eine weitere Kontrastierung für 75 Sekunden in Bleicitratlösung (Ultrostain II) bei 20 C vorgenommen. Die elektronenmikroskopische Auswertung der Ultradünnschnitte fand an einem Transmisionselektronenmikroskop (EM 10C, Fa. Carl Zeiss, Oberkochem) bei einer Spannung von 60 kv statt. Die Befunde wurden mit Planfilmmaterial (8,3 x 10,2; Fa. Guilleminot, Paris) fotographisch dokumentiert Haarproben Trichogramme Zur Anfertigung von Trichogrammen wurden Haare von Tieren der Altersstufen 21, 42 und 182 Tagen entnommen. Während dieser Lebensaltersstufen ist bei den normal behaarten Tieren aufgrund ihres Haarzyklus wegen der vorwiegenden Ausbildung von ausgewachsenen Telogenhaaren mit einer einheitlichen Größe zu rechnen. Die Haarproben wurden von der Kruppe der Tiere mit einer Pinzette ausgezupft, in 80%igem Alkohol gewaschen, auf einen Objektträger verbracht und mit Corbitbalsam eingedeckt. Jeweils 100 Haare der +/sht Tiere und 50 Haare der minderbehaarten Mäuse (sht/sht) wurden vermessen. Es erfolgte eine Erfassung der Längen und maximalen Breiten der einzelnen Haare mit dem halbautomatischen Bildanalysesystem Leitz ASM 68 K (Fa. Ernst Leitz Wetzlar GmbH, Wetzlar) unter Berücksichtigung des Alters und des Geschlechts. Aufgrund der oftmals vorhandenen rinnenartigen Einziehung der Haare besitzen sie keinen runden Querschnitt. Bei der Untersuchung der Maße der Haare wurde daher auf die Verwendung des Begriffs Durchmesser verzichtet. Stattdessen wird der Begriff Breite verwendet. Außer der Erfassung der Maße der Haare wurde die Verteilung der vier Haarsubtypen des Rumpfes bei den verschiedenen Altersstufen und den beiden Geschlechtern ermittelt. Die Ergebnisse sind unter Verwendung des SAS- Programms (Fa. SAS Institute Inc., Cary, NC, USA 1988) statistisch ausgewertet worden.
65 Eigene Untersuchungen Rasterelektonenmikroskopische Untersuchungen Von den Tieren der Altersstufen 21 und 182 Tagen wurden einzelne telogene Haare von der Kruppe ausgezupft und in 80%igem Alkohol gewaschen. Anschließend erfolgte eine Trocknung im Critical-Point-Drying-Apparatus (E3000, Fa. Polaron, London). Die getrockneten Haare wurden auf einen Aluminiumträger aufgeklebt (Plano Leittabs, Fa. Plannet GmbH, Wetzlar) und mit Goldpartikeln (15 nm Durchmesser) gesputtert (SCD 040, Fa. Balzers Union, Wiesbaden). Repäsentative Exemplare der Haarsubtypen der Mäuse wurden ausgewählt und mit Hilfe des Rasterelektronenmikroskops (Digital Scanning Microscope DAM 940, Fa. Zeiss, Oberkochem) betrachtet. Die Oberfläche der Haare und ihr Kutikulamuster wurden auf abnorme Veränderungen untersucht und fotographisch dokumentiert (Ilford FP4 Plus 125, Fa. Ilford, Cheshire, England) worden Organproben Bei den vollständigen Sektion der Tiere an den Lebenstagen 0, 21, 42 und 182 sind alle inneren Organe makroskopisch untersucht worden. Die Gewichte der Lebern und der rechten Nieren aller sezierter Tiere wurden erfaßt (Mettler PB 3000, Fa. Mettler Waagen GmbH, Giessen). Zusätzlich zu den Proben der Haut sind Teile der folgenden Organe bei der Sektion entnommen worden: Herz: Lunge: Leber: Milz: Niere: Magen: Pankreas: Jejunum: Kolon: Skelettmuskel: Thymus: Gesamtquerschnitt durch beide Ventrikel und Vorhöfe Gesamtquerschnitt durch den linken Lobus Gesamtquerschnitt Gesamtquerschnitt Gesamtquerschnitt (einschließlich Nebenniere) Gesamtquerschnitt Gesamtquerschnitt Querschnitt einer Darmschlinge Querschnitt einer Darmschlinge Querschnitt (M. quadriceps) Gesamtquerschnitt
66 66 Eigene Untersuchungen Schilddrüse: Lymphknoten: Hoden: Ovar: Uterus: Mamma: Querschnitt durch den Kehlkopf mit lateralen Drüsenanteilen Gesamtquerschnitt (Ln. cervicalis superficialis) Gesamtquerschnitt einschließlich Nebenhoden Gesamtquerschnitt eines Ovars Längsschnitt eines Uterushornes Querschnitt eines kaudalen Mammakomplexes Für die lichtmikroskopische Untersuchung der Organproben wurde die Fixierung, Einbettung und Färbung analog zu der Vorgehensweise bei den lichtmikroskopischen Untersuchungen der Hautproben vorgenommen (siehe Kapitel ). Desweiteren wurden nach Entkalkung der fixierten Gewebe in 5%iger Salpetersäure für 24 Stunden ebensolche Schnitte der Tibia, des Brustbeins und des Schädels (mit Gehirn und Hypophyse) angefertigt. An Lebenstag null, dem Tag ihrer Geburt, wurden die Tiere aufgrund der geringen Größe und der weichen Konsistenz der Gewebe abweichend von dieser Vorgehensweise behandelt. Zuerst erfolgte die normale Hautprobenentnahme von Rücken und Bauch. Dann sind die Extremitätenenden, der Schwanz und der Kopf abgetrennt worden. Nach entsprechender Fixierung und Einbettung wurden nun von Rumpf und Kopf 3 µm dicke Stufenschnitte zur Erfassung aller inneren Organe angefertigt und mit Hämalaun-Eosin gefärbt. Eine Entnahme und Wägung der Leber und Niere erfolgte bei diesen Tieren aufgrund der geringen Größe nicht.
67 Eigene Untersuchungen Voruntersuchungen an Tieren der Ursprunglinie Auch an den Tieren der Ursprungslinie HDA32/Thb wurden Untersuchungen vorgenommen um zu zeigen, inwieweit sich die Tiere des phänotypisch unveränderten +/sht Genotyps schon von den Tieren der Ursprungslinie unterscheiden. Diese Untersuchungen fanden analog zu denen der sht/sht bzw. +/sht Tiere statt, wurden allerdings in reduziertem Umfang vorgenommen: Zu den Zeitpunkten der ausschließlichen Hautprobennahmen (Lebenstag drei bis 18) wurde jeweils nur ein männliches und ein weibliches Tier untersucht. Ihre Körpermaße wurden erfaßt und ihre Organe makroskopisch und lichtmikroskopisch untersucht. Die Gewichte der Leber und einer Niere wurden ebenfalls ermittelt. Proben der Haut von Rücken und Bauch wurden ebenso wie ausgezupfte Haare des Rückens lichtmikroskopisch untersucht Auswertung der Voruntersuchungen Äußerlich können die Tiere der Ursprungslinie HDA32/Thb in keiner Weise von den heterozygoten, normal behaarten Anlageträgern (+/sht) unterschieden werden. Bei der makroskopischen und lichtmikroskopischen Untersuchung ihrer inneren Organe im Rahmen der Sektionen fanden sich keinerlei pathomorphologische Veränderungen. Alle Organe waren normal ausgebildet und zeigten einen makroskopisch und mikroskopisch normalen Aufbau im Vergleich zu den Tieren des +/sht Genotyps. Ebenso glichen die Haarfollikel und ihr Wachstumsrhythmus denen der +/sht Mäuse. Auch ihre Haare ließen makroskopisch und lichtmikroskopisch keine Unterschiede zu den normal behaarten Mäusen erkennen. Es sind alle vier Haarsubtypen mit vergleichbaren Maßen zu denen des +/sht Genotyps nachweisbar. Ein Vergleich der erfaßten Daten der +/sht Tiere mit denen der Ursprungslinie HDA 32 in den Merkmalen Rumpf- und Schwanzlänge, Körper-, Leber- und Nierengewichte wurde durch eine mit linearen Modellen gerechnete Varianzanalyse (SAS, Procedure GLM) mit den fixen Effekten Alter, Geschlecht und Genotyp vorgenommen. Dieser Vergleich ergab keine über alle Altersstufen einheitlichen, signifikanten Abweichungen (Signifikanzgrenze p < 0,05) zwischen den beiden Genotypen. Bei den älteren Tieren (> 21 Tage) konnte eine nur zum Teil signifikante Abweichung der Körper-, Leberund Nierengewichte festgestellt werden.
68 68 Eigene Untersuchungen Diese Abweichungen der Gewichte sind jedoch nicht konstant über die verschiedenen Altersstufen zu beobachten. So wiegen die Lebern der 21 Tage alten Tiere der Ursprungslinie HDA32/Thb beispielsweise hochsignifikant weniger, als die der heterozygoten Mäuse. An Tag 42 sind diese Organe jedoch wieder hochsignifikant schwerer, an Tag 182 wiegen sie wiederum nicht signifikant weniger. Einschränkend muß dabei auf die geringe Zahl der Beobachtungen bei den Tieren der Ursprungslinie hingewiesen werden. Auch unterschiedliche Wurfgrößen und damit verbundene wurfabhängige Effekte, aber auch andersartige maternale Effekte müssen als beeinflussende Faktoren berücksichtigt werden, da bei diesen Untersuchungen keine Vollgeschwister verglichen wurden. Optimale Kontrolltiere für den Vergleich mit den sht/sht Mäusen wären sicherlich die Tiere der Ursprungslinie HDA32/Thb. Da diese jedoch innerhalb eines einzigen Wurfes mit Nachkommen aller drei verschiedener Genotypen (+/+, +/sht, sht/sht) nicht zu identifizieren und von den +/sht Tieren zu unterscheiden sind, wurde hiervon Abstand genommen und der oben beschriebene Anpaarungsmodus gewählt. Eine Verwendung der Mäuse der Ursprungslinie als Kontrolltiere bietet sich erst an, wenn nach Identifizierung des veränderten Gens die Anwendung eines Gentestes möglich ist.
69 Eigene Untersuchungen Ergebnisse Befunde an den Organe Es sind, abgesehen von der andersartigen Behaarung, zu keinem Zeitpunkt der Untersuchungen weitere äußerliche Abnormitäten an den sht/sht Tieren feststellbar. Alle Extremitäten, einschließlich der Zehenendorgane und des Schwanzes, sind normal ausgebildet. Die Öffnung der Ohren und Augen findet zeitgleich mit den normal behaarten Wurfgeschwistern statt. Die körperliche Entwicklung erscheint ungestört und weder verzögert, noch beschleunigt. Im Verhalten sind bei den Tieren beider Genotypen keine Auffälligkeiten feststellbar. Physiologische Lautäußerungen können bei beiden untersuchten Genotypen wahrgenommen werden. Bei den durchgeführten Sektionen der sht/sht- und der +/sht Tiere der vier Altersstufen (0, 21, 42 und 182 Lebenstage) ließen sich folgende Befunde erheben: Bei allen sezierten Mäusen war in der Abdominalhöhle ein gut gefüllter Magen-Darm-Trakt und eine gefüllte Blase zu finden. Die Milzen der Tiere aus der jüngsten Altersgruppe (Lebenstag 21) zeigten zumeist eine gering- bis mittelgradig ausgeprägte Splenomegalie und Hyperämie. In der Thorakalhöhle fiel bei der Mehrzahl der Tiere eine geringgradig emphysematöse Veränderung der Lungen auf. Diese betraf vor allem die kranialen Lappen beider Lungen. Bei vier männlichen Tieren konnte ein hochgradiger Hämothorax festgestellt werden. Diese vier Tiere wurden im Alter von 21 (ein sht/sht Männchen und ein +/sht Männchen) und 42 Tagen (ein sht/sht Männchen und ein +/sht Männchen) seziert. Die im Thorakalbereich und an der Milz erhobenen Befunde können als tötungsbedingte Artefakte interpretiert werden. Durch die vorgenommene Streckung der Tiere kann es zu Zerreißungen der blutführenden Gefäße im Hals- und Brustbereich kommen. Dies führt in der Folge zu einer hochgradigen Blutung in die umliegenden Gewebe bzw. Hohlräume. Ein weibliches +/sht Tier zeigte im Alter von 42 Tagen einen mit geringen Mengen eines weißlich-trüben, mukösen Sekretes gefüllten Uterus. Beide Uterushörner dieser Maus waren dabei geringgradig vergrößert und hatten einen Durchmesser von drei bis vier Millimetern. Histologisch handelte es sich
70 70 Eigene Untersuchungen dabei um eine geringgradige, nichtzystische, papilliforme Hyperplasie des Endometriums. Die Ovarien zeigten dabei keine Veränderungen. Außer den beschriebenen Befunden wiesen die bei den Sektionen der Lebenstage 0, 21, 42 und 182 untersuchten inneren Organe der sht/sht Mäuse keine weiteren pathomorphologischen Veränderungen auf. Herz, Lunge, Leber, Milz, Niere mit Nebenniere, Magen, Pankreas, Dünndarm, Dickdarm, Skelettmuskel (M. quadriceps), Thymus, Schilddrüse, Lymphknoten (Ln. cervicalis superficialis), Mamma und Hoden mit Nebenhoden, resp. Ovar mit Uterus waren vollständig vorhanden und makroskopisch normal ausgebildet. Es waren keinerlei kongenitale Mißbildungen feststellbar. Auch die lichtmikroskopische Untersuchung der histologischen Schnitte dieser Organe ergab einen normalen histologisch-anatomischen Aufbau ohne Abweichungen zu den Befunden der normal behaarten Wurfgeschwister des +/sht Genotyps. Bei der Betrachtung der Sektionsbefunde muß auf die relativ geringe Zahl adulter sezierter Tiere hingewiesen werden. Insgesamt wurden lediglich 16 Mäuse in einem Alter von 42 oder mehr Lebenstagen beider Genotypen (sht/sht und +/sht) untersucht. Da keine kongenitalen Mißbildungen vorliegen, ist die Wahrscheinlichkeit des Auftretens anderer pathomorphologischer Veränderungen, die bei einer größeren Anzahl von untersuchten älteren Tieren zu erwarten wären, relativ gering.
71 Eigene Untersuchungen Statistische Auswertung der erfaßten Körpermaße Wegen der relativ geringen Anzahl der Beobachtungen pro Lebenstag wurden die Beobachtungswerte für die statistische Auswertung in Altersgruppen zusammengefaßt: Lebenstag null (der Tag der Geburt) bildet dabei alleine eine Gruppe, da hier aufgrund der durchgeführten Sektionstechnik ein Teil der ausgewerteten Daten nicht erhoben werden konnte. Da die Tiere als Ganzes eingebettet wurden, konnten Nieren- und Lebergewichte nicht ermittelt werden. Die Tiere der Altersstufen drei, sechs und neun bilden die zweite Untersuchungsgruppe, die juvenilen Mäuse im Alter von 12 bis 21 Tagen bilden die Dritte. Die beiden ältesten Tiergruppen (Sektionstag 42 und 182) stellen mit den voll adulten Mäusen die vierte Untersuchungsgruppe dar. Die Auswertung der erfaßten Daten erfolgte auch hier, wie schon bei den Tieren der Ursprungslinie beschrieben, durch eine mit linearen Modellen gerechnete Varianzanalyse (SAS, Procedure GLM) mit den Effekten Alter, Geschlecht und Genotyp (Signifikanzgrenze: p < 0.05). Es wurden sowohl die Rumpf- und Schwanzlängen, als auch die Körper-, Leber- und Nierengewichte auf Unterschiede innerhalb der Altersgruppen hinsichtlich des Geschlechts und des Genotyps und der Wechselwirkung dieser beiden Effekte untersucht. Alle erfaßten Daten der Körpermaße und Körpergewichte beider Genotypen sind in Abhängigkeit des Alters und des Geschlechtes der Tiere vollständig in Tabelle 3.1 dargestellt. Bei allen Genotypen ist der für Mäuse charakteristische Verlauf der Entwicklung des Körpergewichts nachzuvollziehen. Einer Phase rapiden Wachstums mit einer ungefähren Verdoppelung des Körpergewichtes innerhalb einer Woche (Lebenstag drei bis neun) folgt eine verlangsamte, aber stetige Gewichtszunahme. Signifikante Unterschiede sind bei den beiden ersten Altersgruppen nicht feststellbar. Die Tiere der verschiedenen Genotypen zeigen dagegen hinsichtlich ihres Körpergewichtes und der Körpermaße deutliche Unterschiede ab dem Lebenstag neun (Altersgruppen drei und vier). Dies wird in Abbildung 3.2 und Abbildung 3.3 verdeutlicht.
72 72 Eigene Untersuchungen Desweiteren sind in der Altersgruppe vier (42 bis 182 Tage) neben den im folgenden näher beschriebenen Abweichungen hinsichtlich der Körpermaße der beiden Genotypen auch geschlechtsabhängige Unterschiede innerhalb eines Genotyps zu verzeichnen. Die adulten weiblichen Tiere sowohl des sht/sht Genotyps, als auch des +/sht Genotyps sind deutlich leichter, als die jeweils gleichaltrigen Männchen. Diese geschlechtsabhängigen Gewichtsunterschiede lassen sich innerhalb der vierten Gruppe auch bei den Gewichten der Einzelorgane nachvollziehen. Die Lebergewichte (p = 0,59), aber vor allem die Nierengewichte (hochsignifikant, p = 0,002) liegen bei den Weibchen unter denen der entsprechenden Böcke. Hinsichtlich des Genotyps finden sich Unterschiede lediglich bei den Gewichten der Lebern der Tiere der dritten Altersgruppe (Tag 12 bis 21). Die minderbehaarten Tiere zeigen hier signifikant (p = 0,03) geringere Lebergewichte als ihre normal behaarten Wurfgeschwister. Die Organgewichte weisen ansonsten keine signifikanten Unterschiede zwischen den beiden Genotypen auf.
73 Eigene Untersuchungen 73 Abbildung 3.2: Entwicklung des Körpergewichts beider Genotypen Gewicht (g) sht/sht +/sht Grp. 1 Grp. 2 Grp. 3 Grp Lebensalter in Tagen (Einteilung der Altersgruppen: Gruppe 1 = Tag 0 / Grp. 2 = Tag 3 bis 9 / Grp. 3 = Tag 12 bis 21 / Grp. 4 = Tag 42 bis 182) Das Gesamtkörpergewicht zeigt bei den beiden verschiedenen Genotypen deutliche Unterschiede: die minderbehaarten Tiere (sht/sht) der dritten Altersgruppe (Tag 12 bis 21) wiegen hochsignifikant (p = 0,0001) weniger als die gleichaltrigen Mäuse des +/sht Genotyps. Das geringere Gewicht der minderbehaarten Tiere läßt sich bei der vierten Altersguppe ebenfalls signifikant nachweisen (p = 0,08). Ab dem Lebenstag neun ist die Tendenz zu erkennen, daß die minderbehaarten Tiere ein geringeres Körpergewicht aufweisen. Die Tiere bleiben ab diesem Zeitpunkt konstant leichter als die normal behaarten Wurfgeschwister des +/sht Genotyps.
74 74 Eigene Untersuchungen Abbildung 3.3: Entwicklung der Körpermaße beider Genotypen Längen (mm) Grp. 1 Grp. 2 Grp. 3 Grp. 4 Rumpflänge sht/sht Rumpflänge +/sht Schwanzlänge sht/sht Schwanzlänge +/sht Lebensalter in Tagen (Einteilung der Altersgruppen: Gruppe 1 = Tag 0 / Grp. 2 = Tag 3 bis 9 / Grp. 3 = Tag 12 bis 21 / Grp. 4 = Tag 42 bis 182) Die Tiere der beiden Genotypen zeigen ab der dritten Altersgruppe (Lebenstag 12 bis 21) hochsignifikante Unterschiede ihrer Rumpf- und Schwanzlängen: Die minderbehaarten sht/sht Genotypen besitzen einen deutlich kleineren Rumpf (p = 0,006) und einen kürzeren Schwanz (p = 0,004). Diese Unterschiede lassen sich signifikant (Rumpflänge; p = 0,02) bzw. schwach signifikant (Schwanzlänge; p = 0,05) auch bei den adulten Tieren der vierten Altersgruppe nachweisen. Die sht/sht Tiere bleiben also zeitlebens kleiner, als ihre normal behaarten Wurfgeschwister. Dies ist sowohl anhand der Rumpf- als auch anhand der Schwanzlängen nachvollziehbar.
75 Eigene Untersuchungen 75 Tabelle 3.1: Körpermaße und Organgewichte beider Genotypen in den verschiedenen Altersstufen und Geschlechtern Genotyp: sht/sht +/sht sht/sht +/sht Geschlecht: Parameter Alter Ø Ø Ø Ø Alter Ø Ø Ø Ø Rumpfl. (mm) 0 30,0 30,0 31,0 29, ,5 52,0 54,5 62,5 Schwanzl. (mm) 11,0 10,5 9,5 10,0 34,5 43,5 48,0 46,5 Körpergew. (g) 1,3 1,2 1,35 1,3 6,9 4,6 7,3 9 Lebergew. (mg) Nierengew.(mg) Rumpfl. (mm) 3 35,0 34,5 36,0 35, ,0 57,5 59,5 56,5 Schwanzl. (mm) 13,0 14,0 15,0 14,0 52,0 50,0 58,0 56,0 Körpergew. (g) 2,1 1,9 2,4 2,2 7,4 6,9 8,6 7,9 Lebergew. (mg) Nierengew.(mg) 13 14, Rumpfl. (mm) 6 45,0 43,5 42,5 41, ,5 57,5 66,0 65,5 Schwanzl. (mm) 24,0 23,5 16,0 23,0 58,5 58,0 61,5 63,5 Körpergew. (g) 4,6 4,3 3,5 4,5 8,9 9,4 10,8 10,7 Lebergew. (mg) Nierengew.(mg) Rumpfl. (mm) 9 44,5 48,0 50,0 48, ,5 77,0 83,5 74,0 Schwanzl. (mm) 28,5 32,5 31,0 28,0 68,0 56,0 79,5 83,5 Körpergew. (g) 5,4 5,6 6,5 5,9 20,0 15,7 21,3 18,3 Lebergew. (mg) Nierengew.(mg) Rumpfl. (mm) 12 52,0 52,0 54,5 49, ,5 88,0 95,5 97,0 Schwanzl. (mm) 33,5 38,5 38,0 39,5 88,5 89,0 93,0 92,0 Körpergew. (g) 5,9 6,5 6,9 7,6 28,0 26,7 32,3 27,9 Lebergew. (mg) Nierengew.(mg) Abkürzungen: : durchschnittliche Maße; Körpergew.: Körpergewicht; Lebergew.: Lebergewicht; Nierengew.: Nierengewicht; Rumpfl.: Rumpflänge; Schwanzl.: Schwanzlänge;
76 76 Eigene Untersuchungen Ophthalmologische Veränderungen Über den Beobachtungszeitraum von etwa eineinhalb Jahren wurden in dem Gesamtbestand der homo- und heterozygoten Mäusepopulation (sht/sht und +/sht) bei sechs adulten, minderbehaarten, männlichen Tieren Veränderungen im Bereich der Augen festgestellt. Diese minderbehaarten Mäuse gehörten nicht zu den Tieren des Sektionsplans, sondern dienten lediglich der weiteren Zucht zur Aufrechterhaltung der Population. Es handelte sich meist um einseitige, chronisch-rezidivierende Blepharitiden, die nicht durch eine intensive antibiotische und antiinflammatorische Therapie (Cortiphenol ) dauerhaft zu kontrollieren waren. Die histologische Untersuchung der Augen nach der Sektion eines Teils der hiervon betroffenen Tiere ergab eine fokale Ulzeration der Schleimhaut der Augenlider. Einzelne Haarfollikel waren hier hochgradig tordiert, so daß ihre Infundibula auf der Innenseite der Augenlider mündeten. Es lag in diesen Bereichen eine fokale, hochgradige perifollikuläre Infiltration der Dermis mit leukozytären Entzündungszellen vor. Vermutlich haben einzelne, verdreht wachsende Haarfollikel Haare bzw. Wimpern produziert, die zu Irritationen des Auges und Pruritus führten. Die daraus resultierende permanente Abwehrreaktion des Tieres hat vermutlich die chronische Entzündungsreaktion aufrecht erhalten. Da derartige Veränderungen nur bei einer sehr kleinen Anzahl von Tieren gesehen wurde, müssen sie als Nebenbefund interpretiert werden. Sie können nicht als charakteristisch für sht/sht Inzuchtlinie angesehen werden.
77 Eigene Untersuchungen Befunde am Haarkleid Behaarung des Rumpfes Die Rumpfbehaarung der unveränderten +/sht Tiere wird im Alter von drei bis vier Lebenstagen erstmals makroskopisch sichtbar. Die sht/sht Mäuse zeigen die ersten erkennbaren Haare erst fünf bis sechs Tage später, also im Lebensalter von acht bis neun Tagen. Bei den +/sht Wurfgeschwistern ist etwa ab dem 12. Lebenstag ein normales, dichtes Fell ausgebildet. Da es sich um eine Albino-Linie handelt, wird kein Pigment in die Haare eingelagert, weshalb die Haare weiß erscheinen. Bis zum Alter von drei Wochen hat das Fell seine endgültige Länge erreicht. Bei den sht/sht Mäusen wächst dagegen nur ein sehr spärliches, dünnes und gewelltes Haarkleid. Ab dem Alter von etwa vier Wochen ist keine Zunahme der Länge oder der Dichte ihres Haarkleides mehr feststellbar. Die Haare der sht/sht Tiere sind ebenfalls nicht pigmentiert, weshalb sie durchsichtig bis weiß erscheinen. Dieses Fell ist gleichmäßig über den gesamten Körper einschließlich des Kopfes und der Extremitäten ohne andersartige Areale ausgebildet. Ein einheitlicher Haarstrich ist nur eingeschränkt, vor allem an den Extremitäten, zu erkennen. Durch das dünne und kurze Haarkleid hindurch ist die gut vaskularisierte Haut der Tiere zu erkennen, woraus das rosafarbene Erscheinungsbild der minderbehaarten Mäuse resultiert (siehe Abbildung 3.4 und Abbildung 3.5).
78 78 Eigene Untersuchungen Abbildung 3.4: +/sht (oberes Tier) und sht/sht (unteres Tier) Wurfgeschwister, weiblich, Lebenstag neun. Es sind bereits in diesem Alter deutliche Unterschiede in der Behaarung des Körpers zu erkennen. Die Haarfollikel befinden sich noch in der Anagenphase; das Haarwachstum ist noch nicht abgeschlossen. Die sht/sht Tiere sind trotzdem anhand ihrer deutlich kürzeren Behaarung leicht von ihren normal behaarten +/sht Wurfgeschwistern zu unterscheiden. Makroaufnahme Abbildung 3.5: Adulte +/sht (oberes Tier) und sht/sht (unteres Tier) Wurfgeschwister, weiblich, Lebenstag 182. Die sht/sht Tiere erscheinen deutlich minderbehaart im Vergleich zu ihren gleichalten Wurfgeschwistern. Diese Hypotrichose ist gleichmäßig über den gesamten Körper ausgebildet. Es sind keine übermäßige Schuppung der Haut oder entzündliche Reaktionen makroskopisch erkennbar. Die Haarfollikel des +/sht Genotyps befinden sich in diesem Lebensalter überwiegend im Telogen, das Haarwachstum ist also abgeschlossen. Makroaufnahme
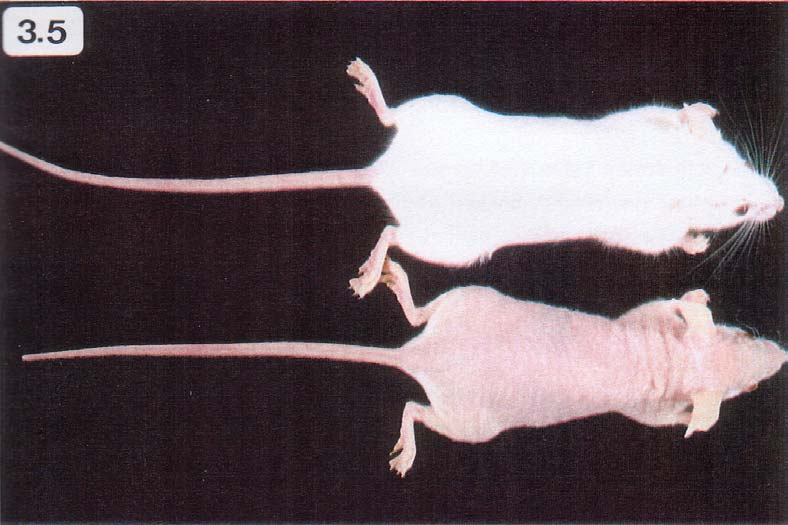
79 Eigene Untersuchungen 79
 von ihren normal behaarten Wurfgeschwistern zu unterscheiden.")
80 80 Eigene Untersuchungen Abbildung 3.6: Vibrissen bei sht/sht und +/sht Wurfgeschwistern, Lebenstag neun. Schon vom ersten Lebenstag an sind die sht/sht Tiere anhand ihrer feineren, kürzeren und gewellten Vibrissen (Schnurrhaare) von ihren normal behaarten Wurfgeschwistern zu unterscheiden. Die Vibrissen der sht/sht Tiere bleiben zeitlebens derartig verkürzt und stark gewellt. Makroaufnahme
81 Eigene Untersuchungen Morphologie der Haare Im Lichtmikroskop zeigt sich ein stark verändertes Erscheinungsbild der Haare der sht/sht Mäuse im Vergleich zu den Haaren der +/sht Tier. Das Haarkleid der +/sht Mäuse ist aus den im Schrifttumsteil beschriebenen vier Haarsubtypen des Rumpfes zusammengesetzt. Die Einzelhaare entsprechen in ihrer Morphologie und Größe den normalen Haaren der Maus. Bei den sht/sht Tieren ist nur ein einziger einheitlicher Haarsubtyp vorhanden. Dieser läßt zwar einen normalen Aufbau des Haares mit Spitze, Granne und Schaft erkennen, ist aber wesentlich verkleinert. Die geringere Größe der Haare wird ausführlich im folgenden Kapitel behandelt. Es kann aufgrund der Morphologie der sht/sht Haare im Lichtmikroskop keine Zuordnung zu den normalen Haarsubtypen der Maus vorgenommen werden. Während der Großteil der Haare des heterozygoten Genotyps (Leithaare und Ahlenhaare) einen geraden Verlauf zeigen, sind die Haare der homozygoten Tiere geringgradig unregelmäßig gewellt. Sie zeigen unterschiedlich starke Krümmungen mit einer ebenfalls sehr variablen Anzahl der ausgebildeten Krümmungen. Zumeist weisen sie lediglich eine einzige, langgezogene Biegung auf. Die Haare sind nicht vollständig eingerollt oder in sich verdreht. Ein Haarmark ist bei den sht/sht Haaren nur rudimentär ausgebildet. In unregelmäßigen Abständen sind kurze Intervalle von Luftkammern, die lediglich maximal einreihig ausgebildet sind, erkennbar. Im Durchlicht ist zumeist nur eine homogene Rindenschicht zu sehen (siehe Abbildung 3.7 ). Diese Art der Ausbildung des Haarmarkes wird als diskontinuierlich bezeichnet. Die Haare der sht/sht Mäuse besitzen das stark verkleinerte Erscheinungsbild eines normalen Haares. Sie gleichen über ihren gesamten Verlauf eher der Spitze oder dem Schaft eines normalen Zick-Zack-Haares. Die sht/sht Haare sind jedoch wesentlich kürzer und dünner als dieser normale Haarsubtyp. Zudem fehlen ihnen die für diesen Haarsubtyp charakteristischen Abknickungen und das kontinuierlich vorhandene, einreihige Haarmark.
82 82 Eigene Untersuchungen Abbildung 3.7: Auswahl von drei +/sht Haarsubtypen des Rumpfes und des einheitlichen sht/sht Haarsubtyps, Lebenstag 21. Der einheitlich ausgebildete, kürzere und schmalere Haarsubtyp der sht/sht Maus läßt sich aufgrund seiner Morphologie keinem der vier normalen Haarsubtypen des Rumpfes der Maus zuordnen (Ahlen- und Knickhaare der +/sht Tiere gleichen sich in ihrer Morphologie weitgehend). Das Haar besitzt zumeist keinen geraden Verlauf. Ein Haarmark ist nur unregelmäßig bzw. rudimentär ausgebildet. +/sht Haare L: Leithaar; A: Ahlenhaar bzw. Knickhaar; Z: Zick-Zack-Haar; sht/sht Haar: S Lichtmikroskopische Aufnahmen der Grannen ausgezupfter Telogenhaare, Eichstrich = 50 µm
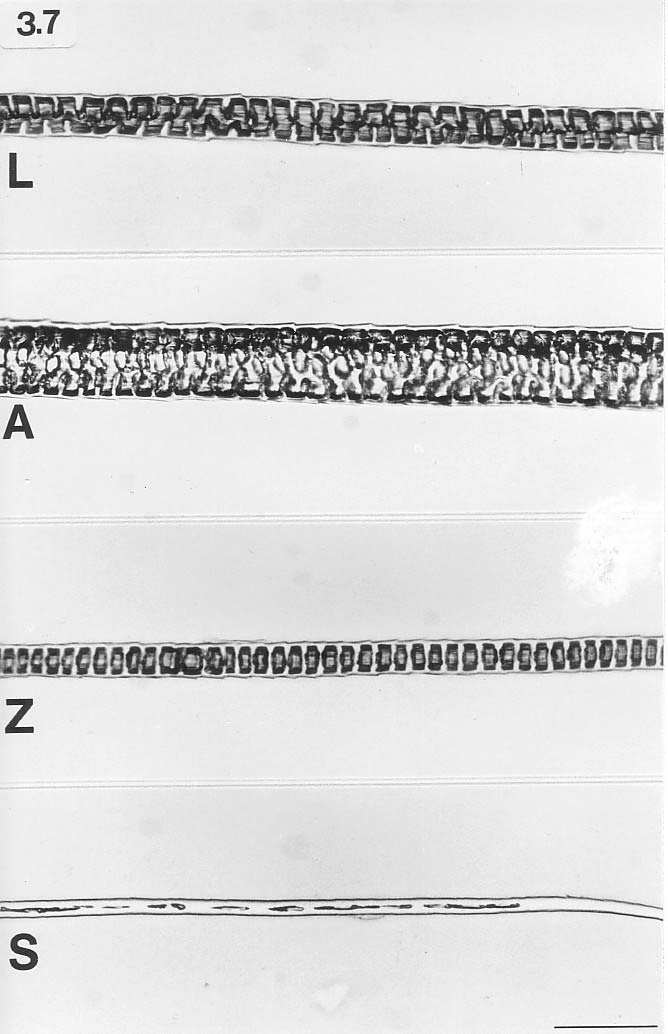
83 Eigene Untersuchungen 83
84 84 Eigene Untersuchungen Abbildung 3.8: Rasterelektronenmikroskopische Aufnahmen aller Haarabschnitte (Schaft, Granne und Spitze) des einheitlichen sht/sht Haarsubtyps und der normalen vier Haarsubtypen des Rumpfes der +/sht Maus. Ausgezupfe Telogenhaare, Lebenstag 21, Eichstrich = 10 µm Legende: Lokalisation: Haarspitze (distales Ende) Haargranne (breiteste Stelle des Haares) Haarschaft (proximales Ende) sht/sht sht/sht sht/sht +/sht Leithaar +/sht Leithaar +/sht Leithaar +/sht Ahlenhaar +/sht Ahlenhaar +/sht Ahlenhaar +/sht Knickhaar +/sht Knickhaar +/sht Knickhaar +/sht Zick-Zack-Haar +/sht Zick-Zack-Haar +/sht Zick-Zack-Haar

85 Eigene Untersuchungen 85
86 86 Eigene Untersuchungen Auch im Rasterelektronenmikroskop ist eine Zuordnung der vier +/sht Haarsubtypen des Rumpfes anhand ihrer Morphologie, der Größe und des Kutikulamusters möglich. Eine entsprechende Zuordnung der Haare der sht/sht Tiere zu den normalen Haarsubtypen kann jedoch nicht vorgenommen werden. Die Haare des sht/sht Genotyps zeigen eine einheitliche Morphologie mit einer wesentlich geringeren Länge und einem kleineren Durchmesser. Es ist auch im Rasterelektronenmikroskop ein normaler Aufbau des Haares mit einer geringen Zunahme des Durchmessers im Bereich der Granne zu erkennen. Die Haare sind in ihrem Verlauf unregelmäßig leicht gewellt. Sie erscheinen jedoch nicht eingerollt oder in sich verdreht. Brüche, Abknickungen oder begrenzte Auftreibungen sind im Verlauf der sht/sht Haare nicht vorzufinden. Die Kutikula ist vorhanden und ihre einzelnen Kutikulaschuppen umgeben das Haar vollständig, so daß sie dem kranzförmigen Typ zugerechnet werden müssen. Dieser Kutikulatyp ist über die gesamte Länge der Haare zu beobachten. Die Kutikulaschuppen sind überwiegend glatt und liegen dem Haar an. Sie weisen keine longitudinalen Fissuren auf (siehe Abbildung 3.8). Eine rinnenartige, parallel zur Längsachse verlaufende Vertiefung der sht/sht Haare, wie sie vor allem bei bei Ahlen- und Knickhaaren der normal behaarten Wurfgeschwister vorzufinden ist, ist im rasterelektronenmikroskopischen Bild nicht vorhanden. Die sht/sht Haare besitzen also einen überwiegend runden Querschnitt Maße der Haare Alle bei den +/sht Tieren gewonnen Meßwerte zeigen eine große Übereinstimmung mit den von DRY (1926) erhobenen Werten der Haare von Mäusen (siehe Tabelle 3.2). Auch die umfangreichen Untersuchungen von UHR (1984) bei der Maus (Mus musculus domesticus) ergeben vergleichbare Haarmaße.
87 Eigene Untersuchungen 87 Tabelle 3.2: Übersicht über die durchschnittlichen Maße der Haare des Rumpfes (Mittelwerte sowie Standardfehler s e der Varianzanalyse Genotyp Haarsubtyp Länge (mm) ± s e Breite (µm) ± s e Länge nach Dry (mm) Breite nach Dry (µm) sht/sht 1,17 0,03 8,83 0, /sht Zick-Zack-Haare 5,25 0,04 16,05 0,15 5,4 15 +/sht Knickhaare 6,34 0,12 27,57 0,69 6,5 24 +/sht Ahlenhaare 6,05 0,05 32,04 0,20 6,8 33 +/sht Leithaare 9,53 0,12 27,87 0,54 9,4 26 Abkürzungen: : männliche Tiere; : weibliche Tiere; : durchschnittliche Maße Die Haare der sht/sht Tiere sind insgesamt wesentlich kürzer als die der +/sht Mäuse. Ihre Länge beträgt etwa nur ein Fünftel der kürzesten Haare (Zick- Zack-Haare) ihrer normal behaarten Wurfgeschwister. Sie besitzen zirka ein Zehntel der Länge der längsten Haare (Leithaare) der +/sht Tiere. Auch ihre Breite ist deutlich geringer als die der normalen Haare der Maus. Sie beträgt durchschnittlich etwa die Hälfte der maximalen Breite der Zick-Zack-Haare und etwa nur ein Viertel der maximalen Breite der breitesten Haare, den Ahlenhaaren der +/sht Maus (siehe Abbildung 3.9 und Abbildung 3.10). Auch die geringen Schwankungen der Haarlängen und -breiten weisen, abgesehen von der einheitlichen Morphologie der sht/sht Haare, auf das Vorhandensein von nur einem einzigen Haarsubtyps hin (siehe Standardabweichungen der sht/sht Haare in Tabelle 3.3).
88 88 Eigene Untersuchungen Abbildung 3.9: Durchschnittliche Haarlängen der Genotypen sht/sht und +/sht 10 mm 9, ,25 6,34 6, ,17 0 sht/sht +/sht +/sht +/sht +/sht Zick-zack- Knickhaare Ahlenhaare Leithaare Haare (Standardfehler siehe Tabelle 3.2) Abbildung 3.10: Durchschnittliche maximale Haarbreiten der Genotypen sht/sht und +/sht µm ,57 32,04 27, ,83 16,05 0 sht/sht +/sht +/sht +/sht +/sht Zick-zack- Knickhaare Ahlenhaare Leithaare Haare (Standardfehler siehe Tabelle 3.2)
89 Eigene Untersuchungen 89 Tabelle 3.3: Haarmaße der Genotypen sht/sht und +/sht in Abhängigkeit von Alter und Geschlecht Länge (mm) Breite (µm) Genotyp/ Haarsubtyp/ Alter ± s ± s ± s ± s sht/sht 21 1,16 0,40 1,26 0,51 8,2 1,1 8,8 1,2 sht/sht 42 1,15 0,23 1,25 0,38 8,9 1,1 8,6 1,2 sht/sht 182 1,21 0,32 1,16 0,25 9,2 1,3 9,3 1,1 +/sht Zick-Zack-Haare 21 4,57 1,00 4,88 1,20 15,5 2,1 15,0 1,8 +/sht Zick-Zack-Haare 42 5,48 0,40 5,49 0,89 15,6 2,5 16,5 1,8 +/sht Zick-Zack-Haare 182 5,43 0,84 5,66 0,89 17,6 2,4 15,6 1,5 +/sht Knickhaare 21 4,99 1,2 5,92 0,90 25,8 7,7 25,3 8,7 +/sht Knickhaare 42 6,11 1,79 6,35 0,70 26,5 2,4 25,9 3,5 +/sht Knickhaare 182 7,33 0,51 7,09 0,75 38,9 0,0 30,5 8,5 +/sht Ahlenhaare 21 5,19 1,23 5,60 1,22 28,0 4,4 29,8 1,7 +/sht Ahlenhaare 42 5,89 0,90 6,14 0,81 35,2 7,5 31,1 5,7 +/sht Ahlenhaare 182 6,63 0,96 6,38 1,01 35,4 4,3 31,2 5,3 +/sht Leithaare 21 10,1 1,59 9,38 1,47 25,5 4,2 23,4 2,7 +/sht Leithaare 42 8,29 1,30 8,74 1,41 28,1 3,4 28,9 4,0 +/sht Leithaare ,9 1,14 10,3 1,10 37,2 3,5 27,0 1,4 Abkürzungen: : männliche Tiere; : weibliche Tiere; ± s: plus/minus Standardabweichung; : durchschnittliche Maße Zusammengefaßt zeigen die in Tabelle 3.3 dargestellten Ergebnisse, daß die +/sht Weibchen längere aber dünnere Haare aufweisen, als die männlichen Tiere dieses Genotyps. Auch die sht/sht Weibchen zeigen - allerdings bei gleicher Breite wie die Männchen - geringgradig längere Haare. Weiterhin ist festzustellen, daß die Einzelhaare aller Tiere (sht/sht und +/sht) mit zunehmendem Alter sowohl länger als auch dicker werden.
90 90 Eigene Untersuchungen Verteilung der Haarsubtypen des Rumpfes Aufgrund der einheitlichen Morphologie und der Maße der Haare des sht/sht Genotyps muß von der Ausbildung nur eines einheitlichen Haarsubtyps bei den minderbehaarten Mäusen ausgegangen werden. Dieser Haarsubtyp ist also bei der sht/sht Maus zu 100 Prozent ausgebildet. Er gleicht keinem der bei der Maus bisher bekannten Haarsubtypen. Zur Bestimmung des Verteilungsmusters der vier verschiedenen Haarsubtypen der Rumpfbehaarung der Maus wurden bei den normal behaarten +/sht Tieren 100 Haare pro Tier untersucht. Diese Untersuchung der Haarsubtypen der normal behaarten +/sht Tiere im Vergleich beider Geschlechter ergab keine signifikanten Unterschiede. Bei isolierter Betrachtung der Altersstufen konnte ein vermehrtes Vorkommen der Leithaare bei den 21-Tage-alten Tieren im Vergleich zu den 182-Tage-alten Mäusen festgestellt werden. Dagegen war ein geringerer Anteil von Ahlenhaaren bei den jungen Tieren (Lebenstag 21) vorzufinden. Die Anteile der Knick- und der Leithaare lassen über den Beobachtungszeitraum keine eindeutige Tendenz in Richtung Zu- oder Abnahme erkennen, wie aus Tabelle 3.4 zu entnehmen ist. Tabelle 3.4: Altersabhängige prozentuale Verteilung der Haarsubtypen des sht/sht und des +/sht Genotyps Genotyp: sht/sht +/sht +/sht +/sht +/sht Haarsubtyp: Zick-Zack- Knickhaare Ahlenhaar Leithaare (%) Haare (%) (%) e (%) (%) Lebenstag ,7 3,5 28,5 5,3 Lebenstag ,7 5,0 37,8 5,5 Lebenstag ,5 3,5 43,0 3,0 Ø ,0 4,0 36,4 4,6
91 Eigene Untersuchungen Vibrissen Schon zum Zeitpunkt der Geburt sind die homozygoten Anlageträger (sht/sht) von den heterozygoten +/sht Mäusen phänotypisch anhand der Vibrissen zu unterscheiden. Die Schnurrhaare sind bei den Mäusen die ersten makroskopisch erkennbaren Haare. Sie sind bei den nicht pigmentierten Mäusen schon am Tag der Geburt als kräftige, weiße, gerade Haare an der Schnauze sichtbar. Sie besitzen schon am ersten Lebenstag eine Länge von bis zu sechs Millimetern. Bei den sht/sht Tieren sind jedoch nur sehr feine, gelockte Vibrissen ausgebildet, die am Tag der Geburt makroskopisch nur schwer zu erkennen sind. Die Vibrissen erreichen bei den adulten +/sht Tieren eine maximale Länge von 30 Millimetern. Die dünnen und gewellten Schnurrhaare der adulten sht/sht Tiere errreichen eine maximale Länge von 15 Millimetern, wenn sie gestreckt werden. Sie besitzen desweiteren nur die Hälfte des Durchmessers der normalen Vibrissen (siehe Abbildung 3.6).
92 92 Eigene Untersuchungen Schwanzbehaarung Makroskopisch ist kein deutlicher Unterschied in der Behaarung des Schwanzes der beiden Genotypen festzustellen. Auch die lichtmikroskopische Beurteilung der Anordnung der Haarfollikel in der Schwanzhaut zeigt keine wesentlichen Abweichungen. Bei beiden Genotypen ist eine regelmäßige Anordnung der Haarfollikel in Dreiergruppen quer zur Längsachse des Schwanzes und ein daraus resultierendes schuppenartiges Erscheinungsbild der Schwanzhaut zu erkennen. Die Haarfollikel des Schwanzes der sht/sht Tiere weisen prinzipiell ähnliche Veränderungen wie die Haarfollikel des Rumpfes auf. Diese liegen jedoch in wesentlich geringerer Ausprägung vor. Die pathomorphologischen Veränderungen der Wurzelscheiden sind in ihrer Ausprägung und Häufigkeit maximal als geringgradig zu bezeichnen, entsprechen in ihrer Art aber denen der Haarfollikel des Rumpfes (siehe Kapitel ). Die Haarfollikel des Schwanzes und die von ihnen gebildeten Haare der sht/sht Tiere sind in ihrer Größe mit denen der normal behaarten Wurfgeschwister vergleichbar. Die Haare weisen im Gegensatz zu den Haaren des Rumpfes zumeist ein Haarmark auf. Sie zeigen zudem einen überwiegend geraden Verlauf.
93 Eigene Untersuchungen Befunde an der Haut und den Haarfollikeln Veränderungen der Haut des sht/sht Genotyps Die Haut der minderbehaarten sht/sht Mäuse ist durch das nur spärlich ausgebildete Fell hindurch gut sichtbar. Sie erscheint makroskopisch mattdunkelrosafarben und glatt. Es ist keine übermäßige Schuppenbildung erkennbar. Die Haut liegt dem Körper glatt an, ist verschieblich und zeigt keine vermehrte Faltenbildung. Ulzerationen oder Krusten sind an der Hautoberfläche makroskopisch nicht zu erkennen (siehe Abbildung 3.4 und Abbildung 3.5). Die Haut der sht/sht Tiere weist somit makroskopisch keine Veränderungen im Vergleich zu der Haut der +/sht Mäuse auf. Die lichtmikroskopische Untersuchung der Haut und der Haarfollikel läßt jedoch, ebenso wie die Beurteilung der Haare, deutliche Unterschiede zwischen den normalbehaarten und den minderbehaarten Mäusen erkennen. Die Befunde der aus verschiedenen Lokalisationen entnommenen Hautproben sind vergleichbar. An den Hautproben der Ohren und Augen wurden keine speziellen Veränderungen festgestellt. Abgesehen von der im folgenden beschriebenen geringeren Behaarung unterscheiden sich die Hautproben von Rücken und Bauch nicht. Die Dicke der gesamten Haut, das heißt der Epidermis, der Dermis und des Fettgewebes (Pannus) bis zum Hautmuskel, erscheint im histologischen Schnitt über den untersuchten Zeitraum bei beiden Genotypen vergleichbar. Die im Verhältnis zu anderen Säugern sehr dünne Epidermis der Maus weist mit Stratum basale bis Stratum corneum lediglich bis zu maximal vier Zell- Lagen auf. Die Epidermis der sht/sht Tiere läßt jedoch bis zu fünf Zell-Lagen erkennen und ist somit geringgradig verdickt. Diese geringgradige Akanthose ist bereits ab dem neunten Lebenstag bei allen untersuchten sht/sht Tieren zu sehen und bleibt über den gesamten Untersuchungszeitraum bestehen.
94 94 Eigene Untersuchungen Auf der Oberfläche der Epidermis der minderbehaarten Mäuse sind histologisch vermehrte Mengen verhornter Zellen, die keine Kerne aufweisen, vorzufinden. Der Keratinisierungprozeß der Epidermis ist jedoch als normal zu bezeichnen. Alle sht/sht Tiere zeigen lichtmikroskopisch ab dem 12. Lebenstag eine geringgradig ausgeprägte, orthokeratotische Hyperkeratose. Diese Hyperkeratose ist einheitlich bei allen minderbehaarten Tieren vorzufinden und bleibt ebenfalls über den gesamten Untersuchungszeitraum bestehen (siehe Abbildung 3.12). Schon ab dem sechsten Tag post partum sind in der Haut aller sht/sht Tiere vermehrt Entzündungszellen vorzufinden. Normalerweise finden sich in der Dermis und im Pannus normaler Mäuse gering- bis mittelgradige Mengen diffus verteilter Mastzellen. Neben diesem Entzündungszelltyp zeigt sich bei den sht/sht Tieren zusätzlich eine gemischtzellige Infiltration der Dermis, aber auch des Pannus. Diese liegt in individuell variabler Ausprägung vor. Die Entzündungszellen können in gering- bis hochgradiger Zahl vorhanden sein. Die Infiltrate bestehen aus neutrophilen Granulozyten, Lymphozyten und Histiozyten in veränderlichen Anteilen. Sie sind in erster Linie perifollikulär konzentriert. Innerhalb der verschiedenen Schichten der Follikel selber sind dabei keine Entzündungszellen nachzuweisen. Diejenigen Haarfollikel, in deren direkter Umgebung die Infiltrate vorliegen, weisen jedoch häufig hochgradige Defekte auf. Bei ihnen können im HE-gefärbten Schnitt unter Umständen nicht mehr alle üblicherweise vorhandenen Schichten eines normalen Haarfollikels (äußere Wurzelscheide, innere Wurzelscheide mit Henlescher und Huxleyscher Schicht) nachgewiesen werden. Es können entweder nur die äußere Wurzelscheide oder aber auch Schichten der inneren Wurzelscheide fokal begrenzt fehlen. Dieser Befund kann jedoch nicht einheitlich bei allen Haarfollikeln, die eine Entzündungszellansammlung in ihrer Umgebung zeigen, erhoben werden (siehe Abbildung 3.12 und Abbildung 3.17).
95 Eigene Untersuchungen 95 Vereinzelt liegen im Zentrum der Entzündungszellaggregate Teile von Haarschäften, ohne daß Strukturen eines sie umgebenden Haarfollikels erkennbar wären (siehe Abbildung 3.23). Derartige Aggregationen sind vorwiegend im Pannus, aber auch in der Dermis vorzufinden. Durch die Abwehrreaktion gegen diese normalerweise nicht frei in der Haut liegenden Strukturen kommt es zur Bildung von Fremdkörpergranulomen. Derartige Fremdkörpergranulome mit Teilen von Haaren in ihrem Zentrum werden auch als Trichogranulome bezeichnet. Diese Trichogranulome sind vorwiegend bei einzelnen Tieren des sht/sht Genotyps vorzufinden, die mehr als 21 Tage alt sind. Bei einzelnen minderbehaarten Tieren ist die Lokalisation der gemischtzelligen Entzündungszellinfiltrate im histologischen Schnitt nicht fokal auf die direkte Follikelumgebung begrenzt. Die Entzündungszellen liegen dann eher diffus in den interfollikulären Abschnitten der Dermis oder aber auch des pannikulären Fettgewebes verteilt. Es liegt somit bei allen Tieren des sht/sht Genotyps ab dem sechsten Tag post partum eine gering- bis mittelgradige, gemischtzellige, perifollikuläre oder diffuse Dermatitis vor. Bei adulten sht/sht Tieren können desweiteren vereinzelt Trichogranulome festgestellt werden. Zur Bestimmung der Dichte der Haarfollikel wurden Transversalschnitte der Haut angefertigt. Die Untersuchung dieser Präparate ergab dabei eine Anordnung der Infundibula der sht/sht Haarfollikel in der Haut in parallelen Reihen quer zur Längsrichtung des Tieres. Das entspricht der Ausrichtung der Haarfollikel ihrer normal behaarten +/sht Wurfgeschwister (siehe Abbildung 3.29 und Abbildung 3.27).
96 96 Eigene Untersuchungen Abbildung 3.11: Übersichtsaufnahme der Haarfollikel der +/sht Tiere, Lebenstag neun. Die anagenen Haarfollikel (Pfeil) liegen langgestreckt parallel in der Haut (D: Dermis; E: Epidermis). Ihre Bulbi (B) in der Subkutis (S) reichen einheitlich bis tief an den Hautmuskel (M) heran. Paraplasteinbettung, HE-Färbung, Eichstrich = 200 µm Abbildung 3.12: Übersichtsaufnahme der Haarfollikel der sht/sht Tiere, Lebenstag neun. Die geringgradig kleineren Haarfollikel (Pfeil) zeigen keine parallele Anordnung und einen stark gewundenen Verlauf, so daß nur Teilanschnitte von ihnen zu sehen sind. Die Haarfollikelbulbi reichen überwiegend nicht bis an den Hautmuskel heran. E: Epidermis; D: Dermis; M: Hautmuskel; S: Subkutis Paraplasteinbettung, HE-Färbung, Eichstrich = 200 µm
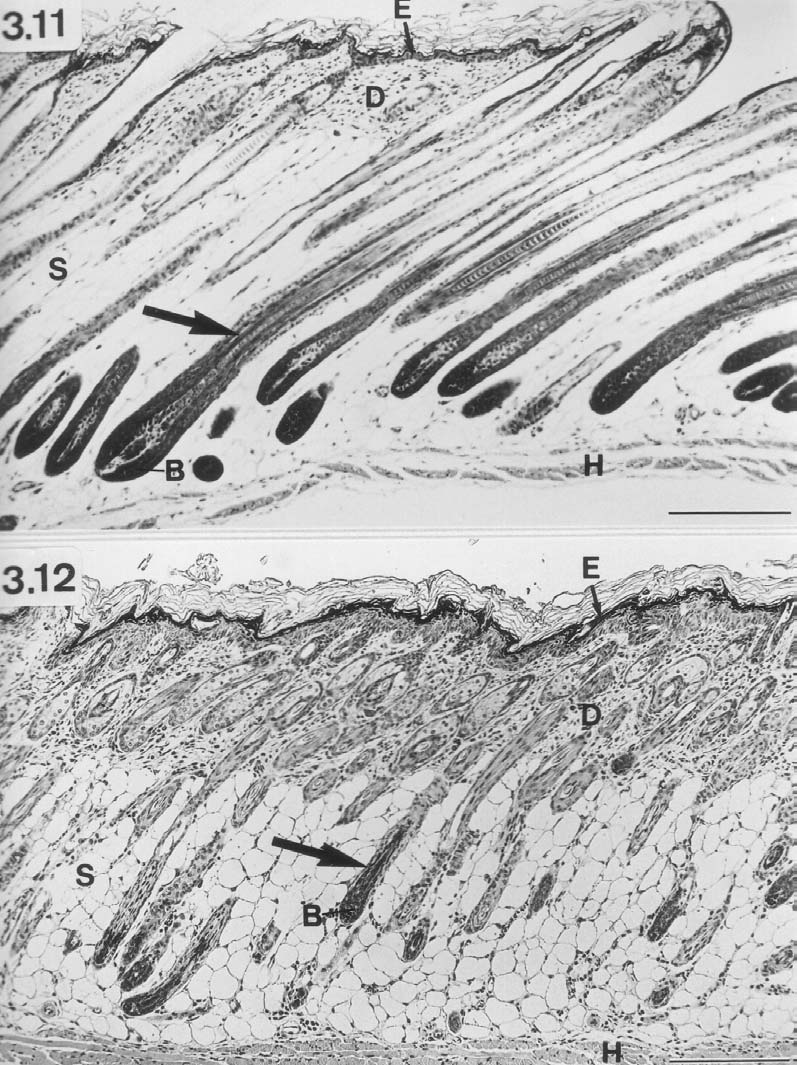
97 Eigene Untersuchungen 97
98 98 Eigene Untersuchungen Abbildung 3.13: Vollständiger anagener +/sht Haarfollikel, Lebenstag 12. Pfeil: Haar; B: Bulbus; D: Dermis; I: Infundibulum; S: Subkutis; Eponeinbettung, Toluidinblaufärbung, Eichstrich = 100 µm Abbildung 3.14: Anagener sht/sht Haarfollikel, Lebenstag 12. Das aus diesem Haarfollikel hervorgehende Haar besitzt eine deutlich geringere Größe und einen gewundenen Verlauf. Pfeil: Haar; B: Bulbus; D: Dermis; S: Subkutis; Eponeinbettung, Toluidinblaufärbung, Eichstrich = 200 µm
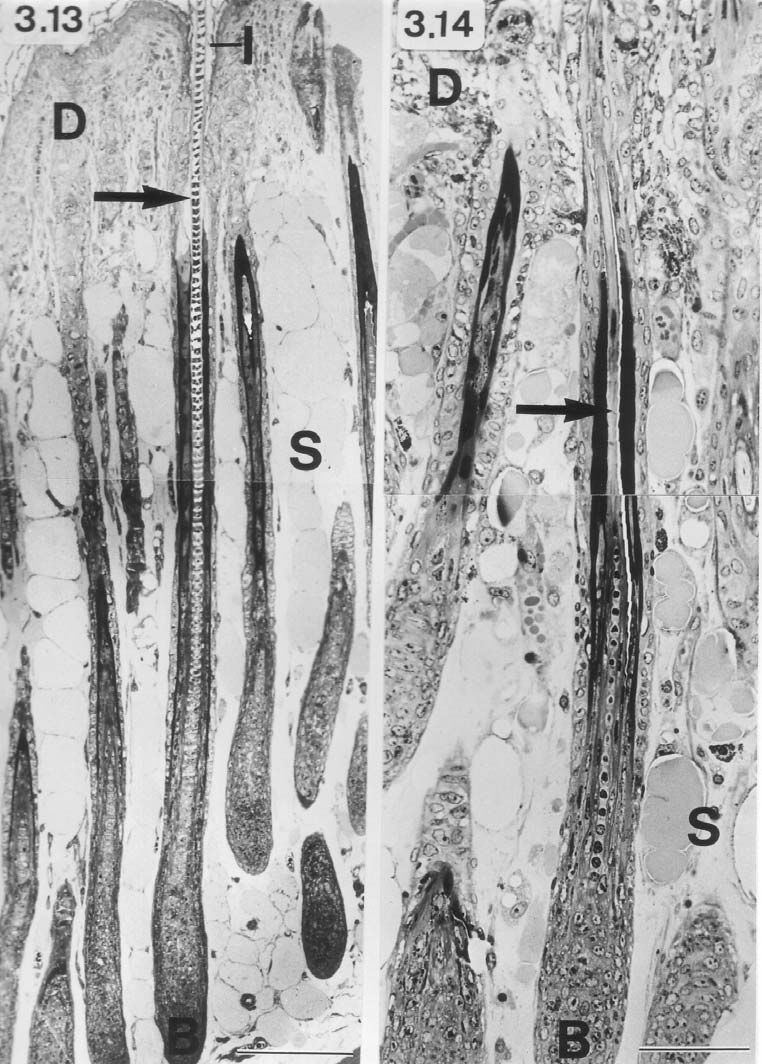
99 Eigene Untersuchungen 99
100 100 Eigene Untersuchungen Abbildung 3.15: Telogene Haarfollikel der +/sht Tiere, Lebenstag 21. Alle Haarfollikel der normal behaarten Mäuse befinden sich in diesem Lebensalter einheitlich im selben Zyklusstadium. Sie besitzen im Telogen eine wesentlich geringere Größe. Die innere Wurzelscheide ist nicht mehr ausgebildet. Ihr proximales Ende liegt etwa an der Grenze zwischen Dermis (D) und Subkutis (S). Im Haarfollikel liegen sogenannte Kolbenhaare (kleine Pfeile), die nun nicht mehr wachsen. Ihr proximales Ende ist geringgradig verdickt und marklos. Paraplasteinbettung, HE-Färbung, Eichstrich = 50 µm Abbildung 3.16: Haarfollikel der sht/sht Tiere, Lebenstag 21. Bei den sht/sht Mäusen befinden sich die Haarfollikel in diesem Lebensalter nicht einheitlich im selben Zyklusstadium. Es sind neben telogenen Follikeln mit kleinen Kolbenhaaren (kleiner Pfeil) auch katagene und sogar anagene Stadien (großer Pfeil) in der Subkutis (S; D: Dermis) zu sehen. Paraplasteinbettung, HE-Färbung, Eichstrich = 50 µm
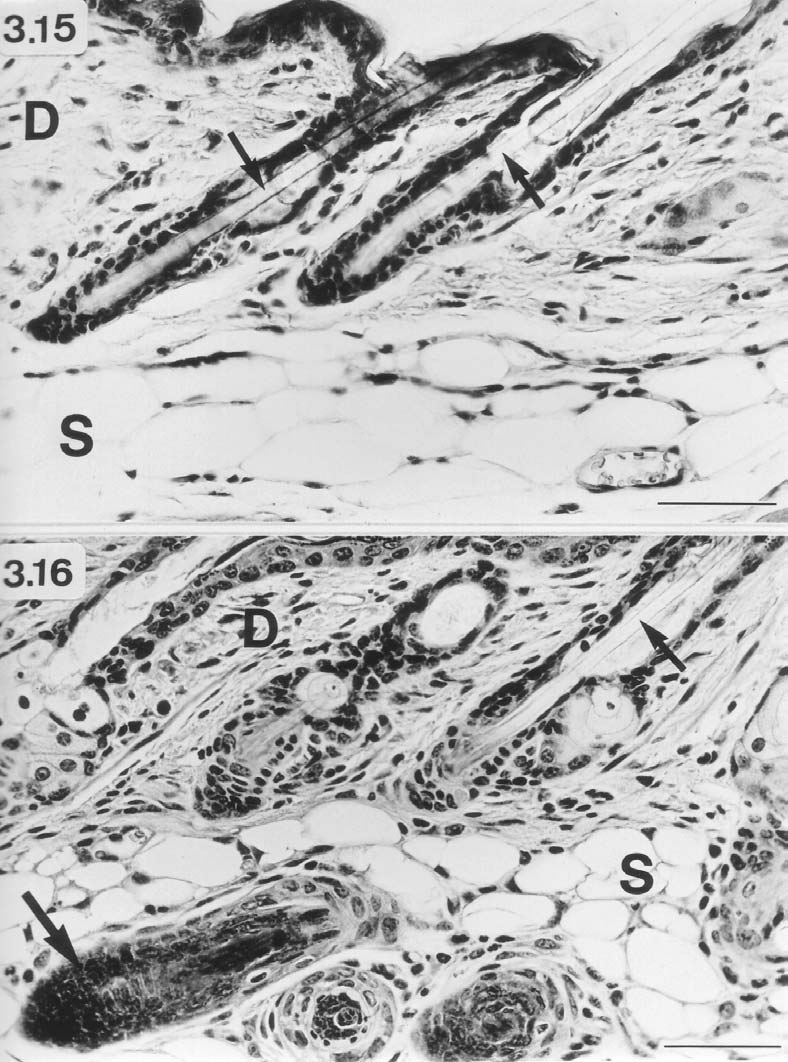
101 Eigene Untersuchungen 101
102 102 Eigene Untersuchungen Morphologie der Haarfollikel des sht/sht Genotyps Die Haarfollikel des minderbehaarten Genotyps produzieren Haare mit einer deutlich geringeren Größe als die Follikel der normal behaarten Wurfgeschwister. Diese Haare besitzen nur etwa ein Zehntel ihrer Länge und etwa die Hälfte bis ein Viertel des Durchmessers der normalen Haare. Obwohl die Haarfollikel des sht/sht Genotyps zwar geringgradig kleiner sind, weisen sie dennoch eine mit den kleineren Follikeln der normal behaarten Tiere vergleichbare Größe auf. Der mittlere Durchmesser der sht/sht-follikel entspricht in etwa dem der Haarfollikel, die bei den normal behaarten Tieren die Zick-Zack-Haare produzieren. Daraus resultiert ein Mißverhältnis zwischen der Größe der Haarfollikel der sht/sht Tiere zu der Größe der von ihnen gebildeten Haarfaser, denn das produzierte Haar ist wesentlich kleiner. Bei den sht/sht Tieren sind alle Anteile und Schichten eines normalen Haarfollikels angelegt. Sie weisen jedoch mehrere, zum Teil hochgradige morphologische Unterschiede zu den Follikeln ihrer normal ausgebildeten Wurfgeschwister auf: Die Haarfollikelinfundibula der minderbehaarten Tiere sind ab dem neunten Lebenstag mittel- bis hochgradig dilatiert. Bei normal behaarten Mäusen finden sich im Lumen der Haarfollikelöffnung zur epidermalen Oberfläche physiologischerweise nur geringe Mengen von Talgdrüsensekret und von Resten der verhornten inneren Wurzelscheide. Diese zerfällt und wird mit dem wachsenden Haar nach außen abgegeben. In den Infundibula des sht/sht Genotyps finden sich dagegen große Mengen keratinisierten Materials bzw. Talg. Die Lumina der Infundibula sind von der Öffnung des Talgdrüsenausführungsganges ausgehend nach distal stark erweitert. Es liegt eine mittel- bis hochgradige Infundibuladilatation und eine mittelgradige Follikelkeratose bei den Haarfollikeln der sht/sht Mäuse vor. Während die Follikel aller Altersstufen bzw. Haarzyklusstadien der +/sht Tiere wie zu erwarten einheitlich in streng paralleler Anordnung unter Bildung eines stumpfen Winkels mit der Epidermis in der Haut liegen, ist diese Ordnung bei den minderbehaarten Mäusen völlig aufgehoben.
103 Eigene Untersuchungen 103 Es ist lichtmikroskopisch keine einheitliche Ausrichtung der Haarfollikel erkennbar. Alle Follikel wachsen unabhängig voneinander in verschiedene Richtungen zur epidermalen Oberfläche. In einem histologischen Schnittpräparat sind sowohl Längsanschnitte, als auch Querschnitte von Haarfollikeln nebeneinander vorzufinden. Auch makroskopisch ist ein normaler, nach kaudal gerichteter Haarstrich nur undeutlich ausgebildet. Desweiteren ist die Lage der Haarfollikelbulbi bei den Tieren des sht/sht Genotyps sehr variabel. Entsprechend des normalerweise synchronen Haarzyklusstandes finden sich in einem histologischen Längsschnitt die Bulbi auf etwa demselben einheitlichen Niveau im Pannus. Bei den minderbehaarten Tieren finden sich die geringgradig kleineren Haarfollikelbulbi in einem histologischen Präparat in unterschiedlicher Tiefe im subkutanen Fettgewebe. Sie können unter Umständen innerhalb eines histologischen Präparates sowohl an der Grenze zwischen Dermis und Pannus als auch knapp über dem Hautmuskel angetroffen werden. Zudem zeigen die ungeordnet wachsenden Haarfollikel der sht/sht Tiere keinen geraden Verlauf. Ein normaler Haarfollikel zeigt ausgehend vom Bulbus, der im Anagen tief in der Dermis knapp über dem Hautmuskel liegt, einen geraden Verlauf in Richtung Epidermis (siehe Abbildung 3.11). Die Haarfollikel des sht/sht Gentyps sind dagegen mittel- bis hochgradig tordiert. Diese Torsionen sind völlig ungerichtet und ungeordnet. Die einzelnen Follikel sind unabhängig voneinander und verschieden stark abgebogen. Sie können einen Verlauf sowohl überwiegend parallel zur Längsachse des Tieres als auch quer zu dieser aufweisen. Es ist möglich, daß sich die Verlaufsrichtung eines einzelnen Follikels auf dem Weg zur epidermalen Oberfläche mehrmals ändert. Bei der Mehrzahl der Follikel schließen die Biegungen des Haarfollikels einen wesentlich größeren Winkel als 90 Grad ein. Es ist aufgrund dieser Torsionen nicht möglich, einen einzigen anagenen Follikel des sht/sht Genotyps in einem histologischen Längsschnitt vom Bulbus ausgehend bis zu seinem Infundibulum zu verfolgen (siehe Abbildung 3.12 und Abbildung 3.16).
104 104 Eigene Untersuchungen Abbildung 3.17: Haarfollikel des sht/sht Genotyps, Lebenstag 182. Die Infundibula der Follikel sind dilatiert (Pfeile) und mit verhorntem Detritus und Talgdrüsensekret angefüllt. Die Talgdrüsen (T) zeigen eine geringgradig ausgeprägte Hyperplasie. Eine milde, perifollikuläre Infiltration mit Entzündungszellen (EZ) ist in der Dermis (D) des sht/sht Genotyps häufig festzustellen. Die Haarfollikel sind aufgrund ihrer hochgradigen Torsionen nicht vollständig darstellbar. Es finden sich Längs- und Queranschnitte in einem Bild. In diesem Lebensalter befinden sich die sht/sht Haarfollikel ebenfalls nicht einheitlich im Telogen wie die des +/sht Genotyps. Paraplasteinbettung, HE-Färbung, Eichstrich = 100 µm Abbildung 3.18: Haarfollikel des sht/sht Genotyps, Lebenstag 182. Bei einigen Follikeln dieses Genotyps sind desweiteren unregelmäßige girlandenartige Einziehungen der äußeren Wurzelscheide (große Pfeile) festzustellen. Der Haarbalgmuskel (M) liegt normal ausgebildet in der Dermis (D). Paraplasteinbettung, HE-Färbung, Eichstrich = 100 µm

105 Eigene Untersuchungen 105
106 106 Eigene Untersuchungen Zusätzlich zu ihrem ungewöhnlichen Verlauf weisen die Haarfollikel der minderbehaarten Mäuse weitergehende morphologische Besonderheiten auf. Das von einem Follikel gebildete Haar liegt normalerweise im Zentrum des Follikelkanals und wird von den gleichmäßigen, konzentrischen Schichten der äußeren und inneren Wurzelscheide umgeben. Die feinen Haare der sht/sht Tiere liegen dagegen zumeist deutlich exzentrisch im Follilkellumen. Die Haare selbst behalten dabei ihren überwiegend runden Querschnitt. Die Schichten der inneren Wurzelscheide sind teilweise stark komprimiert. Auf der gegenüberliegenden Seite des Follikelquerschnitts kann die innere Wurzelscheide dagegen stark hyperplastisch erscheinen. Schichten, die normalerweise aus ein bis zwei Zell-Lagen bestehen, können dann bis zu vier Zellschichten aufweisen. Die Anzahl der Zell-Lagen kann aber auch der Norm entsprechen. In diesem Fall ist jedoch die Dicke der Zellschicht verändert. Die größte Variabilität bezüglich ihrer Anzahl an Zell- Lagen zeigt vor allem die Huxleysche Schicht der inneren Wurzelscheide. Die Kutikula der inneren Wurzelscheide und die Henlesche Schicht zeigen dagegen in vollständig keratinisiertem Zustand deutliche Abweichungen von ihrer normalen Dicke. Im Querschnitt erreichen diese Schichten eine bis zu dreifache Dicke im Vergleich zu den normalen Haarfollikeln. Die Anzahl ihrer Zell-Lagen ist dabei jedoch zumeist unverändert. Es liegt also eine fokal begrenzte Hypertrophie oder Hyperplasie der einzelnen Lagen der inneren Wurzelscheide vor. Auch die äußere Wurzelscheide variiert in der Anzahl ihrer Zelllagen, jedoch fallen die jeweiligen Unterschiede in den proximalen Ebenen nicht so deutlich aus, wie bei den Schichten der inneren Wurzelscheide. Distal der Talgdrüsenmündung kann die äußere Wurzelscheide auch deutlich hyperplastisch sein. Es sind in dieser Lokalisation im histologischen Schnitt bis zu drei Zelllagen nebeneinander im Querschnitt erkennbar. Im HE-gefärbten histologischen Schnitt erscheinen die äußere und inneren Wurzelscheiden bei einzelnen Follikeln teilweise aufgelöst bzw. abgebaut. Die transelektronenmikroskopischen Untersuchungen der Haarfollikel kann dies jedoch nicht bestätigen. Stellenweise erscheinen die Schichten des Follikels stark komprimiert und wesentlich dünner als normal.
107 Eigene Untersuchungen 107 Es sind jedoch auch in den sht/sht-follikeln alle Schichten ausgebildet. Zellkerne sind dabei in den stark komprimierten Bereichen der Zellschichten kaum vorzufinden. Ihre Lage ist vorwiegend auf die nicht komprimierten Bereiche der Zellen beschränkt. Bei den transelektronenmikroskopisch untersuchten Haarfollikeln konnten keine Entzündungszellinfiltrate gefunden werden. Bei einzelnen Follikeln des sht/sht Genotyps weist die äußere Wurzelscheide eine ungewöhnliche Morphologie auf. Erstmalig ist dies im Alter von neun Tagen post partum zu sehen. Entsprechende Befunde konnten zudem bei einzelnen Tieren der Altersstufen 12, 21 und 182 Tage im histologischen Schnitt vorgefunden werden. Die äußere Wurzelscheide zeigt dabei im Längsschnitt unregelmäßige Einziehungen, bei denen sich die Basalmembran der inneren Wurzelscheide annähert bzw. anlagert. Die normalerweise vorhandene einschichtige Zell-Lage der äußeren Wurzelscheide kann dabei vollständig verdrängt werden. An benachbarten Stellen ist sie dagegen stark hyperplastisch und auf mehrere Zell-Lagen verdickt. Die Wand des Haarfollikels erhält dadurch ein girlandenartiges Aussehen. Im histologischen Längsschnitt scheint die innere Wurzelscheide bei diesen Veränderungen nicht mitbetroffen zu sein. Die Einziehungen sind in einem histologischen Schnittpräparat zumeist nur bei einzelnen Follikeln vorzufinden. Direkt benachbarte Haarfollikel können nahezu unveränderte äußere Wurzelscheiden aufweisen (siehe Abbildung 3.18 und Abbildung 3.22). Vereinzelt sind im histologischen Schnittpräparat bei den minderbehaarten Mäusen Teile von Haarschäften völlig frei in der Dermis liegend vorzufinden. Strukturen eines sie umgebenden Haarfollikels sind dabei nicht erkennbar. Es scheint, als hätten in diesem Fall die Haarschäfte die Wurzelscheiden perforiert bzw. durch einen permanenten Reiz eine Follikulitis / Furunkulose hervorgerufen. Dies löst dann in der Folge die bereits oben beschriebenen Fremdkörperreaktionen der Haut mit Bildung von Trichogranulomen aus (siehe Abbildung 3.23).
108 108 Eigene Untersuchungen Abbildung 3.19: Hochgradige Torsion eines anagenen Haarfollikels (Pfeil) in der Haut einer sht/sht Maus im Längsschnitt, Lebenstag 21. Die Wurzelscheiden werden von dem bereits keratinisierten Haar einseitig verdrängt. Siehe auch Abbildung D: Dermis, S: Subkutis Paraplasteinbettung, HE-Färbung, Eichstrich = 50 µm Abbildung 3.20: Haarfollikel einer sht/sht Maus im Längsschnitt, Lebenstag 12. Innerhalb des vergleichsweise gerade verlaufenden Follikels zeigt das Haar einen deutlich gewundenen Verlauf und eine daraus resultierende exzentrische Lage. Die Wurzelscheiden weisen eine hochgradig variable Dicke auf. Es sind jedoch keine Einziehungen der äußeren Wurzelscheide, wie in Abbildung 3.22, vorhanden. Eponeinbettung, Toluidinblaufärbung, Eichstrich = 50 µm Abbildung 3.21 Haarfollikel einer sht/sht Maus im Querschnitt, Lebenstag 21. Das Haar liegt exzentrisch im Follikel (Pfeil) Der Haarfollikel ist hochgradig tordiert und seine Wurzelscheiden werden durch das Haar verdrängt, bzw. beinahe durchbrochen. Perifollikulär findet sich ein gemischtzelliges Entzündungszellinfiltrat in der Dermis (D). Siehe auch Abbildung Paraplasteinbettung, HE-Färbung, Eichstrich = 25 µm

109 Eigene Untersuchungen 109
110 110 Eigene Untersuchungen Abbildung 3.22: Deutliche, girlandenartige Einziehungen (Pfeile) der äußeren Wurzelscheide (äw) eines anagenen Haarfollikels einer sht/sht Maus, Lebenstag 12. In den Bereichen zwischen den Einziehungen ist die normalerweise einschichtige äußere Wurzelscheide mehrlagig ausgebildet. Das bereits vollständig keratinisierte Haar (H) liegt dabei zentral in der inneren Wurzelscheide (iw), die erst nach dem Haar verhornt. Die innere Wurzelscheide zeigt dagegen keine Einziehungen oder Unregelmäßigkeiten. D: Dermis; S: Subkutis, Eponeinbettung, Toluidinblaufärbung, Eichstrich = 50 µm Abbildung 3.23: Frei in der Dermis (D) liegender Teil eines Haarschaftes (Pfeil) mit reaktivem gemischtzelligem Entzündungszellinfiltrat in der Haut einer sht/sht Maus, Lebenstag 12. Umgebende follikuläre Strukturen sind dabei nicht sichtbar. Paraplasteinbettung, HE-Färbung, Eichstrich = 50 µm

111 Eigene Untersuchungen 111
112 112 Eigene Untersuchungen Die exzentrische Lage des Haares im Follikel bleibt nicht über die gesamte Länge des Haarfollikels konstant. Im Verlauf seines Wachstums wandert das Haar innerhalb des Follikels korkenzieherartig in mehreren Windungen innerhalb der äußeren Wurzelscheide nach distal. Die Anzahl der Windungen und das Ausmaß der Krümmung des Haares können dabei stark variieren. Die äußere Form des Haarfollikels bleibt trotz der inneren Veränderungen im Querschnitt rund bis rund-oval (siehe Abbildung 3.20). Hinsichtlich der ultrastrukturellen Ausstattung der Zellen in verschiedenen Schichten der Haarfollikel finden sich keine Unterschiede zwischen den Zellen der beiden Genotypen. Qualitative und quantitative Unterschiede lassen lediglich die Trichohyalingranula der inneren Wurzelscheide der Haarfollikel der sht/sht Tiere erkennen. Die Granula zeigen einen zum Teil deutlich vergrößerten Durchmesser. Ihr Durchmesser kann bis zu 4 Mikrometer betragen, während er normalerweise maximal 2 Mikrometer mißt. Zudem erscheint die Anzahl der Trichohyalingranula im Vergleich zu den Granula der inneren Wurzelscheide der normal behaarten Mäuse geringgradig verringert zu sein. Ein weiterer gravierender Unterschied zwischen den beiden Genotypen ist desweiteren eine andersartige Reihenfolge der Verhornung der einzelnen Schichten der Haarfollikel. Bei den normal behaarten Mäusen verhornen zuerst die Zellen der Henleschen Schicht und die Kutikula der inneren Wurzelscheide. Es folgen darauf die Rinde und das Mark des Haares. Schließlich verhornen die Huxleysche Schicht der inneren Wurzelscheide und die Kutikula des Haares. Bei den minderbehaarten sht/sht Tieren liegt dagegen eine deutlich abweichende Reihenfolge vor, denn Rinde, Mark (sofern vorhanden) und Kutikula des Haares zeigen bereits eine vollständige Verhornung, bevor die Zellen der inneren Wurzelscheide keratinisiert sind. Somit sind alle Anteile des Haares ausgehärtet, bevor der formgebende Kanal für das wachsende Haar gebildet ist. Da die Verhornung der haarbildenden Schichten des Haarfollikels zeitlich sehr viel früher einsetzt, kann in der Folge auch eine
113 Eigene Untersuchungen 113 veränderte Lokalisation des Keratinisierungsprozesses festgestellt werden. Bei den minderbehaarten Tieren sind Anschnitte von Haarfollikeln mit vollständig verhornten Haaren bereits tief in der Dermis vorzufinden. In der unveränderten Haut der Maus sind komplett verhornte Haare dagegen erst distal der sogenannten Keratinisierungszone des Haarfollikels in der Dermis, auf Höhe der Talgdrüsenausführungsgänge, zu sehen (siehe Abbildung 3.13 / Abbildung 3.14 und Abbildung 3.24 / Abbildung 3.25). Befinden sich die Haarfollikel der Mäuse im Telogenstadium, sind zwischen den beiden Genotypen hinsichtlich der Morphologie keine wesentlichen Unterschiede mehr festzustellen. Die Follikel sind in diesem Stadium bei beiden Mäusestämmen sehr kurz und gerade und beinhalten ein Kolbenhaar mit seinem charakteristisch verdickten proximalen Ende (siehe Abbildung 3.15). Eine Dilatation der Infundibula zeigen allerdings lediglich die Haarfollikel des sht/sht Genotyps, wie in Abbildung 3.17 dargestellt. Zudem können bei diesen Tieren in unmittelbarer Nachbarschaft der Telogenfollikel auch Follikel in anderen Haarzyklusstadien gefunden werden (siehe Abbildung 3.16). Zusammenfassend kann man also feststellen, daß die Haarfollikel der sht/sht Tiere im Vergleich zu den normal behaarten Mäusen eine Vielzahl unterschiedlicher und variabler Veränderungen im aufweisen. So zeigen sie eine Follikelkeratose mit dilatierten Infundibula bei einem völlig ungeordneten Wachstum und ungleichmäßig tief in der Dermis liegenden Haarfollikelbulbi. Sowohl die Follikel als auch die Haarschäfte sind dabei stark tordiert. Die Haare liegen deutlich exzentrisch in Windungen in den Follikeln, deren Wurzelscheiden Defekte oder Einziehungen aufweisen. Vereinzelt sind sogar freiliegende Stücke von Haarschäften in der Dermis oder Subkutis mit entsprechenden entzündlichen Reaktionen vorzufinden. Transmissionselektronenmikroskopisch konnte gezeigt werden, daß das Haar vor den Anteilen der inneren Wurzelscheide verhornt.
 zeigen Anzeichen der Verhornung vor den Anteilen des Haares (R / M).")
114 114 Eigene Untersuchungen Abbildung 3.24: Transmissionselektronenmikroskopische Aufnahme eines quergeschnittenen anagenen +/sht Haarfollikels, Lebenstag 12. Normaler konzentrischer Aufbau der Schichten des Haarfollikels. Die Zellen der inneren Wurzelscheide (He / Hu / K) zeigen Anzeichen der Verhornung vor den Anteilen des Haares (R / M). Abkürzungen von Abbildung 3.24 und Abbildung 3.25: äw: äußere Wurzelscheide; He: Henlesche Schicht der inneren Wurzel-scheide; Hu: Huxleysche Schicht der inneren Wurzelscheide; K: Kutikula der inneren Wurzelscheide; M: Haarmark; R: Haarrinde; T: Trichohyalingranula in der inneren Wurzelscheide und im Haarmark; Z: Zellkern einer Haarmarkzelle Eichstrich = 5 µm
 liegt deutlich exzentrisch im Follikel.")
115 Eigene Untersuchungen 115 Abbildung 3.25: Transmissionselektronenmikroskopische Aufnahme eines quergeschnittenen anagenen sht/sht Haarfollikels, Lebenstag 12. Das bereits vollständig keratinisierte Haar (R) liegt deutlich exzentrisch im Follikel. Die Zellen der inneren Wurzelscheide (He / Hu) sind noch nicht verhornt und unilateral stark hyperplastisch. Eichstrich = 5 µm
116 116 Eigene Untersuchungen Bestimmung des Zyklusstandes der Haarfollikel Die Hautproben der Tiere der Altersstufen von 0 bis 21 Lebenstagen und der adulten 42 und 182 Tage alten Mäuse beider Genotypen wurden vergleichend untersucht. Die Untersuchung der histologischen Schnitte des Rückens und des Bauches erstreckt sich also vorwiegend über den Zeitraum des kompletten ersten Haarzyklus der Tiere. Auch die zur Verfügung stehenden Hautproben der adulten Mäuse, die sich in späteren, nicht genau zu definierenden Haarzyklusstadien befinden, wurden darüberhinaus kontrolliert. An Rücken und Bauch lassen sich dabei prinzipiell die gleichen Befunde erheben. Am Tage der Geburt (Tag 0) befinden sich die Haarfollikel beider Genotypen noch in der Anbildung. Von Tag drei bis zum Tag 12 zeigen alle Haarfollikel des normal behaarten +/sht Genotyps anagene Wachstumsstadien (siehe Abbildung 3.26, breite schwarze Linie). An Lebenstag 15 ist bei diesen Tieren ein frühes Katagenstadium festzustellen, an Tag 18 befinden sich die Follikel entweder noch in einem späten katagenen Stadium oder bereits im Telogen, der Ruhephase des Haarfollikels. Im Alter von 21 Tagen sind dann nur noch ausschließlich telogene Follikel in der Haut der +/sht Mäuse vorzufinden. Die adulten Tiere der Altersstufen 42 und 182 Tage zeigen einheitlich ebenfalls telogene Ruhefollikel. Lediglich ein einzelnes Weibchen des +/sht Genotyps zeigt im Alter von 42 Tagen Haarfollikel, die sich im Anagen befinden. Die Tiere des minderbehaarten sht/sht Genotyps zeigen dagegen ein stark verändertes, sehr heterogenes Bild des Haarfollikelzyklus. Bis zum sechsten Lebenstag befinden sich alle Follikel im Anagen. Bereits an Tag neun sind jedoch schon bei allen Tieren katagene Stadien vorzufinden. Neben diesen sind innerhalb des selben Präparates weiterhin anagene Follikel festzustellen. Dieses Bild des gleichzeitigen Vorkommens anagener und katagener Stadien bleibt bis einschließlich des 15. Lebenstages unverändert. An Tag 18 sind neben verschiedenen Stadien katagener Haarfollikel bei allen Tieren auch telogene Follikel vorzufinden. Wiederum drei Tage später ist annähernd der gleiche Befund zu erheben: neben späten Katagenfollikeln sind telogene Ruhefollikel vorhanden.
117 Eigene Untersuchungen 117 Im Alter von sechs Wochen (Tag 42) sind bei allen minderbehaarten Tieren alle Haarfollikelstadien nebeneinander vorzufinden. Neben einigen wenigen Anagenfollikeln finden sich vorwiegend katagene bis telogene Stadien. Bei den ältesten untersuchten Tieren (Tag 182) sind wiederum sowohl anagene, als auch katagene Follikel in der Haut vorhanden (siehe Abbildung 3.26, dünne Linie mit Kreisen und grau hinterlegte Fläche). Zusammenfssend ist festzustellen, daß bei den normal behaarten Tieren ein einheitlicher, synchronisierter, wellenförmiger Haarwachstumszyklus beobachtet werden kann. Bei den minderbehaarten sht/sht Mäusen liegt dagegen ein sehr heterogenes Bild vor: Bei einem einzigen Tier können in einem Hautareal mehrere Haarfollikelstadien gleichzeitig nebeneinander beobachtet werden, was bei dem normalen +/sht Genotyp nicht möglich ist. Hier findet sich innerhalb eines Hautareals immer nur ein einheitliches Zyklusstadium. Desweiteren liegt beim sht/sht Genotyp eine deutlich andersartige Geschwindigkeit des Haarzyklus vor. Katagene Haarfollikel können hier bereits wesentlich früher vorgefunden werden. Bereits im Alter von neun Lebenstagen sind katagene Follikel in der Haut der sht/sht Tiere zu sehen. Andererseits scheint diese Phase des Haarwachstums stark verlängert zu sein, denn katagene Follikel sind noch zu Zeitpunkten in der Haut der minderbeharten Mäuse vorhanden, zu denen bei den normal behaarten Tieren der Haarzyklus bereits vollständig abgeschlossen ist. Noch am letzten Tag des dann normalerweise beendeten ersten Haarzyklus (Lebenstag 21) sind bei den sht/sht Tieren katagene Follikel - neben telogenen Stadien - zu finden. Auch das Auftreten dieser katagenen Stadien bei den Altersstufen 42 und 182 Tage deutet auf eine ungewöhnliche Länge dieser Phase hin. Hierbei ist jedoch durch das Zeitschema der Probennahme nicht zu klären, ob sich diese Haarfollikel noch oder schon wieder im Katagen befinden.
118 118 Eigene Untersuchungen Abbildung 3.26: Schematische Darstellung der Haarfollikelzyklusstadien beider Genotypen im zeitlichen Verlauf Anagen sht/sht +/sht Katagen Telogen Lebensalter in Tagen Dichte der Haarfollikel Anhand der Dichte der Haarfollikelinfundibula in den histologischen Transversalschnitten wurde untersucht, ob der spärlichen Behaarung des sht/sht Genotyps möglicherweise zusätzlich zu der andersartigen Ausbildung der Haare auch eine Hypogenesie der Haarfollikel zugrunde liegt. Die Evaluierung der erfaßten Daten erfolgte auch hier durch eine mit linearen Modellen gerechnete Varianzanalyse (SAS, Procedure GLM) mit den fixen Effekten Geschlecht, Genotyp und Hautlokalisation (Signifikanzgrenze: p > 0,05). Eine Hypogenesie der Haarfollikel ist jedoch nicht festzustellen, denn die sht/sht Tiere beiderlei Geschlechts weisen innerhalb der Haut ihres Rückens sogar eine signifikant höhere Haarfollikeldichte auf (p = 0,003). Demgegenüber ist die Haarfollikeldichte der Bauchhaut bei den sht/sht Tieren bei beiden Geschlechtern geringer als bei den +/sht Tieren.
119 Eigene Untersuchungen 119 Die Haarfollikeldichte der Bauchhaut der sht/sht Tiere ist jedoch nicht signifikant geringer, als bei den normal behaarten Wurfgeschwistern (p = 0,47). Betrachtet man sowohl den Genotyp als auch das Geschlecht gemeinsam, liegt bei den männlichen und weiblichen sht/sht Tieren eine höhere Infundibulumsdichte vor. Diese Erhöhung der Dichte ist jedoch nicht signifikant ( : p = 0,50, : p = 0,07; siehe Tabelle 3.6). Unabhängig vom Genotyp weisen die weiblichen Tiere eine hochsignifikant höhere Haarfollikeldichte als die Böcke auf. Dies ist sowohl bei den minderbehaarten (sht/sht), als auch bei den normal behaarten Mäusen (+/sht) festzustellen (siehe Tabelle 3.5). Wie in Abbildung 3.29 und Tabelle 3.6 dargestellt, ist beiden Genotypen gemein, daß im Bereich des Rückens eine hochsignifikant höhere Follikeldichte als am Bauch festzustellen ist. Dieser Befund deckt sich mit Angaben aus der Literatur über die Behaarung der Maus (UHR 1984). Die Differenzierung nach Genotyp und Entnahmelokalisation (Rücken und Bauch) ergibt eine bei den sht/sht Tieren eine hochsignifikant höhere Follikeldichte in der Rückenhaut (p = 0,003). Im Bereich des Bauches ist allerdings eine nicht signifikant niedrigere Dichte bei den sht/sht Tieren feststellbar (siehe Tabelle 3.6). In den Transversalschnitten (Ebene distal der Talgdrüsenausführungsgänge) der sht/sht Tier ist die mittelgradige Infundibuladilatation und Follikelkeratose deutlich zu sehen. Die Haarfollikel sind ebenso, wie bei ihren normal behaarten Wurfgeschwistern, regelmäßig in Reihen quer zur Längsachse des Tieres angeordnet (siehe Abbildung 3.27und Abbildung 3.28 ).
120 120 Eigene Untersuchungen Abbildung 3.27: Transversalschnitt der Haut durch die Haarfollikelinfundibula (Pfeil) des +/sht Genotyps, Lebenstag 21. Die Haarfollikel sind in Reihen quer zur Längsachse des Tieres angeordnet. Die bei jüngeren Mäusen zu beobachtende Bildung von Follikelgruppen verliert sich mit zunehmendem Alter und ist hier kaum noch zu erkennen. Paraplasteinbettung, HE-Färbung, Eichstrich = 50 µm Abbildung 3.28: Transversalschnitt der Haut durch die Haarfollikelinfundibula (Pfeil) des sht/sht Genotyps, Lebenstag 21. Die Infundibula sind mittel- bis hochgradig dilatiert und mit Keratin- und Talgmassen angefüllt. Die Haarfollikel sind auch bei diesem Genotyp in Reihen quer zur Längsachse des Tieres angeordnet. Paraplasteinbettung, HE-Färbung, Eichstrich = 50 µm
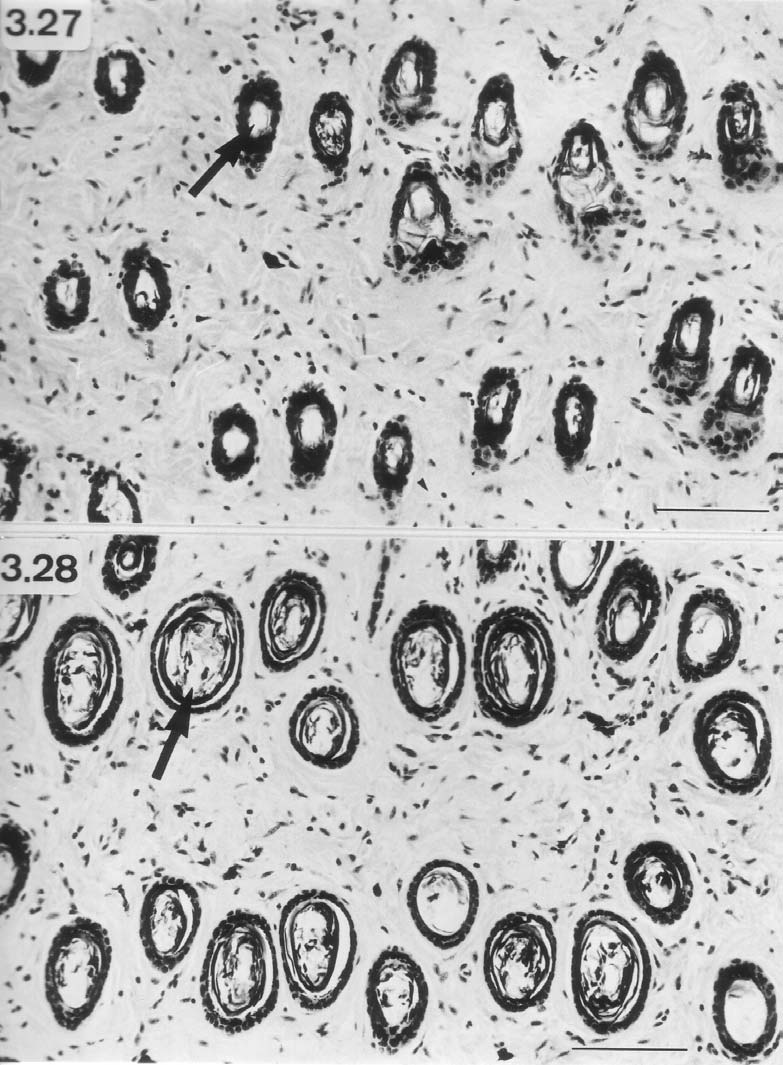
121 Eigene Untersuchungen 121
Dr. Chiara Noli, DVM, Dip ECVD
 Die Haut Ökosystem Haut: Dr. Chiara Noli, Dip ECVD Ospedale Veterinario Cuneese Borgo S. Dalmazzo (CN) - Italien Haut - Cutis Epidermis Dermis Unterhaut - Subcutis Adnexa Drüsen Haare Die Haut Bauch Haut
Die Haut Ökosystem Haut: Dr. Chiara Noli, Dip ECVD Ospedale Veterinario Cuneese Borgo S. Dalmazzo (CN) - Italien Haut - Cutis Epidermis Dermis Unterhaut - Subcutis Adnexa Drüsen Haare Die Haut Bauch Haut
Haut und Hautanhangsgebilde
 Kurstag 10 Haut und Hautanhangsgebilde Themen Leistenhaut Haare Hautdrüsen Brustdrüse Lernziele Prüfungsrelevante Lerninhalte Haut: Bauprinzip und funktionelle Bedeutung, Turgor, Pigmentierung, Leisten-
Kurstag 10 Haut und Hautanhangsgebilde Themen Leistenhaut Haare Hautdrüsen Brustdrüse Lernziele Prüfungsrelevante Lerninhalte Haut: Bauprinzip und funktionelle Bedeutung, Turgor, Pigmentierung, Leisten-
Mikroskopische Anatomie
 Mikroskopische Anatomie Haut und Anhangsorgane Herz und Blutgefäße Atmungstrakt Blut und Blutbildung Lymphatische Organe Endokrine Organe Auge Verdauungstrakt Harntrakt Genitalorgane Allgemeines: Gesamtfläche:
Mikroskopische Anatomie Haut und Anhangsorgane Herz und Blutgefäße Atmungstrakt Blut und Blutbildung Lymphatische Organe Endokrine Organe Auge Verdauungstrakt Harntrakt Genitalorgane Allgemeines: Gesamtfläche:
Harnkanälchen. Proximaler Tubulus
 Harnkanälchen Der Harnkanälchenapparat des Nephrons ist etwa 3-4 cm lang und gliedert sich in den proximalen, intermediäre und distalen Tubulus. Alle Tubuli bestehen aus einem einschichtigen Epithel, das
Harnkanälchen Der Harnkanälchenapparat des Nephrons ist etwa 3-4 cm lang und gliedert sich in den proximalen, intermediäre und distalen Tubulus. Alle Tubuli bestehen aus einem einschichtigen Epithel, das
Epi Histopathologische Untersuchung auf metastatische Aktivität
 49 In den Straßenformen waren sowohl das Stratum superficiale als auch das Stratum profundum des Koriums deutlich verbreitert und wurden von breiten Bindegewebssträngen durchzogen (Abb.12). iese bestanden
49 In den Straßenformen waren sowohl das Stratum superficiale als auch das Stratum profundum des Koriums deutlich verbreitert und wurden von breiten Bindegewebssträngen durchzogen (Abb.12). iese bestanden
Schattauer Schattauer Schattauer. Schattauer. Schattauer. 15 Allgemeine Körperdecke.
 15 Allgemeine Körperdecke 2010 GmbH, Stuttgart Haut Haare mit anliegenden Talgdrüsen Talgdrüsen Epidermis, mit abschilferndem Stratum corneum Stratum papillare exoepitheliale, exokrine, tubuläre, apokrine
15 Allgemeine Körperdecke 2010 GmbH, Stuttgart Haut Haare mit anliegenden Talgdrüsen Talgdrüsen Epidermis, mit abschilferndem Stratum corneum Stratum papillare exoepitheliale, exokrine, tubuläre, apokrine
Kompendium Basiswissen Haut. Aufbau Funktionen Größe
 Kompendium Basiswissen Haut Aufbau Funktionen Größe Aufbau der Haut Die Haut (dermatologisch auch Cutis oder Integument genannt) ist ein schichtig aufgebautes Organ. Aufbau der Haut Die Haut besteht aus
Kompendium Basiswissen Haut Aufbau Funktionen Größe Aufbau der Haut Die Haut (dermatologisch auch Cutis oder Integument genannt) ist ein schichtig aufgebautes Organ. Aufbau der Haut Die Haut besteht aus
Duale Reihe Dermatologie Wissen ist für alle für alle! A2K Free Access to Knowledge
 Duale Reihe Dermatologie Wissen ist für alle da @lles für alle! Selbst informieren und protestieren, z.b. gegen TRIPS: http://de.wikipedia.org/wiki/trips z.b. beim G8-Gipfel 2007 in Heiligendamm: www.g8-2007.de
Duale Reihe Dermatologie Wissen ist für alle da @lles für alle! Selbst informieren und protestieren, z.b. gegen TRIPS: http://de.wikipedia.org/wiki/trips z.b. beim G8-Gipfel 2007 in Heiligendamm: www.g8-2007.de
1 Anatomische und funktionelle Grundlagen
 1 Anatomische und funktionelle Grundlagen 1.1 Anatomischer Aufbau der Haut Die Haut (Kutis) besteht aus Epidermis und Dermis. Unterhalb der Dermis liegt das subkutane Fettgewebe (Subkutis), das streng
1 Anatomische und funktionelle Grundlagen 1.1 Anatomischer Aufbau der Haut Die Haut (Kutis) besteht aus Epidermis und Dermis. Unterhalb der Dermis liegt das subkutane Fettgewebe (Subkutis), das streng
Die den gesamten Körper eines Säugetieres kontinuierlich einhüllende. Schutz gegen mechanische, physikalische, chemische, biologische
 Allgemeine Körperdecke Die den gesamten Körper eines Säugetieres kontinuierlich einhüllende Grenz- und Kontaktschicht zur Umwelt Das größte Organ eines Säugetiers Vielfältige Aufgaben: Schutz gegen mechanische,
Allgemeine Körperdecke Die den gesamten Körper eines Säugetieres kontinuierlich einhüllende Grenz- und Kontaktschicht zur Umwelt Das größte Organ eines Säugetiers Vielfältige Aufgaben: Schutz gegen mechanische,
Ergebnisse der transmissionselektronenmikroskopischen (TEM) Untersuchungen
 Untersuchungen") 4 Ergebnisse 101 4.2.2. Ergebnisse der transmissionselektronenmikroskopischen (TEM) Untersuchungen Anhand der TEM- Aufnahmen sollte untersucht werden, welche Schäden Albendazolsulfoxid allein beziehungsweise
4 Ergebnisse 101 4.2.2. Ergebnisse der transmissionselektronenmikroskopischen (TEM) Untersuchungen Anhand der TEM- Aufnahmen sollte untersucht werden, welche Schäden Albendazolsulfoxid allein beziehungsweise
C. MUSKELGEWEBE : 4. Glattes Muskelgewebe (Harnblase, Ratte), Semi-Feinschnitt, Toluidinblau : 16. Glattes Muskelgewebe (Harnblase, Mensch), HE :
, Semi-Feinschnitt, Toluidinblau : 16. Glattes Muskelgewebe (Harnblase, Mensch), HE :") C. MUSKELGEWEBE : 4. Glattes Muskelgewebe (Harnblase, Ratte), Semi-Feinschnitt, Toluidinblau : - Der zigarrförmige Kern liegt zentral und scheint im Querschnitt rund. - Kleine längliche, spindelförmige
C. MUSKELGEWEBE : 4. Glattes Muskelgewebe (Harnblase, Ratte), Semi-Feinschnitt, Toluidinblau : - Der zigarrförmige Kern liegt zentral und scheint im Querschnitt rund. - Kleine längliche, spindelförmige
Schattauer Schattauer Schattauer. Schattauer. Schattauer. 2 Epithelgewebe.
 2 Epithelgewebe 2010 GmbH, Stuttgart Exokrine Drüsen exokrine Sekretgranula Lumen einer Schlauchdrüse (Querschnitt) mit Sekret exokrine Sekretgranula Exokrine Drüsen, Magen, Hund. Vergr. 5 500fach. ekkrine,
2 Epithelgewebe 2010 GmbH, Stuttgart Exokrine Drüsen exokrine Sekretgranula Lumen einer Schlauchdrüse (Querschnitt) mit Sekret exokrine Sekretgranula Exokrine Drüsen, Magen, Hund. Vergr. 5 500fach. ekkrine,
Normales histologisches Präparat mit Azan gefärbt, wodurch eine Differenzierung der azidophilen, basophilen und chromophoben Zellen möglich wird.
 Präparatedetails Organ Herkunft Färbung HYPOPHYSE MENSCH AZAN Methode Normales histologisches Präparat mit Azan gefärbt, wodurch eine Differenzierung der azidophilen, basophilen und chromophoben Zellen
Präparatedetails Organ Herkunft Färbung HYPOPHYSE MENSCH AZAN Methode Normales histologisches Präparat mit Azan gefärbt, wodurch eine Differenzierung der azidophilen, basophilen und chromophoben Zellen
Therapiebegleitende Hautpflege bei onkologischen Erkrankungen. Fachinformationen
 Therapiebegleitende Hautpflege bei onkologischen Erkrankungen. Fachinformationen Einleitung Aufgrund des antiproliferativen Charakters einer onkologischen Therapie kommt es beim Einsatz von klassischen
Therapiebegleitende Hautpflege bei onkologischen Erkrankungen. Fachinformationen Einleitung Aufgrund des antiproliferativen Charakters einer onkologischen Therapie kommt es beim Einsatz von klassischen
7. Diskussion. Rezeptoren bestimmt zu werden [3, 4, 8, 12, 95, 118, 119].
![7. Diskussion. Rezeptoren bestimmt zu werden [3, 4, 8, 12, 95, 118, 119].](/thumbs/34/13070185.jpg "7. Diskussion. Rezeptoren bestimmt zu werden [3, 4, 8, 12, 95, 118, 119].") 7. Diskussion Der Auf- und Abbau eines Haarfollikels erfordert eine exakt aufeinander abgestimmte Interaktion zwischen proliferierenden, differenzierenden und apoptotischen Zellen neuroektodermaler Herkunft
7. Diskussion Der Auf- und Abbau eines Haarfollikels erfordert eine exakt aufeinander abgestimmte Interaktion zwischen proliferierenden, differenzierenden und apoptotischen Zellen neuroektodermaler Herkunft
Learn4Med. Ein Gen steuert die Haarfarbe einer Katze. Es gibt ein Allel (also eine Version) für ein schwarzes Fell und ein Allel für rote Haare.
 für ein schwarzes Fell und ein Allel für rote Haare.") 1. Mendelsche Regeln Bei Mendel ist ein Gen als Teil des Erbmaterials definiert, der für die Ausbildung eines bestimmten Merkmals verantwortlich ist. Gibt es für dieses Gen verschiedene Ausprägungen, nennt
1. Mendelsche Regeln Bei Mendel ist ein Gen als Teil des Erbmaterials definiert, der für die Ausbildung eines bestimmten Merkmals verantwortlich ist. Gibt es für dieses Gen verschiedene Ausprägungen, nennt
B. Epithelgewebe : E. Niere (Mensch), Goldner : - Darstellung der Grenze zwischen Rinde (Cortex) und Mark (Medulla) : 1.
, Goldner : - Darstellung der Grenze zwischen Rinde (Cortex) und Mark (Medulla) : 1.") B. Epithelgewebe : E. Niere (Mensch), Goldner : - Darstellung der Grenze zwischen Rinde (Cortex) und Mark (Medulla) : 1. Nierenkörperchen sind nur im Cortex vorhanden 2. Aa. et Vv. Arcuata verlaufen auf
B. Epithelgewebe : E. Niere (Mensch), Goldner : - Darstellung der Grenze zwischen Rinde (Cortex) und Mark (Medulla) : 1. Nierenkörperchen sind nur im Cortex vorhanden 2. Aa. et Vv. Arcuata verlaufen auf
Haar. Welche Struktur hat das Haar? Wie wächst das Haar? befindet sich normalerweise in der Anagenphase.
 Welche Struktur hat das Haar? Das Haarorgan besteht im wesentlichen aus dem Haarschaft, der Haarpapille, den Matrix- Trichozyten und deren Tochterzellen, den Trichozyten. Im Zuge vielfältiger Prozesse
Welche Struktur hat das Haar? Das Haarorgan besteht im wesentlichen aus dem Haarschaft, der Haarpapille, den Matrix- Trichozyten und deren Tochterzellen, den Trichozyten. Im Zuge vielfältiger Prozesse
Dermatologie derma logos δερµα λογος
 Dermatologie derma logos δερµα λογος Dermato-* Histo pathologie Andrologie* Medikamentöse Tumortherapie* Dermatologie Infektionen Entzündliche ndliche Dermatosen Hauttumoren Genetische Erkrankungen Allergologie*
Dermatologie derma logos δερµα λογος Dermato-* Histo pathologie Andrologie* Medikamentöse Tumortherapie* Dermatologie Infektionen Entzündliche ndliche Dermatosen Hauttumoren Genetische Erkrankungen Allergologie*
3 Eigene Untersuchungen
 3 Eigene Untersuchungen - 56-3.2.5.1.2 Zitzen Es werden 65 unveränderte, gesunde Zitzen sonographisch dargestellt. Diese stammen von 24 Hündinnen. Die Zitze stellt sich in allen Fällen als ein queroval
3 Eigene Untersuchungen - 56-3.2.5.1.2 Zitzen Es werden 65 unveränderte, gesunde Zitzen sonographisch dargestellt. Diese stammen von 24 Hündinnen. Die Zitze stellt sich in allen Fällen als ein queroval
Nichtneoplastische Veränderungen, die nicht vom Gewebe der Mamma ausgehen
 3 Eigene Untersuchungen - 69-3.2.5.2.3 Nichtneoplastische Veränderungen, die nicht vom Gewebe der Mamma ausgehen 3.2.5.2.3.1 Lymphknoten Im Untersuchungsgut werden zwei palpierte und sonographisch dargestellte
3 Eigene Untersuchungen - 69-3.2.5.2.3 Nichtneoplastische Veränderungen, die nicht vom Gewebe der Mamma ausgehen 3.2.5.2.3.1 Lymphknoten Im Untersuchungsgut werden zwei palpierte und sonographisch dargestellte
Haut. 17.2 Dermis 312. 17.3 Hypodermis 313. 17.4 Anhangsgebilde der Haut 313 17.4.1 Haare 313 17.4.2 Nägel 315 17.4.3 Drüsen 316
 Haut.1 Epidermis (Oberhaut) 307.1.1 Keratinozyten Schichtung 307.1.2 Melanozyten Pigment 309.1.3 Langerhans-Zellen Immunabwehr 312.1.4 Merkel-Zellen Mechanosensoren 312.2 Dermis 312.3 Hypodermis 313.4
Haut.1 Epidermis (Oberhaut) 307.1.1 Keratinozyten Schichtung 307.1.2 Melanozyten Pigment 309.1.3 Langerhans-Zellen Immunabwehr 312.1.4 Merkel-Zellen Mechanosensoren 312.2 Dermis 312.3 Hypodermis 313.4
3 Ergebnisse. 3.1 Die Speckling-Domäne von Wt1 umfaßt die Aminosäuren
 3 Ergebnisse 3.1 Die Speckling-Domäne von Wt1 umfaßt die Aminosäuren 76-120 Die Existenz verschiedener Isoformen von WT1 ist unter anderem auf die Verwendung einer für die Aminosäuren KTS kodierenden alternativen
3 Ergebnisse 3.1 Die Speckling-Domäne von Wt1 umfaßt die Aminosäuren 76-120 Die Existenz verschiedener Isoformen von WT1 ist unter anderem auf die Verwendung einer für die Aminosäuren KTS kodierenden alternativen
Entwicklung der Tiere
 Entwicklung der Tiere Entwicklung beginnt, wie immer, mit einer ersten Zelle. Diese Zelle ist die befruchtete, diploide Zygote, deren Kern durch Verschmelzung der Kerne einer männlichen und einer weiblichen
Entwicklung der Tiere Entwicklung beginnt, wie immer, mit einer ersten Zelle. Diese Zelle ist die befruchtete, diploide Zygote, deren Kern durch Verschmelzung der Kerne einer männlichen und einer weiblichen
1. Benennen Sie die dargestellten Zellorganellen! 2. Beschreiben Sie jeweils den Aufbau! 3. Erläutern Sie jeweils kurz ihre Funktion!
 Sek.II Arbeitsblatt 1 Zellorganellen mit Doppelmembran 1. Benennen Sie die dargestellten Zellorganellen! 2. Beschreiben Sie jeweils den Aufbau! 3. Erläutern Sie jeweils kurz ihre Funktion! Zellkern Mitochondrium
Sek.II Arbeitsblatt 1 Zellorganellen mit Doppelmembran 1. Benennen Sie die dargestellten Zellorganellen! 2. Beschreiben Sie jeweils den Aufbau! 3. Erläutern Sie jeweils kurz ihre Funktion! Zellkern Mitochondrium
Wie funktioniert Muskelaufbau? Eine Reise in die Welt des Muskels.
 Wie funktioniert Muskelaufbau? Eine Reise in die Welt des Muskels. Wie funktioniert Muskelaufbau? Wie funktioniert Muskelaufbau also wirklich. Immer wieder hört man Märchen wie zum Beispiel, dass Muskeln
Wie funktioniert Muskelaufbau? Eine Reise in die Welt des Muskels. Wie funktioniert Muskelaufbau? Wie funktioniert Muskelaufbau also wirklich. Immer wieder hört man Märchen wie zum Beispiel, dass Muskeln
mehrschichtiges Plattenepithel mit einer
 Mehrschichtiges Plattenepithel Mensch Beim sorgfältigen Betrachten von Abb. 1 und 2 wird der Aufbau der Schleimhaut sichtbar. Die unterste Zelllage besteht aus gleichmäßig großen Zellen mit einem zentralen
Mehrschichtiges Plattenepithel Mensch Beim sorgfältigen Betrachten von Abb. 1 und 2 wird der Aufbau der Schleimhaut sichtbar. Die unterste Zelllage besteht aus gleichmäßig großen Zellen mit einem zentralen
Haarausfall Schlechte Haarqualität Vorzeitiges Ergrauen
 Haarausfall Schlechte Haarqualität Vorzeitiges Ergrauen Ein Ratgeber für Betroffene Merz Pharma (Schweiz) AG, Allschwil www.pantogar.ch Erste Anzeichen Vermehrt Haare in der Bürste oder im Waschbecken?
Haarausfall Schlechte Haarqualität Vorzeitiges Ergrauen Ein Ratgeber für Betroffene Merz Pharma (Schweiz) AG, Allschwil www.pantogar.ch Erste Anzeichen Vermehrt Haare in der Bürste oder im Waschbecken?
Bau des Nervengewebes
 Bau des Nervengewebes Das Nervengewebe hat eine zelluläre Gliederung und wird prinzipiell in die erregbaren Neuronen und die nicht erregbaren Zellen der Neuroglia unterteilt. Das Nervengewebe organisiert
Bau des Nervengewebes Das Nervengewebe hat eine zelluläre Gliederung und wird prinzipiell in die erregbaren Neuronen und die nicht erregbaren Zellen der Neuroglia unterteilt. Das Nervengewebe organisiert
+DDUDXVIDOOEHVLHJHQ 1XUZHUGLH8UVDFKHILQGHWNDQQGHQ+DDUDXVIDOOEHVHLWLJHQ
 +DDUDXVIDOOEHVLHJHQ 1XUZHUGLH8UVDFKHILQGHWNDQQGHQ+DDUDXVIDOOEHVHLWLJHQ Autorin: Eva Menderi www.gesundheits-ebooks.de günstige Bücher für Ihre Gesundheit! ,QKDOWVYHU]HLFKQLV Vorwort... Seite 3 Aufbau des
+DDUDXVIDOOEHVLHJHQ 1XUZHUGLH8UVDFKHILQGHWNDQQGHQ+DDUDXVIDOOEHVHLWLJHQ Autorin: Eva Menderi www.gesundheits-ebooks.de günstige Bücher für Ihre Gesundheit! ,QKDOWVYHU]HLFKQLV Vorwort... Seite 3 Aufbau des
Sammlung von elektronenmikroskopischen Aufnahmen
 Sammlung von elektronenmikroskopischen Aufnahmen Hilfsmaterial für die Prüfungen zusammengestellt von Prof. Dr. Pál Röhlich Zellkern Kernhülle Golgi-Apparat Transport- Vesikeln Mitochondrium Lysosom Mitochondrien
Sammlung von elektronenmikroskopischen Aufnahmen Hilfsmaterial für die Prüfungen zusammengestellt von Prof. Dr. Pál Röhlich Zellkern Kernhülle Golgi-Apparat Transport- Vesikeln Mitochondrium Lysosom Mitochondrien
Allgemeine Pathologie. Anpassungsreaktionen. Metaplasie
 Allgemeine Pathologie Anpassungsreaktionen Metaplasie Anpassungsreaktionen Was kann mit Zellen / Geweben geschehen? - sie können ungestört bis an ihr individuelles Ende leben und müssen dann ersetzt werden
Allgemeine Pathologie Anpassungsreaktionen Metaplasie Anpassungsreaktionen Was kann mit Zellen / Geweben geschehen? - sie können ungestört bis an ihr individuelles Ende leben und müssen dann ersetzt werden
Haarausfall? Dünner werdendes Haar? Ein Ratgeber für Patientinnen mit Haarausfall oder dünnem, brüchigem Haar
 Haarausfall? Dünner werdendes Haar? Ein Ratgeber für Patientinnen mit Haarausfall oder dünnem, brüchigem Haar Medizin für für Ihr Ihr Haar Haar Lebenszyklus der Haare 1 Die Wachstumsphase (Anagenphase)
Haarausfall? Dünner werdendes Haar? Ein Ratgeber für Patientinnen mit Haarausfall oder dünnem, brüchigem Haar Medizin für für Ihr Ihr Haar Haar Lebenszyklus der Haare 1 Die Wachstumsphase (Anagenphase)
Kenntnis der Ossifikation eines Röhrenknochens und der Strukturen eines Fingers
 Präparatedetails Organ Herkunft Färbung FINGER MENSCH, NEUGEBORENES HÄMALAUN EOSIN (HE) Methode Paraffinschnitte von Knochengewebe sind nur nach Entmineralisierung möglich. Dieses Präparat wurde jedoch
Präparatedetails Organ Herkunft Färbung FINGER MENSCH, NEUGEBORENES HÄMALAUN EOSIN (HE) Methode Paraffinschnitte von Knochengewebe sind nur nach Entmineralisierung möglich. Dieses Präparat wurde jedoch
Anhang zu Kapitel 06.02: Die Zelle
 Anhang zu Kapitel 06.02: Die Zelle Inhalt Anhang Kapitel 06.02: Die Zelle... 1 Inhalt... 2 Zellorganellen im EM: die Zellmembran... 3 Zellkern einer Leberzelle... 4 Zellkern... 4 Poren der Kernmembran...
Anhang zu Kapitel 06.02: Die Zelle Inhalt Anhang Kapitel 06.02: Die Zelle... 1 Inhalt... 2 Zellorganellen im EM: die Zellmembran... 3 Zellkern einer Leberzelle... 4 Zellkern... 4 Poren der Kernmembran...
14 11 lle Ze a b c d e f g h 15
 14 11 a b c d 15 e f g h 12 16 Mitose Bei der Kernteilung der Euzyten, dermitose, werden die Chromosomen für die späteren Tochterkerne aufgeteilt ( 14). Unsere Abbildung zeigt einen sich teilenden Fibroblasten
14 11 a b c d 15 e f g h 12 16 Mitose Bei der Kernteilung der Euzyten, dermitose, werden die Chromosomen für die späteren Tochterkerne aufgeteilt ( 14). Unsere Abbildung zeigt einen sich teilenden Fibroblasten
Die Oberflächenmodifikationen der Lederhaut im Wandsegment
 Abbildungen 185 Abbildung 10 Die Oberflächenmodifikationen der Lederhaut im Wandsegment Abb. 10 a: Proximaler Bereich des Wandsegmentes Im Grenzbereich vom Kron- zum Wandsegment entspringen die proximalen
Abbildungen 185 Abbildung 10 Die Oberflächenmodifikationen der Lederhaut im Wandsegment Abb. 10 a: Proximaler Bereich des Wandsegmentes Im Grenzbereich vom Kron- zum Wandsegment entspringen die proximalen
Entzündung. Teil 18.
 Teil 18 www.patho.vetmed.uni-muenchen.de/matnew.html Proliferative Entzündungen: Granulomatöse Entzündung Unterschiede granulierende - granulomatöse Entzündung Mononukleäres Phagozyten System (MPS) Dendritische
Teil 18 www.patho.vetmed.uni-muenchen.de/matnew.html Proliferative Entzündungen: Granulomatöse Entzündung Unterschiede granulierende - granulomatöse Entzündung Mononukleäres Phagozyten System (MPS) Dendritische
Histopathologie. Fall 30
 Histopathologie Fall 30 Präp.-Nr.: 30 Färbung: HE Organ: Knochen Tierart: Hund Allgemeines (1): Osteosarkome kommen in unterschiedlichen Lokalisationen vor Axiales Skelett: - Wirbelsäule - Rippen - Brustbein
Histopathologie Fall 30 Präp.-Nr.: 30 Färbung: HE Organ: Knochen Tierart: Hund Allgemeines (1): Osteosarkome kommen in unterschiedlichen Lokalisationen vor Axiales Skelett: - Wirbelsäule - Rippen - Brustbein
Skelettmuskulatur (M.uvulae) quer/längs geschnitten
 quer/längs geschnitten") Mundhöhle, Zunge Kurspräparate: Nr.50 Uvula Nr.39 Zunge, Papillae fungiformes Nr.40 Zungenmitte Nr.42 Zunge, Papillae foliatae Nr.44 Zunge, Papillae vallatae Nr.86 Harter Gaumen Uvula- weicher Gaumen Präparat
Mundhöhle, Zunge Kurspräparate: Nr.50 Uvula Nr.39 Zunge, Papillae fungiformes Nr.40 Zungenmitte Nr.42 Zunge, Papillae foliatae Nr.44 Zunge, Papillae vallatae Nr.86 Harter Gaumen Uvula- weicher Gaumen Präparat
Die Leber Entstehung der Leber. Histologie der Leber
 Die Leber Entstehung der Leber Die Hepatozyten sind endodermaler Herkunft und entwickeln sich aus den selben Vorläuferzellen, die auch den Darm bilden. Bindegewebe und Gefäße der Leber entstehen aus mesenchymalen
Die Leber Entstehung der Leber Die Hepatozyten sind endodermaler Herkunft und entwickeln sich aus den selben Vorläuferzellen, die auch den Darm bilden. Bindegewebe und Gefäße der Leber entstehen aus mesenchymalen
bei den japanischen Von Aus dem Anatomischen Institut der Keio Universitat, Tokyo. (Direktor: Prof. Dr. T. Taniguchi). 13 Textfiguren.
. 13 Textfiguren.") Das Vorkommen und die Verteilung des M. arrector pili in der Augenlidhaut bei den japanischen Feten Von Masuo Ikeda Aus dem Anatomischen Institut der Keio Universitat, Tokyo. (Direktor: Prof. Dr. T. Taniguchi).
Das Vorkommen und die Verteilung des M. arrector pili in der Augenlidhaut bei den japanischen Feten Von Masuo Ikeda Aus dem Anatomischen Institut der Keio Universitat, Tokyo. (Direktor: Prof. Dr. T. Taniguchi).
Der gleichzeitige Anti-Haarverlust und Haar Wachstums Complex
 Capixyl TM Der gleichzeitige Anti-Haarverlust und Haar Wachstums Complex Wie wichtig Haare in unserem Leben sind, kann nicht oft genug erwähnt werden. Wenn Männer, aber auch Frauen ihre Haare verlieren,
Capixyl TM Der gleichzeitige Anti-Haarverlust und Haar Wachstums Complex Wie wichtig Haare in unserem Leben sind, kann nicht oft genug erwähnt werden. Wenn Männer, aber auch Frauen ihre Haare verlieren,
Gewebe und Organe. Keimblätter als Basis für die Gewebebildung
 Gewebe und Organe Keimblätter als Basis für die Gewebebildung 1 Gewebe: Verbände gleichartig differenzierter Zelle und ihrer Abkömmlinge Echte Gewebe Bildungsgewebe (Meristem) Teilungsfähige Embryonalzellen
Gewebe und Organe Keimblätter als Basis für die Gewebebildung 1 Gewebe: Verbände gleichartig differenzierter Zelle und ihrer Abkömmlinge Echte Gewebe Bildungsgewebe (Meristem) Teilungsfähige Embryonalzellen
4 Ergebnisse. 4.1 Evaluierung der Methode. 4.2 Allgemeine Beobachtungen. 4.3 CD68-positive Zellen 4 ERGEBNISSE 17
 4 ERGEBNISSE 17 4 Ergebnisse 4.1 Evaluierung der Methode Variationskoeffizient Zur Ermittlung der Messgenauigkeit wurde der mittlere Variationskoeffizient bestimmt [5]. Zur Bestimmung wurden 18 Präparate
4 ERGEBNISSE 17 4 Ergebnisse 4.1 Evaluierung der Methode Variationskoeffizient Zur Ermittlung der Messgenauigkeit wurde der mittlere Variationskoeffizient bestimmt [5]. Zur Bestimmung wurden 18 Präparate
Experimentelle Untersuchungen uber die Umwandlungs= fahigkeit der Zellen im Subkutangewebe der Maus.
 Experimentelle Untersuchungen uber die Umwandlungs= fahigkeit der Zellen im Subkutangewebe der Maus. V. Versuche mit grauer Salbe. Von Jushiro Mukohata. (Aus dem Anatomischen Institut der Medizinischen
Experimentelle Untersuchungen uber die Umwandlungs= fahigkeit der Zellen im Subkutangewebe der Maus. V. Versuche mit grauer Salbe. Von Jushiro Mukohata. (Aus dem Anatomischen Institut der Medizinischen
Allgemeine Pathologie
 Allgemeine Pathologie Störungen Eiweißstoffwechsel Störungen der Verhornung Störungen der Verhornung Normale Histologie (1): - äußere Haut besteht aus der Epidermis (Epithel), die auf einer Basalmembran
Allgemeine Pathologie Störungen Eiweißstoffwechsel Störungen der Verhornung Störungen der Verhornung Normale Histologie (1): - äußere Haut besteht aus der Epidermis (Epithel), die auf einer Basalmembran
Binde- und Fettgewebe
 Kurstag 4 Binde- und Fettgewebe Lernziele Prüfungsrelevante Lerninhalte Prinzipieller Aufbau, Klassifikation (Mesenchym, lockeres, retikuläres, retikulärfaseriges, straffes geflechtartiges, straffes parallelfaseriges
Kurstag 4 Binde- und Fettgewebe Lernziele Prüfungsrelevante Lerninhalte Prinzipieller Aufbau, Klassifikation (Mesenchym, lockeres, retikuläres, retikulärfaseriges, straffes geflechtartiges, straffes parallelfaseriges
Testis Nebenhoden Ductus deferens Prostata. Vorlesung Histologie. Die Studierenden sollen
 Kurstag 19 Männliche Geschlechtsorgane Themen Testis Nebenhoden Ductus deferens Prostata Lernziele Prüfungsrelevante Lerninhalte Männliche Geschlechtsorgane Hoden: Tubuli seminiferi, Keimepithel: Sertolizellen,
Kurstag 19 Männliche Geschlechtsorgane Themen Testis Nebenhoden Ductus deferens Prostata Lernziele Prüfungsrelevante Lerninhalte Männliche Geschlechtsorgane Hoden: Tubuli seminiferi, Keimepithel: Sertolizellen,
Unterschied Tiere, Pflanzen, Bakterien u. Pilze und die Zellorganellen
 Unterschied Tiere, Pflanzen, Bakterien u. Pilze und die Zellorganellen Die Organellen der Zelle sind sozusagen die Organe die verschiedene Funktionen in der Zelle ausführen. Wir unterscheiden Tierische
Unterschied Tiere, Pflanzen, Bakterien u. Pilze und die Zellorganellen Die Organellen der Zelle sind sozusagen die Organe die verschiedene Funktionen in der Zelle ausführen. Wir unterscheiden Tierische
Das EM Bild der Zelle
 Übersicht 1 Übersicht Zellinneres ist durch Membranen unterteilt 1 Übersicht Zellinneres ist durch Membranen unterteilt Die Zelle ist kompartimentiert und ermöglicht dadurch den Ablauf unterschiedlicher
Übersicht 1 Übersicht Zellinneres ist durch Membranen unterteilt 1 Übersicht Zellinneres ist durch Membranen unterteilt Die Zelle ist kompartimentiert und ermöglicht dadurch den Ablauf unterschiedlicher
Wie viele Haare haben Sie am Kopf?
 Wie viele Haare haben Sie am Kopf? Die Anzahl der Haare hängt vom Haartyp ab Die natürliche Farbe des menschlichen Haares wird durch die in die Schuppenschicht (Kortex) eingestreuten braun-schwarzen bis
Wie viele Haare haben Sie am Kopf? Die Anzahl der Haare hängt vom Haartyp ab Die natürliche Farbe des menschlichen Haares wird durch die in die Schuppenschicht (Kortex) eingestreuten braun-schwarzen bis
Aus der Klinik und Poliklinik für Hals-, Nasen- und Ohren- Krankheiten, plastische und ästhetische Operationen. der Universität Würzburg
 Aus der Klinik und Poliklinik für Hals-, Nasen- und Ohren- Krankheiten, plastische und ästhetische Operationen der Universität Würzburg Direktor: Professor Dr. Dr. h.c. R. Hagen Untersuchungen zur optimalen
Aus der Klinik und Poliklinik für Hals-, Nasen- und Ohren- Krankheiten, plastische und ästhetische Operationen der Universität Würzburg Direktor: Professor Dr. Dr. h.c. R. Hagen Untersuchungen zur optimalen
Unterrichtsmaterialien in digitaler und in gedruckter Form. Auszug aus: Lernwerkstatt: Genetik & Vererbung. Das komplette Material finden Sie hier:
 Unterrichtsmaterialien in digitaler und in gedruckter Form Auszug aus: : Genetik & Vererbung Das komplette Material finden Sie hier: Download bei School-Scout.de Inhalt Vorwort Seite 4 Einleitung Seite
Unterrichtsmaterialien in digitaler und in gedruckter Form Auszug aus: : Genetik & Vererbung Das komplette Material finden Sie hier: Download bei School-Scout.de Inhalt Vorwort Seite 4 Einleitung Seite
Haarausfall? Patienteninformation. Zuverlässig wirksam gegen Haarausfall
 Haarausfall? Patienteninformation Zuverlässig wirksam gegen Haarausfall Aufbau des Haares Haarausfall viel mehr als nur ein optisches Problem Oberhaut Lederhaut Haarbalg Talgdrüse Haaraufrichtemuskel Wir
Haarausfall? Patienteninformation Zuverlässig wirksam gegen Haarausfall Aufbau des Haares Haarausfall viel mehr als nur ein optisches Problem Oberhaut Lederhaut Haarbalg Talgdrüse Haaraufrichtemuskel Wir
Chemie des Haares HAARE. a-helix-struktur der Haarproteine. Protein-Helix. Wasserstoffbrücke. b-ketten- oder Faltblatt-Struktur der Haarproteine
 Chemie des Haares Das Haar besteht zu 90 % aus dicht aneinander gereihten Zellen, die Keratin ein Eiweiß enthalten. Keratin wiederum besteht aus Aminosäuren, die zu langen Ketten (= Polypeptidketten) aneinander
Chemie des Haares Das Haar besteht zu 90 % aus dicht aneinander gereihten Zellen, die Keratin ein Eiweiß enthalten. Keratin wiederum besteht aus Aminosäuren, die zu langen Ketten (= Polypeptidketten) aneinander
Gegen Haarausfall gibt es ein Mittel.
 Gegen Haarausfall gibt es ein Mittel. Ursachen Therapie Tipps Ihre Chance, den Haarausfall in den Griff zu bekommen. Guten Tag! Heute wird vielleicht das erste Mal sein, dass Sie in den Spiegel schauen
Gegen Haarausfall gibt es ein Mittel. Ursachen Therapie Tipps Ihre Chance, den Haarausfall in den Griff zu bekommen. Guten Tag! Heute wird vielleicht das erste Mal sein, dass Sie in den Spiegel schauen
einschichtiges, hochprismatisches Epithel des Epithelium mucosae Tunica mucosa Lamina propria mucosae mit tubulären Eigendrüsen
 einschichtiges, hochprismatisches Epithel des Epithelium mucosae mit tubulären Eigendrüsen Tunica mucosa Blutgefäße mit Blut angefüllt Lamina muscularis mucosae Verdauungsapparat Drüsenmagen, Eigendrüsen,
einschichtiges, hochprismatisches Epithel des Epithelium mucosae mit tubulären Eigendrüsen Tunica mucosa Blutgefäße mit Blut angefüllt Lamina muscularis mucosae Verdauungsapparat Drüsenmagen, Eigendrüsen,
2. Zusammenfassung der eigene Arbeiten
 15 2. Zusammenfassung der eigene Arbeiten 2.1. Kontinuierliche postnatale plastische Veränderungen in der Innervation und Neurotrophinexpression der Haut Während der Morphogenese der murinen Rückenhaut,
15 2. Zusammenfassung der eigene Arbeiten 2.1. Kontinuierliche postnatale plastische Veränderungen in der Innervation und Neurotrophinexpression der Haut Während der Morphogenese der murinen Rückenhaut,
1.1 Eine komplexe Architektur Ein Organ mit zahlreichen Funktionen Ein Kommunikationsmodell 2
 sverzeichnis 1. Einleitung 1.1 Eine komplexe Architektur 1 1.2 Ein Organ mit zahlreichen Funktionen 1 1.3 Ein Kommunikationsmodell 2 Struktur und Physiologie der Haut Die Epidermis 2.1 Allgemeiner Aufbau
sverzeichnis 1. Einleitung 1.1 Eine komplexe Architektur 1 1.2 Ein Organ mit zahlreichen Funktionen 1 1.3 Ein Kommunikationsmodell 2 Struktur und Physiologie der Haut Die Epidermis 2.1 Allgemeiner Aufbau
Abb. 27: Durasinus mit Erythrozyten (TEM). kf kollagene Fibrillen, BM Basalmembran, En Endothel mit Vesikeln, E Erythrozyten, F Fibrozyt, v Mikrovilli
. kf kollagene Fibrillen, BM Basalmembran, En Endothel mit Vesikeln, E Erythrozyten, F Fibrozyt, v Mikrovilli") 66 Die Dura mater wird von Gefäßen durchzogen, wobei Venen häufiger als Arterien anzutreffen sind. Die Gefäße sind mit einem flachen, kontinuierlichen Endothel ausgekleidet, das einer kontinuierlichen
66 Die Dura mater wird von Gefäßen durchzogen, wobei Venen häufiger als Arterien anzutreffen sind. Die Gefäße sind mit einem flachen, kontinuierlichen Endothel ausgekleidet, das einer kontinuierlichen
3.2.1 Rassenverteilung der untersuchten Tiere
 3 Eigene Untersuchungen - 34-3.2 Ergebnisse Die Ergebnisse dieser Untersuchungen sind im Anhang in tabellarischer Form zusammengefasst. 3.2.1 Rassenverteilung der untersuchten Tiere Die für diese Arbeit
3 Eigene Untersuchungen - 34-3.2 Ergebnisse Die Ergebnisse dieser Untersuchungen sind im Anhang in tabellarischer Form zusammengefasst. 3.2.1 Rassenverteilung der untersuchten Tiere Die für diese Arbeit
Abbildungen 197. Abbildung 16
 Abbildungen 197 Abbildung 16 Die dermoepidermale Grenzfläche und Architektur des Zellverbundes vom epidermalen Teil des Klauenbeinträgers im mittleren und distalen Bereich des Wandsegmentes (Horizontschnitte)
Abbildungen 197 Abbildung 16 Die dermoepidermale Grenzfläche und Architektur des Zellverbundes vom epidermalen Teil des Klauenbeinträgers im mittleren und distalen Bereich des Wandsegmentes (Horizontschnitte)
Porcine Intestinale Adenomatose PIA
 Porcine Intestinale Adenomatose PIA Ätiologie: - bis 1994 wurden Campylobacter-ähnliche Bakterien verantwortlich gemacht (z.b. C. mucosalis), diese Bakterien wurden z.t. überhaupt erst im Zusammenhang
Porcine Intestinale Adenomatose PIA Ätiologie: - bis 1994 wurden Campylobacter-ähnliche Bakterien verantwortlich gemacht (z.b. C. mucosalis), diese Bakterien wurden z.t. überhaupt erst im Zusammenhang
Was versteht man unter Polyneuropathie?
 7 2 Was versteht man unter Polyneuropathie? Udo Zifko 2.1 Unser Nervensystem: Anatomie und Physiologie 8 2.1.1 Länge des Nervs 8 2.1.2 Dicke des Nervs 10 2.1.3 Struktur des Nervs 10 2.2 Arten der Polyneuropathie
7 2 Was versteht man unter Polyneuropathie? Udo Zifko 2.1 Unser Nervensystem: Anatomie und Physiologie 8 2.1.1 Länge des Nervs 8 2.1.2 Dicke des Nervs 10 2.1.3 Struktur des Nervs 10 2.2 Arten der Polyneuropathie
Biologie I/B: Klassische und molekulare Genetik, molekulare Grundlagen der Entwicklung Theoretische Übungen SS 2016
 Biologie I/B: Klassische und molekulare Genetik, molekulare Grundlagen der Entwicklung Theoretische Übungen SS 2016 Fragen für die Übungsstunde 4 (20.06. 24.06.) Regulation der Transkription II, Translation
Biologie I/B: Klassische und molekulare Genetik, molekulare Grundlagen der Entwicklung Theoretische Übungen SS 2016 Fragen für die Übungsstunde 4 (20.06. 24.06.) Regulation der Transkription II, Translation
Modul Biologische Grundlagen Kapitel I.2 Grundbegriffe der Genetik
 Frage Was sind Fachbegriffe zum Thema Grundbegriffe der Genetik? Antwort - Gene - Genotyp - Phänotyp - Genom - Dexoxyribonucleinsäure - Träger genetischer Information - Nukleotide - Basen - Peptid - Start-Codon
Frage Was sind Fachbegriffe zum Thema Grundbegriffe der Genetik? Antwort - Gene - Genotyp - Phänotyp - Genom - Dexoxyribonucleinsäure - Träger genetischer Information - Nukleotide - Basen - Peptid - Start-Codon
Vorlesungsinhalt. Bau der Pflanzenzelle. Einführung Entstehung des Lebens Organisationstufen der Pflanzen Stellung im Ökosystem
 Vorlesungsinhalt Einführung Entstehung des Lebens Organisationstufen der Pflanzen Stellung im Ökosystem Bau der Pflanzenzelle Anatomie, Entwicklung und Funktion der Pflanzenorgane - Gewebe - Primärer Pflanzenkörper
Vorlesungsinhalt Einführung Entstehung des Lebens Organisationstufen der Pflanzen Stellung im Ökosystem Bau der Pflanzenzelle Anatomie, Entwicklung und Funktion der Pflanzenorgane - Gewebe - Primärer Pflanzenkörper
5. HAARWACHSTUM. FIFCO Hair 4. KOPFHAUT TIEFENREINIGUNG ALLGEMEINE INFORMATIONEN ZUR HAARWACHSTUM -STUDIE 3. KREISRUNDER HAARAUSFALL 2.
 ALLGEMEINE INFORMATIONEN ZUR HAARWACHSTUM -STUDIE 1. DÜNNER WERDENDES HAAR 2. HAARAUSFALL 3. KREISRUNDER HAARAUSFALL 4. KOPFHAUT TIEFENREINIGUNG 5. HAARWACHSTUM VORHER NACHHER SCHÖNHEITSINSTITUT WWW.FIF-CO.DE
ALLGEMEINE INFORMATIONEN ZUR HAARWACHSTUM -STUDIE 1. DÜNNER WERDENDES HAAR 2. HAARAUSFALL 3. KREISRUNDER HAARAUSFALL 4. KOPFHAUT TIEFENREINIGUNG 5. HAARWACHSTUM VORHER NACHHER SCHÖNHEITSINSTITUT WWW.FIF-CO.DE
Die Männer-Medizin gegen Haarausfall *
 Rette Deine Haarwurzel so lange sie noch lebt! Stoppt Haarausfall * Stoppt Geheimratsecken ALOPEXY 5 % Lösung Die Männer-Medizin gegen Haarausfall * * Bei anlagebedingtem Haarausfall des Mannes OPEXY-Patientenbroschüre_1_2015_RZ.indd
Rette Deine Haarwurzel so lange sie noch lebt! Stoppt Haarausfall * Stoppt Geheimratsecken ALOPEXY 5 % Lösung Die Männer-Medizin gegen Haarausfall * * Bei anlagebedingtem Haarausfall des Mannes OPEXY-Patientenbroschüre_1_2015_RZ.indd
4 Ergebnisse / -/- Tag 1. Tag 2. Tag 3. Tag 5. Tag 7. Tag 10. Tag 14
 4.3.4 Mikroskopische Analyse der Wundheilung Bei der oberflächlichen Betrachtung der Wunden lässt sich kein signifikanter Unterschied feststellen. Allerdings kann ein eventuell mikroskopischer Unterschied
4.3.4 Mikroskopische Analyse der Wundheilung Bei der oberflächlichen Betrachtung der Wunden lässt sich kein signifikanter Unterschied feststellen. Allerdings kann ein eventuell mikroskopischer Unterschied
Stützgewebe 1/3. Knorpel
 Stützgewebe 1/3 Knorpel Knorpel Knorpelzellen Matrix Chondroblasten Chondrozyten Wasser (80%) Kollagen Typ II (Hauptanteil) Kollagen Typ IX (verbindet Typ II Fibrillen) Kollagen Typ X Kollagen Typ XI (Teil
Stützgewebe 1/3 Knorpel Knorpel Knorpelzellen Matrix Chondroblasten Chondrozyten Wasser (80%) Kollagen Typ II (Hauptanteil) Kollagen Typ IX (verbindet Typ II Fibrillen) Kollagen Typ X Kollagen Typ XI (Teil
Timing der embryologischen Stadien
 Blastogenese Embryogenese - Fetogenese Blastogenese: ca. 8 Tage speziesübergreifend Embryogenese: Embroygenese Trächtigkeitsdauer = sinkt mit der Länge der Trächtigkeit Fetogenese: Fetogenese = steigt
Blastogenese Embryogenese - Fetogenese Blastogenese: ca. 8 Tage speziesübergreifend Embryogenese: Embroygenese Trächtigkeitsdauer = sinkt mit der Länge der Trächtigkeit Fetogenese: Fetogenese = steigt
Matthias Birnstiel Modul Nervensystem Medizinisch wissenschaftlicher Lehrgang Wissenschaftliche Lehrmittel, Medien, Aus- und Weiterbildung
 Matthias Birnstiel Modul Nervensystem Medizinisch wissenschaftlicher Lehrgang CHRISANA Wissenschaftliche Lehrmittel, Medien, Aus- und Weiterbildung Inhaltsverzeichnis des Moduls Nervensystem Anatomie des
Matthias Birnstiel Modul Nervensystem Medizinisch wissenschaftlicher Lehrgang CHRISANA Wissenschaftliche Lehrmittel, Medien, Aus- und Weiterbildung Inhaltsverzeichnis des Moduls Nervensystem Anatomie des
in vivo -- Das Magazin der Deutschen Krebshilfe vom
 Seite 1/5 in vivo -- Das Magazin der Deutschen Krebshilfe vom 13.10.2009 Expertengespräch Weißer Hautkrebs Und zu diesem Thema begrüßen wir Prof. Dr. Cornelia Mauch, Oberärztin für dermatologische Onkologie
Seite 1/5 in vivo -- Das Magazin der Deutschen Krebshilfe vom 13.10.2009 Expertengespräch Weißer Hautkrebs Und zu diesem Thema begrüßen wir Prof. Dr. Cornelia Mauch, Oberärztin für dermatologische Onkologie
Zelle (Biologie) (http://www.youtube.com/watch?v=kxslw1lmvgk)
 (http://www.youtube.com/watch?v=kxslw1lmvgk)") Zelle (Biologie) 1. Allgemeines 2. Aufbau von Zellen 2.1 Unterschiede zwischen prokaryotischen und eukaryotischen Zellen 2.2 Unterschiede zwischen tierischen und pflanzlichen Zellen 2.3 Aufbau der pflanzlichen
Zelle (Biologie) 1. Allgemeines 2. Aufbau von Zellen 2.1 Unterschiede zwischen prokaryotischen und eukaryotischen Zellen 2.2 Unterschiede zwischen tierischen und pflanzlichen Zellen 2.3 Aufbau der pflanzlichen
Unser grösstes Sinnesorgan
 Die Haut Unser grösstes Sinnesorgan Die Haut Unser grösstes Sinnesorgan Die Hautoberfläche eines Menschen beträgt ungefähr 2 m 2. Dies ist etwa gleich gross wie eine Matratze. Mit Hilfe verschiedener Sinneszellen
Die Haut Unser grösstes Sinnesorgan Die Haut Unser grösstes Sinnesorgan Die Hautoberfläche eines Menschen beträgt ungefähr 2 m 2. Dies ist etwa gleich gross wie eine Matratze. Mit Hilfe verschiedener Sinneszellen
Das Zytoskelett. Struktur der Intermediärfilamente. Polymerisation/ Depolymerisation. Einteilung der Intermediärfilamente
 Das Zytoskelett Struktur der Intermediärfilamente Polymerisation/ Depolymerisation Einteilung der Intermediärfilamente Zell-Zell- und Zell-Matrix-Verbindungen Einteilung der Zytoskelettsysteme Aktin-Mikrofilamente
Das Zytoskelett Struktur der Intermediärfilamente Polymerisation/ Depolymerisation Einteilung der Intermediärfilamente Zell-Zell- und Zell-Matrix-Verbindungen Einteilung der Zytoskelettsysteme Aktin-Mikrofilamente
Die Physiologie des Muskels. Inhalt
 Die Physiologie des Muskels Inhalt Die Einteilung der Muskulatur Der Bau des Skelettmuskels Die molekularen Grundlagen der Muskelkontraktion Die Muskelkraft Die Muskelfasertypen Die Einteilung der Muskulatur
Die Physiologie des Muskels Inhalt Die Einteilung der Muskulatur Der Bau des Skelettmuskels Die molekularen Grundlagen der Muskelkontraktion Die Muskelkraft Die Muskelfasertypen Die Einteilung der Muskulatur
Fachinformation Rassekaninchenzucht oder 1
 In diesem Beitrag geht der Autor auf die Zusammenhänge des Fellhaars unserer Kaninchen ein, welches sich aus unterschiedlichen Haartypen zusammensetzt. Allgemeines Die Haut bildet einen Abschluss des Tierkörpers
In diesem Beitrag geht der Autor auf die Zusammenhänge des Fellhaars unserer Kaninchen ein, welches sich aus unterschiedlichen Haartypen zusammensetzt. Allgemeines Die Haut bildet einen Abschluss des Tierkörpers
Diabetes & Dialyse. Ein Praxiskompendium. von Werner Kleophas. 1. Auflage. Thieme 2000
 Diabetes & Dialyse Ein Praxiskompendium von Werner Kleophas. Auflage Thieme 000 Verlag C.H. Beck im Internet: www.beck.de ISBN 978 0558 Zu Inhaltsverzeichnis schnell und portofrei erhältlich bei beck-shop.de
Diabetes & Dialyse Ein Praxiskompendium von Werner Kleophas. Auflage Thieme 000 Verlag C.H. Beck im Internet: www.beck.de ISBN 978 0558 Zu Inhaltsverzeichnis schnell und portofrei erhältlich bei beck-shop.de
Unsere Haare. Jedes Haar hat seinen eigenen Zyklus. Anzahl der Haare Blond: Schwarz: Brünett: Rot:
 Haarexperte/ -in Unsere Haare Anzahl der Haare Blond: 150.000 Schwarz: 110.000 Brünett: 100.000 Rot: 75.000 Haardichte: 200/cm 2 Haarwachstum pro Tag: 0,3 mm Haardurchmesser: 0,1 mm Jedes Haar hat seinen
Haarexperte/ -in Unsere Haare Anzahl der Haare Blond: 150.000 Schwarz: 110.000 Brünett: 100.000 Rot: 75.000 Haardichte: 200/cm 2 Haarwachstum pro Tag: 0,3 mm Haardurchmesser: 0,1 mm Jedes Haar hat seinen
Bindegewebe Zellen und Extrazellulärmatrix (EZM)
") Bindegewebe Zellen und Extrazellulärmatrix (EZM) Ortsständige Zellen: Fibroblast ist für Stoffwechsel der Extrazellulärmatrix-Bestandteile zuständig (Kollagenfibrillen, elastische Fasern, retikuläre Fasern,
Bindegewebe Zellen und Extrazellulärmatrix (EZM) Ortsständige Zellen: Fibroblast ist für Stoffwechsel der Extrazellulärmatrix-Bestandteile zuständig (Kollagenfibrillen, elastische Fasern, retikuläre Fasern,
In jedem Zellkern befinden sich 46 Chromosomen, auf denen die Erbanlagen eines Menschen festgelegt sind.
 Zelle kleinste Einheit eines lebenden Organismus Eigenschaften Stoffwechsel Größe einer Zelle 5-20 Mikrometer Wachstum menschliche Eizelle 0,15 Millimeter Vermehrung, Vererbung kann sterben Bewegung Alterungsprozesse
Zelle kleinste Einheit eines lebenden Organismus Eigenschaften Stoffwechsel Größe einer Zelle 5-20 Mikrometer Wachstum menschliche Eizelle 0,15 Millimeter Vermehrung, Vererbung kann sterben Bewegung Alterungsprozesse
Glia- sowie Nervenzellen (= Neuronen) sind die Bausteine des Nervensystems. Beide Zellarten unterscheiden sich vorwiegend in ihren Aufgaben.
 sind die Bausteine des Nervensystems. Beide Zellarten unterscheiden sich vorwiegend in ihren Aufgaben.") (C) 2014 - SchulLV 1 von 5 Einleitung Du stehst auf dem Fußballfeld und dein Mitspieler spielt dir den Ball zu. Du beginnst loszurennen, denn du möchtest diesen Ball auf keinen Fall verpassen. Dann triffst
(C) 2014 - SchulLV 1 von 5 Einleitung Du stehst auf dem Fußballfeld und dein Mitspieler spielt dir den Ball zu. Du beginnst loszurennen, denn du möchtest diesen Ball auf keinen Fall verpassen. Dann triffst
beauty is our passion HAIR PLUS Ein Muss für jede Frau!
 beauty is our passion HAIR PLUS Ein Muss für jede Frau! Schönes, volles Haar macht jede Frau glücklich! Haarverlust ist für alle Frauen ein Albtraum, denn durch unsere Haarpracht fühlen wir uns selbstbewusst,
beauty is our passion HAIR PLUS Ein Muss für jede Frau! Schönes, volles Haar macht jede Frau glücklich! Haarverlust ist für alle Frauen ein Albtraum, denn durch unsere Haarpracht fühlen wir uns selbstbewusst,
Themenkomplex 1: Grundlagen der Anatomie von Herz und Kreislauf. Einstiegsfragen:
 Themenkomplex 1: Grundlagen der von Herz und Kreislauf 1. Beschreiben Sie den Weg des Blutes durch den Körper. 2. Erklären Sie den Unterschied von Arterien und Venen. Themenkomplex 2: Orientierung am Körper
Themenkomplex 1: Grundlagen der von Herz und Kreislauf 1. Beschreiben Sie den Weg des Blutes durch den Körper. 2. Erklären Sie den Unterschied von Arterien und Venen. Themenkomplex 2: Orientierung am Körper
Seite 1. Aus der Klinik für Gastroenterologie, Hepatologie und Endokrinologie der Medizinischen Fakultät der Charité Universitätsmedizin Berlin
 Seite 1 Aus der Klinik für Gastroenterologie, Hepatologie und Endokrinologie der Medizinischen Fakultät der Charité Universitätsmedizin Berlin DISSERTATION Immunhistochemische Charakterisierung von Wachstumsfaktoren
Seite 1 Aus der Klinik für Gastroenterologie, Hepatologie und Endokrinologie der Medizinischen Fakultät der Charité Universitätsmedizin Berlin DISSERTATION Immunhistochemische Charakterisierung von Wachstumsfaktoren
Sezieren eines Schweinefußes
 Anleitungen zu Modul 1 / NAWI-LAB Biologie am 13.10.2015 Sezieren eines Schweinefußes Materialien: Schweinefuß, Handschuhe, Küchenrolle, ( Skalpell+Klingen, Pinzette :wenn vorhanden), Fotoapparat; Ihr
Anleitungen zu Modul 1 / NAWI-LAB Biologie am 13.10.2015 Sezieren eines Schweinefußes Materialien: Schweinefuß, Handschuhe, Küchenrolle, ( Skalpell+Klingen, Pinzette :wenn vorhanden), Fotoapparat; Ihr
Haar und Kopfhaut und deren Pflege
 Haar und Kopfhaut und deren Pflege 1 Haararten Haar und Kopfhaut und deren Pflege 1.1 Erklären Sie die Begriffe a) Primärbehaarung, b) Sekundärbehaarung, c) Terminalbehaarung. 1.2 Nennen Sie die Haararten
Haar und Kopfhaut und deren Pflege 1 Haararten Haar und Kopfhaut und deren Pflege 1.1 Erklären Sie die Begriffe a) Primärbehaarung, b) Sekundärbehaarung, c) Terminalbehaarung. 1.2 Nennen Sie die Haararten
D D D Bestimmung des Zytokeratinprofils
 54 D D D D Abb. 17: Defekte Desmosomen, die zwei atypische Keratinozyten miteinander verbinden. Durch fehlende interzelluläre Zytokeratinverbindungen im Bereich der Desmosomen dense lines (D) erscheint
54 D D D D Abb. 17: Defekte Desmosomen, die zwei atypische Keratinozyten miteinander verbinden. Durch fehlende interzelluläre Zytokeratinverbindungen im Bereich der Desmosomen dense lines (D) erscheint
Schattauer Schattauer Schattauer. Schattauer. Schattauer. 6 Kreislaufsystem.
 6 Kreislaufsystem 2010 GmbH, Stuttgart Arterie univakuoläres Fettgewebe Adventitia (Tunica externa) Media (Tunica media) Lumen der Arterie mit Blut Intima (Tunica interna) Arterie, Gekröse, Pferd. Färbung
6 Kreislaufsystem 2010 GmbH, Stuttgart Arterie univakuoläres Fettgewebe Adventitia (Tunica externa) Media (Tunica media) Lumen der Arterie mit Blut Intima (Tunica interna) Arterie, Gekröse, Pferd. Färbung
MORPHOLOGISCHE VARIATIONEN DER WAAGRECHTEN WAND DES LÄNGSPARENCHYMS IM HOLZ VON TAXODIUM ASCENDENS BRONGN M. KEDVES
 161 MORPHOLOGISCHE VARIATIONEN DER WAAGRECHTEN WAND DES LÄNGSPARENCHYMS IM HOLZ VON TAXODIUM ASCENDENS BRONGN M. KEDVES Botanisches Institut der Universität, Szeged (Eingegangen: 15. April, 1959).. Einleitung
161 MORPHOLOGISCHE VARIATIONEN DER WAAGRECHTEN WAND DES LÄNGSPARENCHYMS IM HOLZ VON TAXODIUM ASCENDENS BRONGN M. KEDVES Botanisches Institut der Universität, Szeged (Eingegangen: 15. April, 1959).. Einleitung
Uber den Nasenhaarbalg bei einigen
 Uber den Nasenhaarbalg bei einigen Saugetieren Von Masanori Maeda Aus dem Anatomischen Institut der Keio Universitat in Tokyo. (Direktor: Prof. Dr. T. Taniguchi). Ich habe in der friiheren Mitteilung ither
Uber den Nasenhaarbalg bei einigen Saugetieren Von Masanori Maeda Aus dem Anatomischen Institut der Keio Universitat in Tokyo. (Direktor: Prof. Dr. T. Taniguchi). Ich habe in der friiheren Mitteilung ither
Charakteristisch für die Lymphfollikel der Milz sind die darin enthaltenen Arteriolen. Abb. 4 Pinselarteriolen Mensch, HE-Färbung
 Die ist das größte lymphatische Organ und in besonderer Weise in den Blutstrom integriert. Sie ist nicht in den Lymphstrom eingeschaltet und hat folgende Aufgaben: Reinigung des Blutes von körperfremden
Die ist das größte lymphatische Organ und in besonderer Weise in den Blutstrom integriert. Sie ist nicht in den Lymphstrom eingeschaltet und hat folgende Aufgaben: Reinigung des Blutes von körperfremden
Der Medizinkamm fürs Haar Ein Ratgeber bei Haarausfall für Sie und Ihn
 Der Medizinkamm fürs Haar Ein Ratgeber bei Haarausfall für Sie und Ihn Ein Wort zuvor: Etwas Haarverlust ist ganz natürlich Das große Problem: Erblich bedingter Haarausfall as Leben eines Haares dauert
Der Medizinkamm fürs Haar Ein Ratgeber bei Haarausfall für Sie und Ihn Ein Wort zuvor: Etwas Haarverlust ist ganz natürlich Das große Problem: Erblich bedingter Haarausfall as Leben eines Haares dauert
Dermatologie Wissenswertes über Haarausfall.
 Dermatologie Wissenswertes über Haarausfall. Wer einmal Originalmedikamente hergestellt hat, kann erst recht Generika in Originalqualität produzieren: Sandoz seit 1886. Aufbau der Haare 4 Haarzyklus 6
Dermatologie Wissenswertes über Haarausfall. Wer einmal Originalmedikamente hergestellt hat, kann erst recht Generika in Originalqualität produzieren: Sandoz seit 1886. Aufbau der Haare 4 Haarzyklus 6
Biologie. Was ist das? Was tut man da? Womit beschäftigt man sich?
 Biologie Was ist das? Was tut man da? Womit beschäftigt man sich? Wiederholung Merkmal des Lebens Aufbau aus Zellen Alle Lebewesen bestehen aus Zellen. Man kann grob drei verschiedene Zelltypen unterscheiden?
Biologie Was ist das? Was tut man da? Womit beschäftigt man sich? Wiederholung Merkmal des Lebens Aufbau aus Zellen Alle Lebewesen bestehen aus Zellen. Man kann grob drei verschiedene Zelltypen unterscheiden?