ECC. -- β-aktin GLUT1 GLUT2 GLUT3 GLUT4 GLUT8
|
|
|
- Teresa Fried
- vor 6 Jahren
- Abrufe
Transkript
1 3 Ergebnisse Im ersten Teil der vorliegenden Arbeit wurde der Einfluss von auf embryonale Stamm- und Karzinomzellen und die Expression der Glukosetransporter-Isoformen untersucht. Dabei wurden 2 Zelllinien verwendet, die D3-ESC und die P19-EC-Zellen. Die ES-Zellen sind fähig, in ektodermale, endodermale und mesodermale Zellen zu differenzieren. In der frühen Phase der spontanen Differenzierung der ESC als embryoid bodies sind die Differenzierungsprozesse mit der frühen Embryonalentwicklung vergleichbar. ES-Zellen brauchen für das Wachstum eine feeder layer Zellschicht. Um toxikologische Effekte auf die feeder layer Zellen und somit indirekte Effekte auf die ESC auszuschließen, wurden die toxikologischen Studien mit an den P19-ECC durchgeführt. P19-ECC können durch Zugabe von Mediensupplementen in myogene und neuronale Zelltypen differenziert werden. 3.1 Charakterisierung von D3-ESC und P19-ECC bezüglich der Expression von Glukosetransportern Um einen Überblick über die Ausstattung der beiden Zelllinien mit GLUT und deren Expression während der spontanen Differenzierung (D3) und kardiomyogenen Differenzierung (P19) zu erhalten, wurden die Transkripte von GLUT1, 2, 3, 4, 8 durch RT- PCR detektiert (vgl. Abb. 14). In undifferenzierten D3-ES-Zellen sind die Isoformen 1, 3 und 8 konstitutiv exprimiert. GLUT4 konnte nach 2 Tagen Kultur, GLUT2 erst ab einer 15 tägiger EB-Kultur nachgewiesen werden. Die GLUT4-Transkriptmenge nimmt während der EB- Differenzierung zu, während die Menge an GLUT3 in 20 Tage alten D3-EBs verringert war. In undifferenzierten P19-EC-Zellen und während der myogenen Differenzierung sind GLUT1, 3, 4, 8 konstitutiv exprimiert. Das Transkript für GLUT2 konnte nicht nachgewiesen werden. Beide Zelllinien zeigen ähnliche Expressionsmuster für GLUT1, 2, 3 und 8, nicht jedoch für GLUT4. A ES ESC D3 EB 2d 6d 9d 15d 20d M L -- β-aktin B ECC ECC P19 EB 2d 5d 5+5d -- β-aktin GLUT1 GLUT2 GLUT3 GLUT4 GLUT8 GLUT1 GLUT2 GLUT3 GLUT4 GLUT8 Abb. 14: RT-PCR-Analyse der GLUT-Expression in D3 (A) und P19 (B)-Zellen Die Expression der Glukosetransporter (GLUT) 1, 2, 3, 4 und 8 wurde in D3-ES-Zellen und EBs nach 2, 6, 9, 15, 20 Tagen (d) Kultur (A), in P19-EC-Zellen (ECC) und myogen differenzierten EBs bestimmt (B). Als Positivkontrollen wurden die cdna aus Muskel (M) bzw. Leber (L) mitgeführt. ES= embryonale Stammzellen, ECC= embryonale Karzinomzellen, d=kultivierungsdauer in Tagen, --=Wasserkontrolle 46
2 3.1.1 Detektion von GLUT4 im Western Blot 3 Ergebnisse Die Transkriptmenge von GLUT4 steigt während der EB-Differenzierung an. Diese Zunahme konnte auch auf der Proteinebene durch Western Blot-Analyse bestätigt werden. Eine spezifische GLUT4-Bande wurde bei 45kD detektiert. Die Bandenintensität nimmt von 2 Tagen Kultur bis 15 Tagen Kultur zu. In undifferenzierten D3-Zellen ist kein Protein nachweisbar. ES EB 2d 6d 9d 15d 20 M 45kD Abb. 15: Detektion von GLUT4 im Western Blot Die Proteinproben (30µg) von undifferenzierten D3-Zellen (ES) und 2, 6, 9, 15 und 20 Tage (d) kultivierten EBs wurden mit dem spezifischen monoklonalen Antikörper anti-glut4 (DPC Biermann) im Western Blot detektiert. Als Referenz wurde Muskelgewebe aus der Maus (M) aufgetragen. ES= embryonale Stammzellen, d= Kultivierungsdauer in Tagen, M= Muskelprotein Lokalisation der GLUT in D3-EBs mittels IHC Spontan differenzierende EBs wie auch myogen und neuronal differenzierte ES-Zellen wurden immunhistochemisch auf die Lokalisation von GLUT1, 2, 3 und 4 untersucht. Die myogen differenzierten EBs wurden zur Lokalisation des GLUT4, die neuronalen für den sog. brain type GLUT3 verwendet. Um zu bestimmen, welcher Zelltyp einen bestimmten Glukosetransporter exprimiert, wurden Doppelfärbungen mit spezifischen Zellmarkern für a) undifferenzierte Zellen mit dem Stage Specific Embryonic Antigen (SSEA-1), b) endodermale Zellen mit Zytokeratin 18, 19, c) myogene mit Desmin und d) neuronale Zellen mit Nestin durchgeführt GLUT1-Lokalisation in spontan differenzierten D3-EBs In EBs nach 2d-20d Kultur ist das GLUT1-Protein in der Membran der Zellen lokalisiert (vgl. Abb. 16). Ab Tag 15 und Tag 20 konnten Zellpopulationen nachgewiesen werden, die GLUT1-negativ sind. Gleichzeitig verändert sich in 20 Tage kultivierten EBs die membranständige in eine zytoplasmatische Lokalisation (vgl. Abb. 16). Die GLUT1 + -Zellen liegen innerhalb des EBs, eine Orientierung zur Oberfläche oder Polarisation in der GLUT1- Verteilung war nicht zu erkennen GLUT2-Lokalisation in spontan differenzierten D3-EBs Übereinstimmend mit dem Nachweis der Transkripte ist das GLUT2-Protein nur in 15d und 20d kultivierten EBs nachweisbar (vgl. Abb. 16). Dabei ist GLUT2 im Zytoplasma und in der Membran einiger weniger Zellen lokalisiert. In der Doppelfärbung von GLUT2 mit GLUT4 ergeben sich Zellpopulationen die sowohl GLUT2 als auch GLUT4 (GLUT2 + /GLUT4 + ) oder 47
. Die GLUT2 + -Zellen ordnen sich in Zellgruppen im Inneren der EBs an.")

 Kulturdauer.")

. Die Kerne sind blau mit Hoechst 33258 gegengefärbt.")

.")
.")
.")

.")

3 nur GLUT2- (GLUT2 + /GLUT4 -- ) oder GLUT4-Färbung (GLUT2 -- /GLUT4 + ) aufweisen (vgl. Abb. 21 C). Die GLUT2 + -Zellen ordnen sich in Zellgruppen im Inneren der EBs an. 2d 9d 15d 20d GLUT1 GLUT1 GLUT1 GLUT1 GLUT2 GLUT2 GLUT2 GLUT2 Abb. 16: Lokalisation von GLUT1- und GLUT2-Protein in spontan differenzierenden EBs Verteilung des GLUT1- und GLUT2-Proteins in spontan differenzierten EBs von 2 bis 20 Tagen (d) Kulturdauer. Die Lokalisation der GLUT Isoformen innerhalb der Zellen ist in einer höheren Vergrößerung in der rechten Ecke dargestellt. GLUT1 ist in 2d EBs nur membranständig ( ), in 20d kultivierten EBs membranständig und zytoplasmatisch lokalisiert (). Die Kerne sind blau mit Hoechst gegengefärbt. Maßstab 50µm, in den kleineren Ausschnitten 20µm GLUT3-Lokalisation in spontan differenzierten D3-EBs Der Glukosetransporter 3 ist in der Zellmembran von ES- und EB-Zellen lokalisiert (vgl. Abb. 17 A-D). Während der EB-Kultur kommt es zur Abnahme der GLUT3 + -Zellen. In 20 Tage alten EBs konnte kein GLUT3-Protein mehr detektiert werden (vgl. Abb. 17 D). Der Mangel an GLUT3-Protein in 20d EBs korreliert mit der geringen Transkriptmenge in der RT-PCR-Analyse (vgl. Abb. 14). Die GLUT3 + -Zellen liegen im EB 2d im äußeren Bereich, bei 9d sogar auffällig in den äußeren 2-3 Zelllagen des EBs. Die Randzellen zeigen eine starke, apikale (zur Oberfläche des EBs gerichtete) GLUT3-Markierung (vgl. Abb. 17 A, 19 K). Die starke Färbung von GLUT3 in der apikalen Membran deutet auf eine Funktion dieses Transporters für den Glukoseinflux in den EB durch die äußere Zellmembran hin (vgl. Abb. 17 B). Um den GLUT3 + Zelltyp näher zu charakterisieren, wurden Kolokalisationen mit SSEA-1, Zytokeratin 18 (CK18), Zytokeratin 19 (CK19), Desmin und Nestin in spontan und neuronal differenzierten EBs durchgeführt (vgl. Abb. 19). GLUT3 ist hauptsächlich in undifferenzierten inneren Zellen der EBs (SSEA-1 + /GLUT3 + ) und in der äußeren Zellschicht früher EBs exprimiert. Die Randzellen im 9d EB werden als endodermal-like cells (ELC) bezeichnet und sind durch SSEA-1 --, CK18 +, CK19 + markiert. 48
")

 lassen sich die ELC als GLUT3 + nachweisen.")


 Nestin nicht detektierbar war, konnten in")
. In 5+3d alten EBs waren alle GLUT3 positiven Zellen auch Nestin positiv.")
 (vgl. Abb. 19 B, J-L).")
. 3.1.2.")

4 A 2d B 9d C 15d D 20d Abb. 17: GLUT3-Proteinlokalisation in spontan differenzierenden EBs Lokalisation von GLUT3 (grün) in Schnitten von spontan differenzierten EBs nach 2 bis 20 Tagen (d) Kultur. Die Kerne sind mit Hoechst blau gegengefärbt. Maßstab 20µm. Durch Doppelmarkierung von GLUT3 mit SSEA1 (vgl. Abb. 19 A-C), CK18 (vgl. Abb. 19 D-F) und mit CK19 (vgl. Abb. 19 G-I) lassen sich die ELC als GLUT3 + nachweisen. GLUT3 ist demnach einerseits mit der Entwicklung von ELC assoziiert. Weiterhin findet man vor allem in der Mitte der EBs auch GLUT3 + -Zellen mit SSEA-1 --, CK18 +, CK Markierung. Da GLUT3 in adulten Zellen der spezifische Transporter im neuronalen Gewebe ist, wurde auch Nestin, ein Protein von neuronalen Vorläuferzellen, in die Studie miteinbezogen. Während in spontan differenzierten EBs (2-20d) Nestin nicht detektierbar war, konnten in neuronal differenzierten EBs (5+3d) Nestin und GLUT3 kolokalisiert werden (vgl. Abb. 19 B, M-O). In 5+3d alten EBs waren alle GLUT3 positiven Zellen auch Nestin positiv. Wie Nestin war auch das filamentöse Muskelprotein Desmin nicht in spontan differenzierten EBs detektierbar (2d-20d) (vgl. Abb. 19 B, J-L). Als Antikörperkontrolle wurde die IHC auch auf Gewebeschnitten vom Intestinum durchgeführt (vgl. Abb. 19 B, L) GLUT4-Lokalisation in spontan differenzierten D3-EBs GLUT4 ist in 2-20d EBs mit zunehmender Kulturdauer nachweisbar (vgl. Abb. 18 A-D). Das Protein ist in EBs nach 2d Kultur (vgl. Abb. 18 A) hauptsächlich in den äußeren Zellen lokalisiert, während an Tag 9-20 ein Großteil der Zellen GLUT4-positiv ist. Das GLUT4- Protein befindet sich hauptsächlich im Zellkern (vgl. Abb. 20 D-F). A 2d B 9d C 15d D 20d Abb. 18: GLUT4-Protein in spontan differenzierten EBs Lokalisation des GLUT4-Proteins (rot) in EBs nach 2 bis 20 Tagen (d) Kultur. Die Kerne sind mit Hoechst blau gefärbt. Maßstab 20µm. 49






















5 3 Ergebnisse B A C SSEA-1 SSEA-1/GLUT3 D SSEA-1/GLUT3 SSEA-1/GLUT3 F E CK18 CK18/GLUT3 G CK18/GLUT3 CK18/GLUT3 I H CK19 CK19/GLUT3 CK19/GLUT3 K L CK19/GLUT3 J Desmin Desmin/GLUT3 Desmin/GLUT3 M N Desmin O Nestin Nestin/GLUT3 Nestin/GLUT3 Nestin/GLUT3 Abb. 19: Kolokalisation des GLUT3-Protein mit zellspezifischen Markern Doppelfärbung von GLUT3 mit verschiedenen Zellmarkern. A-C: GLUT3 und SSEA-1 in 6d EBs D-F: GLUT3 und CK18 in 9d EBs G-I: GLUT3 und CK19 in 9d EBs, J-L: GLUT3 und Desmin in 9d EBs und L Maus Jejunum als Positivkontrolle für Desmin L M-O: GLUT3 und Nestin in 9d EBs M, N und 3+5d neuronal differenzierte Zellen O. Negativkontrollen sind als kleinere Bilder in der rechten Ecke dargestellt. Maßstab 50µm. 50



 und")
 EBs.")

.")
, d.h.")

, wo GLUT4 in der apikalen Membran der äußeren")
6 A B C 6d GLUT4 SSEA-1 GLUT4/SSEA-1 D E F 9d GLUT4 20 HOECHST GLUT4/HOECHST G H I 20d GLUT4 HOECHST GLUT4/HOECHST J K L 5+5d plated EB GLUT4 Desmin GLUT4/Desmin 20µm Abb. 20: Zellspezifische Lokalisation von GLUT4 und Kolokalisation mit zelltypspezifischen Markern in spontan differenzierten und plattierten EBs Doppelfärbung von GLUT4 (rote Fluoreszenz) mit SSEA-1 (braun) (A-C). Zelluläre Lokalisation von GLUT4 im Kern von 9d (D-F) und 20d (G-I) EBs, Membranlokalisation in 20d (G-I) und 5+5d plattierten (J-L) EBs. Die Negativkontrollen sind in der rechten Ecke dargestellt. Maßstab 20µm. Durch Kolokalisation mit SSEA-1 und Desmin wurde die zelluläre Lokalisation näher untersucht (vgl. Abb. 20). SSEA-1 + -Zellen sind nicht GLUT4 positiv (vgl. Abb. 20 A-C), d.h. GLUT4 ist nicht in undifferenzierten Zellen vorhanden. Die erste Membranlokalisation von GLUT4 ist in späten EBs (20d) nachweisbar (vgl. Abb. 20 G-I), wo GLUT4 in der apikalen Membran der äußeren EB-Zellen (zum Medium gerichtet) lokalisiert ist. Um die ungewöhnliche nukleäre Lage des GLUT4 (vgl. Abb. 20 D-F) zu verifizieren, wurden zusätzlich plattierte EBs (5+5d) untersucht, die myogene Zelltypen enthielten. GLUT4 wurde 51
 war GLUT4 nachweisbar, wobei die zelluläre Lage auf die Zellmembran und das Zytoplasma beschränkt war")

, befindet sich dort im Zytoplasma und der Membran von innenliegenden Zellklustern (vgl. Abb. 21 D).")

7 in diesen Zellen nach gleichem Protokoll detektiert und mit dem myogenen Marker Desmin kolokalisiert. Auch in den myogen-differenzierten EB-Zellen (Desmin + /GLUT4 + ) war GLUT4 nachweisbar, wobei die zelluläre Lage auf die Zellmembran und das Zytoplasma beschränkt war (vgl. Abb. 20 J-L). Die Expression von GLUT4 wurde mit GLUT1 und GLUT2 im 20d kultivierten EB kolokalisiert (vgl. Abb. 21). In den äußeren Zellen des EBs, in denen sich GLUT4 in der Zellmembran befindet, liegt GLUT1 im Zytoplasma (vgl. Abb. 21 A, B), wobei in den inneren Zellen GLUT4 weiterhin im Zellkern liegt. Die Transporterisoform 2, die erst in 20 Tage alten EBs nachweisbar wird (vgl. Abb. 21 C), befindet sich dort im Zytoplasma und der Membran von innenliegenden Zellklustern (vgl. Abb. 21 D). Die GLUT2 positiven Zellen können als GLUT2 + /GLUT4 + und GLUT2 + /GLUT4 Zellen bestimmt werden. GLUT4 ist in GLUT2 + Zellen im Kern lokalisiert. A B 20d GLUT1/GLUT4/HOECHST GLUT1/GLUT4/HOECHST C D 20d GLUT2/GLUT4/HOECHST GLUT2/GLUT4/HOECHST Abb. 21: GLUT4 Kolokalisation mit GLUT1 und GLUT2 im 20d kultivierten EB Kolokalisation von GLUT4 (rot) mit GLUT1 (grün) (A; B) und GLUT2 (grün) (C-D). In A und C ist die Übersichtsaufnahme eines 20 Tage kultivierten EBs, in B und D ein vergrößerter Ausschnitt dargestellt. Die Kerne sind mit Hoechst blau gefärbt (A, B, C, G). Maßstab 20µm. 52
 mit 1g/l angesetzt.")
.")


.")


. In der Immunhistochemie ist die Lokalisation von SSEA-1 in der Zellmembran der EBs dargestellt (braune Färbung).")
8 3.1.3 Einfluss der Glukosekonzentration auf die EB-Differenzierung 3 Ergebnisse Die Standard-Glukosekonzentration im Kulturmedium von EBs beträgt 4,5g/l. Um den Einfluss von Glukosemangel auf die Expression von GLUT zu untersuchen, wurde eine Minimalkultur (Glc -- ) mit 1g/l angesetzt. Bereits ein Vergleich der Morphologie der EBs bei einem Glukosegehalt von 1g/l im Medium zeigt deutliche Veränderungen gegenüber der normalen Struktur (vgl. Abb. 22). Da EBs eine Vielzahl verschiedener GLUT-Isoformen exprimieren stellte sich die Frage, welche GLUT durch Glukosemangel reguliert bzw. in der Expression verändert werden. Hierzu wurden die EBs für 4d, 6d, 8d in Glc -- -Medium kultiviert. Anschließend wurden die Transkriptmengen der GLUT-Gene und Oct-4 bestimmt sowie GLUT1, GLUT3 und GLUT4 mittels Immunfluoreszenz lokalisiert. Als Vergleich wurden immer die unter Normalbedingungen kultivierten EBs herangezogen Morphologie der EBs und SSEA-1-Präsentation unter Glc-Mangel Durch IHC wurde die Morphologie und der Anteil undifferenzierter SSEA-1 positiver Zellen in den unter Glc-Mangel kultivierten EBs mit der von Standard-EBs verglichen (vgl. Abb. 22). Die unter Glukosemangel kultivierten EBs waren insgesamt kleiner und bildeten artifizielle Höhlen, sog. cavities (vgl. Abb. 22 D, F). Der Gehalt an undifferenzierten SSEA Zellen war in EBs unter Glukosemangel deutlich erhöht (vgl. Abb. 22 E, B), was darauf hinweist, dass Glukosemangel ein verzögertes Wachstum und eine spätere Differenzierung zur Folge hat. A 4d 6d 8d C Standard Glc D F Niedrig Glc Abb. 22: Morphologie von EBs unter Glukosemangel und Expression von SSEA-1 A-C: EBs kultiviert in Standardglukose-Medium (25mM); D-F: mit Glukosemangelmedium (5mM). In der Immunhistochemie ist die Lokalisation von SSEA-1 in der Zellmembran der EBs dargestellt (braune Färbung). Die Kerne wurden mit Hämalaun blau gegengefärbt. Maßstab 30µm Transkription von Oct-4 unter Glukosemangel Oct-4 ist ein Transkriptionsfaktor, der in undifferenzierten embryonalen Stammzellen der Maus hoch exprimiert ist. Während der Differenzierung nimmt die Oct-4-Menge ab. Oct-4 und SSEA-1 sind Marker für undifferenzierte Zellen während der EB-Differenzierung. In EBs kultiviert unter Standard- und Glc-Mangelbedingungen wurde deshalb die Transkriptmenge 53
9 von Oct-4 mittels Real-time PCR quantifiziert (vgl. Abb. 23). In EBs kultiviert für 4 Tage unter Glukosemangel war die Oct-4 Transkriptmenge weniger verringert als in Standard-EBs (vgl. Abb. 23). In 6d und 8d kultivierten EBs ist keine signifikante Änderung in der Oct-4 Expression nachweisbar, was durch die niedrige Expression des Oct-4 in späteren Stadien der EBs bedingt sein könnte. 35 Oct-4 mrna-menge [% zur ES-Zellkontrolle] ,5g Standard-Glc 1g Glukosemangel Kultivierung in Tagen Abb. 23: Quantifizierung des Oct-4 mrna-gehaltes in EBs unter Standard-Glc (4,5g) und Glukosemangel (1g) Die mrna-expression von Oct-4 wurde mittels Real-time PCR quantifiziert (vgl. 2.11). Die Daten wurden relativ zu undifferenzierten ESC D3 berechnet. N=3, n=4-6, p<0, Expression von GLUT mrna und Protein in EB kultiviert unter Glukosemangel Die mrna-expression für GLUT 1, 3 und 4 wurde mittels kompetitiver PCR, für GLUT8 mittels semiquantitativer PCR bestimmt (vgl. Tab. 7; Abb. 24 A). Nach 4d EB- Differenzierung unter Glukosemangel kam es zu einer generellen Minimierung der Expression aller untersuchten GLUT-mRNA. Nach 6 und 8 Tagen wurden die Glukosetransporter 3 und 4 vermehrt exprimiert verglichen mit unter Standardbedingungen (25mM) kultivierten EBs. GLUT1 und GLUT8 zeigten keine signifikanten Änderungen in der mrna-menge. Die GLUT1-Proteinmenge und seine subzelluläre Lokalisation wird nicht durch Glukosemangel beeinflusst (vgl. Abb. 24 B). 54
10 Tab. 7: mrna Quantifizierung für GLUT1, 3, 4 und 8 unter Glukosemangel Relative RNA Mengen [% der Kontrolle] 4d 6d 8d GLUT1 47,53+9,26 99,45+1,18 126,84+10,69 2 GLUT3 74,98+30,22 187,25+23,18 172,13+52,69 GLUT4 44,96+5,09 193,725+37,13 355,16+42,88 2 GLUT8 50,86+8,75 93,51+28,57 105,38+52,69 N=2, n=2-3; p 0,001, 2 p 0,05 Signifikanz zu den EBs kultiviert mit 25mM Glc GLUT3 wurde mittels Immunfluoreszenz in der Zellmembran der inneren und äußeren Zellen des EBs lokalisiert (vgl. Abb. 24 B). Unter Glukosemangel kam es zu einem geringen Anstieg der Expression in den inneren EB-Zellen, was mit dem Anstieg der GLUT3-Transkriptmenge korreliert. Glukosemangel hat keinen sichtbaren Einfluss auf die Verteilung von GLUT3 in den EBs und auf seine membranständige Lokalisation. GLUT4 war in den untersuchten EB-Stadien hauptsächlich in den äußeren Zellen nachweisbar. In EBs unter Glukosemangel war das GLUT4-Fluoreszenzsignal deutlich stärker als in der Kontrolle (vgl. Abb. 24 B). Der Anstieg der Immunfluoreszenz korreliert mit dem Transkriptionsanstieg. Das zeigt, dass es unter Glukosemangel in EBs zu einer erhöhten Expression von GLUT4 vor allem in den äußeren Zellschichten kommt. 55











")


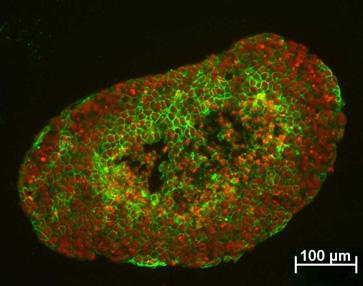




11 A B + relative mrna-menge [%] d 6d 8d 4d 6d 8d A B C GLUT1 GLUT3 GLUT4 GLUT8 GLUT1 - D E F GLUT3 + A B C D E F - A B C + GLUT4 D E F - Abb. 24: Einfluss von Glukosemangel auf die mrna- und Protein-Expression von GLUT1, 3 und 4 A Die Quantifizierung von GLUT1, 3 und 4 wurde mittels crt-pcr durchgeführt. EBs wurden über die gleiche Zeit in normalem (25mM) Glukosemedium kultiviert und als Kontrollen (100%) benutzt. N=2 n=2 Signifikanz: p<0.001; p<0.05. B A-C: EBs kultiviert in Standardglukose-Medium (25mM) (+); D-F: mit Glukosemangel-Medium (5mM) (-). A,D: 4d EB; B,E: 6d EB; C,F: 8d EB. Immunfluoreszenz mit anti-glut1 (grün), anti-glut3 (grün) und anti-glut4 (rot). Die Kerne sind bei GLUT1 und GLUT4 blau durch Hoechst-Färbung, bei GLUT3 rot durch 7-AAD-Färbung. Maßstab 50µm. 56
12 3.2 Einfluss von auf P19-ECC Wirkung von auf die Zellproliferation Die Einwirkung von auf die Zellproliferation von P19-ECC wurde nach dem mitochondrialen Toxizitätstest (MTT) bestimmt (vgl. 2.6, Abb. 25). Unbehandelte P19-Zellen haben eine Proliferationsrate von 100%. Abhängig von der -Konzentration ist bei hohen Konzentrationen (10-100nM) die Proliferation um ca. 10%, bei 1nM um 5% verringert. Im Gegensatz dazu nimmt die Proliferation der nur mit DMSO-behandelten Kontrollzellen um ca. 30%, abhängig von der zugeführten DMSO-Menge, zu. Bei DMSO Konzentration von 6,25x10-4 bzw. 6,25x10-3 %ig (entsprechend der Kontrollmenge bei 10nM und 100nM ) erhöht sich die Proliferation um 24% bzw. 31%. In Bezug auf die DMSO-Kontrollzellen wirkt proliferationshemmend (ca. 30% bei 10nM, 40% bei 100nM). Gegenüber unbehandelten P19-ECC ist die Abnahme durch bei der Proliferation nicht signifikant, wobei DMSO allein die Proliferation signifikant erhöht (vgl. Abb. 25). Zellproliferation [% der unbehandelten Kontrolle] ,1 nm 1 nm 10 nm 100 nm Konzentration 2 2 DMSO Abb. 25: Zellproliferation von P19-ECC unter Zugabe von P19-ECC wurden mit 0,1nM bis 100nM in DMSO und den entsprechenden DMSO-Kontrollen kultiviert und die Zellproliferation bestimmt (vgl. 2.6). Die Zellproliferation unbehandelter P19- Kontrollzellen wurden 100% gesetzt. n=8; p 0,05 Signifikanz zu P19; 2 p 0,001 Signifikanz zu DMSO Einfluss von auf die Apoptose von P19-ECC Der Einfluss von auf die Apoptose der P19-ECC wurde im Kaspase-3/7-Assay untersucht (vgl. 2.5.). Die Aktivität der Kaspasen wurde nach Zugabe von 0,1-100nM im Zellysat bestimmt, wobei die Kaspaseaktivität unbehandelter P19-ECC als 100% gesetzt wurde (vgl. Abb. 26). Bei niedrigen Konzentrationen von 0,1 und 1nM bewirkt eine Erhöhung der Kaspaseaktivität um 138% bzw. 124%. Mit hoher -Konzentration (10, 100nM) verringert sich die Aktivität um 29% (10nM ). Die Aktivität der Kaspasen 3 und 7 wird als Maß für die Apotoserate bewertet. Die Auswertung der Aktivitätsbestimmung zeigt, dass durch die Apoptose konzentrationsabhängig beeinflusst wird, während 57
13 DMSO allein keinen Einfluss hat. Bei geringen -Konzentration (0,1nM) kommt es zum Anstieg der Apoptoserate, während es unter höheren Konzentration (10-100nM) zu einer Verringerung der Apoptose kommt. Die Aktivität der Kaspasen der DMSO-Kontrollen ist nicht signifikant um ca % erhöht. wirkt auf P19-ECC zytotoxisch und beeinflusst die Apoptoserate. Kaspase 3/7-Aktivität [% zur unbehandeltent Kontrolle DMSO 0,1 nm 1 nm 10 nm 100 nm -Konzentration Abb. 26: Bestimmung der Kaspaseaktivität in P19-ECC P19-ECC wurden mit 0,1nM bis 100nM in DMSO und den entsprechenden DMSO-Kontrollen kultiviert und die Zellproliferation bestimmt (vgl. 2.5). Die Zellproliferation unbehandelter P19- Kontrollzellen wurden 100% gesetzt. N=1, n=3; p 0,05, 2 p 0,001 Signifikanz zu P19; p 0,05, 2 p 0,001 Signifikanz zu DMSO Wirkt in P19-ECC über die AhR-Signalkaskade? Um zu klären, ob die Wirkung von über den AhR in P19-Zellen vermittelt wird, wurde die AhR-Signalkaskade (vgl. Abb. 27) genauer analysiert. Dafür wurde untersucht: a) die mrna-expression des CYP1A1-Gens konzentrations- und zeitabhängig (vgl ) b) die mrna-expression des CYP1A1-Gens unter Inhibitorzugabe (vgl ) c) die Translokation des zytoplasmatischen AhR in den Kern (vgl ) d) die CYP1A1-Induktion durch die Aktivierung der XRE-Promotorregion (vgl ) Charakterisierung von D3-ESC und P19-ECC auf die Expression von AhR-Zielgenen Um einen Überblick über die Ausstattung der beiden Zelllinien mit AhR-Zielgenen und deren Expression während der spontanen Differenzierung (D3) und kardiomyogenen Differenzierung (P19) zu erhalten, wurden die Transkripte von AhR, ARNT, CYP1A1 und CYP1B1 durch RT-PCR detektiert. Alle Zielgene sind in beiden Zellinien konstitutiv exprimiert. Es gibt keine Unterschiede in D3- und P19-Zellen. Die CYP1B1-mRNA-Menge nimmt während der myogenen Differenzierung zu. Beide Zelllinien zeigen ähnliche Expressionsmuster. 58
14 AhR-Ligand - α NF AhR ARNT P + AhR Hsp90 Hsp90 + AhR Hsp90 Hsp90 AhR ARNT + P CYP1A1 Abb. 27: Schema der AhR-Signalkaskade ES ESC D3 EB 2d 6d 9d 15d 20d ECC P19 ECC EB -- 2d 5d 5+5d -- AhR AhR ARNT ARNT CYP1A1 CYP1B1 Abb. 28: RT-PCR-Analyse der AhR-Signalmoleküle in D3- und P19-Zellen Die Expression von AhR, ARNT, CYP1A1 und CYP1B1 wurde in D3-ES-Zellen und EBs nach 2, 6, 9, 15, 20 Tagen (d) Kultur und in P19-EC-Zellen (ECC) und kardiomyogener Differenzierung für 2, 5, 5+5 Tagen (d) bestimmt. ES= embryonale Stammzellen, ECC= embryonale Karzinomzellen, d=kultivierungsdauer in Tagen, --=Wasserkontrolle Konzentrations- und zeitabhängige Expression von CYP1A1 in P19-EC-Zellen unter -Zugabe Die Expression des AhR-Zielgens CYP1A1 wurde mittels Real-time PCR untersucht (vgl. Abb. 29). erhöht bei 1nM, 10nM und 100nM die CYP1A1 mrna-menge signifikant. Die Expression steigt bei 100nM um ca. das 85fache an (vgl. Abb. 29). Bereits ab 1nM erhöht sich die Transkriptmenge signifikant. CYP1A1-mRNA wird nach 2h Kultur mit konzentrationsabhängig induziert. Die Konzentration von 10nM wurde für die 59
15 nachfolgenden Experimente als Konzentrationsoptimum gewählt. 10nM ist eine übliche Konzentration, die auch in Studien anderer Zelllinien eingesetzt wird. CYP1A1 mrna-menge [rel. E.] P19 DMSO 0,1nM 1 nm 10 nm 100 nm Abb. 29: Relative mrna-menge von CYP1A1 in Abhängigkeit von der -Konzentration Durch Real-time PCR (vgl. 2.11) wurde die relative CYP1A1-mRNA-Menge nach 2 Stunden Inkubation mit 0,1; 1; 10 und 100nM quantifiziert. N=3 n=2; p 0,05 Signifikanz zu P19 Das Expressionsprofil der CYP1A1-Induktion durch wurde anhand einer Zeitreihe von 1, 2, 3, 4, 5, 6 und 24h erstellt. Die CYP1A1-mRNA-Menge wurde mittels Real-time PCR in P19-EC-Zellen quantifiziert (vgl. 2.11). Die CYP1A1-Menge der behandelten Zellen wurde relativ zum CYP1A1-Gehalt unbehandelter P19-ECC (=1 rel. E.) berechnet (vgl. Abb. 30). Sie erhöht sich nach 10nM für 1 bis 3 Stunden mit einem Maximum (ca. 21,1facher Erhöhung) nach 2h. Der Anstieg der CYP1A1-Transkriptmenge durch verläuft damit sehr schnell. Nach 4h fällt die Transkriptmenge unter die der P19-Kontrolle ab und liegt somit unterhalb der konstitutiven CYP1A1-Expression der P19-Zellen. Ab 24h steigt die CYP1A1- Menge auf 1,55 rel. E. an. Die Induktion der CYP1A1-Transkriptmenge in P19-ECC ist abhängig von der Einwirkzeit und Konzentration des CYP1A1 mrna-menge [rel. E.] DMSO Zeit in Stunden [h] Abb. 30: Relative CYP1A1-Transkriptmenge nach 1 bis 24h in Kultur mit 10nM in P19-ECC Durch quantitative PCR (vgl. 2.11) wurde die CYP1A1 Menge unter Gabe von 10nM nach 1-24h Kulturdauer in P19-ECC bestimmt. N=3 n=2; p 0,05, 2 p 0,001 Signifikanz zu P19-ECC 60
16 Inhibierung des AhR mittels α-naphthoflavon α-naphthoflavon inhibiert die Translokation des AhR in den Kern und unterbindet dadurch die AhR-vermittelte Aktivierung der CYP1A1 mrna-synthese (vgl. Abb. 27). Durch Zugabe von α-naphthoflavon zu den P19-Zellen vor der Exposition mit und der anschließenden Bestimmung der CYP1A1-Expression kann bestimmt werden, ob die - Wirkung über den AhR-Signalweg erfolgt oder davon unabhängig ist (vgl. Abb. 27). 2h vor der Inkubation mit (10nM) wurde α-nf in Konzentrationen von 0,1-100µM in das Zellkulturmedium gegeben. Nach weiteren 2h wurde die mrna-menge von CYP1A1 mittels Real-time PCR bestimmt. Mit steigenden α-nf-konzentrationen kommt es zu einer Abnahme der CYP1A1-Transkriptmenge in -behandelten P19-ECC. Die Induktion der CYP1A1-mRNA-Menge durch -Wirkung wird durch Zugabe von 0,1µM α-nf auf ca. die Hälfte reduziert (vgl. Abb. 31). Die Zugabe von µM α-nf inhibiert die CYP1A1- Induktion vollständig. Durch den Inhibitor α-nf wird die Induktion von CYP1A1 durch konzentrationsabhängig inhibiert, was mit einer kompetitiven Bindung von α-nf und Blockierung der -Bindung erklärbar ist. CYP1A1 mrna-menge [rel. E.] P19 DMSO 10nM 0,1µM a-nf 1µM a-nf 10µM a-nf 100µM a-nf + 10nM Abb. 31: Quantifizierung der mrna-menge von CYP1A1 nach Inhibierung des AhR mit α-nf Die CYP1A1-Transkriptmenge wurde mittels Real-time PCR (vgl. 2.11) bestimmt. P19-ECC wurden mit dem Inhibitor α-naphthoflavon (α-nf, 0,1µM-100µM) behandelt und anschließend mit 10nM exponiert. N=3 n=2; p 0,05, 2 p 0,001 Signifikanz zu P19; p 0,05, 2 p 0,001 Signifikanz zu DMSO Translokation des AhR nach -Wirkung Nach Aktivierung des AhR durch Bindung des Liganden transloziert der Rezeptor vom Zytoplasma (inaktiv) in den Kern (aktiv) (vgl. Abb. 27). Um diese Wanderung des Rezeptors untersuchen zu können wurde ein enhanced green fluorescence protein (EGFP)-AhR Fusionsprotein konstruiert (vgl. 2.7), was die Detektion des AhR anhand der grünen Fluoreszenz (488nm) des Reporterproteins ermöglicht. Die vollständige CDS wurde am C-Terminus mit EGFP in den Vektor pegfp und in P19-ECC 61



 befindet sich das Fluoreszenzsignal")






 fusioniert")
17 transfektiert (vgl. 2.7; 2.4). Nach 4h wurden die transient transfizierten P19-Zellen mit 10nM bzw. mit DMSO inkubiert und nach 45min mit 4%PFA fixiert. Die Auswertung erfolgte mittels Konfokaler Laser Scanning Mikroskopie (KLSM) (vgl. Abb. 32). Bei Versuchsbeginn lag der AhR hauptsächlich im Zytoplasma der P19-Zellen und es war nur ein schwaches Fluoreszenzsignal im Kern detektierbar. Nach 40minütiger Inkubation mit (10nM) befindet sich das Fluoreszenzsignal im Zellkern. Dies wird vor allem in der Doppelmarkierung deutlich, wo der grün markierte AhR mit der roten Kernfärbung übereinanderliegt (vgl. Abb. 32). Die resultierende gelbe Mischfarbe zeigt die Kolokalisation von EGFP-AhR und der Kerne an. Nur EGFP allein lag dagegen in der gesamten Zelle verteilt vor, im Zytoplasma wie auch im Kern, und zeigte keine Veränderung in der Lokalisation nach -Zugabe. Dieser Versuch zeigt, dass der AhR in P19-ECC durch innerhalb von 40 min vom Zytoplasma in den Kern wandert und somit aktiviert wird. EGFP-AhR 7-AAD Gesamt 0h 40 min DMSO Kontrolle P P P 40 min EGFP- Vektor Kontrolle + Abb. 32: Detektion der AhR-Translokation nach -Wirkung 62 Der AhR wurde durch Klonierung mit dem enhanced green fluorescence protein (EGFP) fusioniert und in P19-ECC transient transfektiert. Das Fluoreszenzsignal (grün) wurde bei 488nm im KLSM detektiert. In den linken Bildern ist nur das EGFP-Signal in grün, in der mittleren Bildreihe die Kerngegenfärbung mit 7-AAD in rot und in den rechten der Overlay der beiden Abbildungen dargestellt. EGFP als Vektorkontrolle ist überall in der Zelle verteilt und zeigt keine Lokalisationsveränderung nach -Zugabe. Maßstab 16µm.
18 Untersuchung der Bindung des AhR/ARNT-Transkriptinsfaktorkomplexes an das XRE-Element Im Kern dimerisiert der AhR mit ARNT. Nur dieser Heterodimerkomplex ist in der Lage, an die spezifische XRE-Sequenzen zu binden. Der AhR/ARNT-Transkriptionsfaktorkomplex aktiviert die Transkription von Zielgenen mit XRE-Erkennungssequenzen in der Promotorregion (vgl. Abb. 27). Um die Induktion der Zielgene durch die spezifische Bindung an das XRE untersuchen zu können, wurde ein XRE-Luziferase-Reportervektor konstruiert (vgl. 2.8) und transient in die P19-Zellen transfiziert. Die Bindung der XRE wurde über den Substratumsatz des Reporterenzyms firefly luciferase nachgewiesen. Als Kontrolle der Transfektionseffizienz und des Zellwachstums wurde ein Vektor mit konstitutiv exprimierender renilla luciferase kotransfiziert. Die Aktivität der firefly luciferase wurde im Vergleich zur renilla luciferase ausgewertet (vgl. Abb. 33). Die Luziferaseaktivität ist nach -Zugabe signifikant um 117,06% erhöht. Nur DMSO allein führt auch zu einer Steigerung der Luziferaseaktivität um ca. 15%. Die Steigerung der Luzifreaseaktivität in -behandelten Zellen ist ein Marker für eine XRE-Aktivierung durch den AhR Luziferaseaktivität [%] P19-ECC DMSO 10nM Abb. 33: Luziferaseaktivität von stimulierten P19-ECC In der Grafik ist die relative Luziferaseaktivität der firefly luciferase und renilla luciferase von unbehandelten P19-ECC, -stimulierten P19-ECC (10nM ) und der DMSO-Kontrolle (DMSO) gezeigt. Die Aktivität in den transfizierten P19-ECC wurde mit 100% Aktivität als Kontrolle berechnet. Die Aktivität der DMSO- und -Gruppen wurde in Relation zu den P19-ECC berechnet (firefly luciferase Aktivitäten/ renilla luciferase Aktivität). N=2 n=3 p 0,05, 2 p 0,001 Signifikanz zu P19; p 0,05 Signifikanz zu DMSO 63
19 Wirkung auf die myogene Differenzierung der P19-ECC P19-ECC können nach einem etablierten Zellkulturprotokoll in myogene Zellen differenziert werden. An diesem in vitro Differenzierungsmodell soll untersucht werden, ob einen Einfluss auf Entwicklungs- und Differenzierungsprozesse hat. Bei den P19 Zellen wurde durch 1%DMSO die kardiomyogene Differenzierung induziert, wobei nicht alle Zellen in Kardiomyozyten differenzieren. In der Kultur entwickeln sich zu einem geringeren Anteil auch Skelettmuskelzellen und neuronale Zellen. Der Einfluss von auf die Differenzierung von P19-Zellen wurde durch a) mikroskopische Analyse und Auszählung schlagender Zentren (vgl ) b) Expression des induzierbarem CYP1A1 (vgl ) c) Expression der muskelspezifischen Gene α-mhc, MyoD (vgl ) d) Expression von Mash-1 als Marker der neuronalen Differenzierung (vgl ) e) Expression von Glukosetransportern (vgl ) untersucht Lichtmikroskopische Untersuchung der Bildung kardiomyogener Zentren in 64 plattierten EBs Eine 2-tägige Exposition mit 10nM während der Formierung der EBs (0-2d) beeinflusst die anschließende Differenzierung der EBs zu Kardiomyozyten nicht. Es konnten keine signifikanten Unterschiede zwischen mit kultivierten EBs und den Kontrollen festgestellt werden (vgl. Abb. 34 A). Um einen DMSO-Effekt (Inducer der kardiomyogenen Differenzierung) ausschließen zu können, wurde der Versuch ohne die 1%ige DMSO-Gabe (Standardprotokoll) wiederholt. Dazu wurden die EBs während der ersten 2 Tage der Differenzierung nur mit 10nM in DMSO (6,25x10-4 %ig) inkubiert. Die Zellen formieren auch bei einer niedrigeren Konzentration des DMSO schlagende Herzmuskelzellen. Die Anzahl der Zentren beträgt bei 6,25x10-4 %igem DMSO nur 15% gegenüber 75% dem Standarddifferenzierungsprotokoll mit 1%iger DMSO-Gabe nach einem Tag (5+1d). Nach 5+5d sind bei reduzierter DMSO- Konzentration nur ca. 80% der Zellen in Kardiomyozyten differenziert. Auffällig war aber, dass bei der Kultivierung der EBs von 2d-5d sich ca. 70% der EBs (trotz Verwendung der bakteriologischen Petrischalen) anhefteten. Deshalb war es nicht möglich, alle EBs auszuplattieren und zu differenzieren. Die plattierten EBs dagegen hatten keine signifikanten Abweichungen bei der Ausbildung schlagender Zentren (vgl. Abb. 34 B) Expression von CYP1A1 während der myogenen Differenzierung Eine Expression von undifferenzierten P19-Zellen führt zur signifikanten Erhöhung der CYP1A1-Expression, wobei diese Induktion innerhalb von 3h erfolgte (vgl ). Während der myogenen Differenzierung von P19 kommt es nach 2d zu einem geringen, nach 5, 5+5d zum drastischen Anstieg (13 fach) der CYP1A1-Menge (vgl. Abb. 35) in behandelten Zellen gegenüber unbehandelten Kontrollzellen.
20 A %DMSO Kontrolle 10nM / 1%DMSO Anteil schlagender EBs [%] d 5+2d 5+3d 5+4d 5+5d myogene Differenzierung in Tagen [d] B ,25x10-4 % DMSO Kontrolle 10nM ohne DMSO Anteil schlagender EBs [%] d 5+2d 5+3d 5+4d 5+5d myogene Differenzierung in Tagen [d] Abb. 34: Bestimmung der Anzahl kontraktiler Kardiomyozytenzentren während der myogenen Differenzierung von P19-ECC A Kultur der P19-ECC nach Standardprotokoll (vgl ) der Differenzierung mit 1%DMSO und Zusatz von 10nM N=2 n=72 B Differenzierung nur mit 10nM oder 6,25x10-4 %iger DMSO (Kontrolle) N=3 n=24 65
21 Nach 15 Tagen (5+10d) der Differenzierung erreicht die Transkriptmenge das Ausgangsniveau, vergleichbar auch mit der Menge in undifferenzierten Zellen. Obwohl nur für 2 Tage im Medium vorhanden war (0-2d, während der hanging-drop Phase), führt es zu einer signifikant erhöhten CYP1A1-Expression in den sich myogen differenzierenden P19-Zellen, was auf einen Langzeiteffekt von hindeutet. 16 CYP1A1 mrna-menge [rel. E.] DMSO-Kontrolle d 5d 5+5d 5+10d myogene Differenzierung [d] Abb. 35: CYP1A1 mrna-expression während der myogenen Differenzierung und unter -Einfluss N=3 n=2; p 0,05, 2 p 0,001 Signifikanz zu P19; p 0,05, 2 p 0,001Signifikanz zu DMSO mrna-expression von α-mhc, MyoD und Mash-1 während der myogenen Differenzierung Das Zytoskelettprotein α-mhc ist ein Marker für Kardiomyozyten, während MyoD1 als ein Transkriptionsfaktor vor allem bei der Differenzierung von Skelettmuskelzellen eine Rolle spielt. Als ein Marker des Neuroektoderms wurde der Transkriptionsfaktor Mash-1 gewählt. Alle 3 Gene werden bereits in den undifferenzierten P19-ECC exprimiert, was einen relativen Bezug bei der Real-time PCR zu undifferenzierten Kontrollzellen ermöglicht (undifferenzierte P19-ECC =1 rel. E., Abb. 36). Expression von α-mhc: Während der myogenen Differenzierung ist die α-mhc-menge bis Tag 5+5 erhöht und verringert sich an Tag 5+10d. Durch -Inkubation ist die α-mhc- Menge an 5+5d signifikant ca rel. E. erhöht und nach 5+10d signifikant um ca rel. E. verringert. Die Mengenverhältnisse im Kurvenverlauf zeigen eine Verschiebung des Anstieges zugunsten der -induzierten Zellen, was darauf hindeutet, dass die Differenzierung unter -Einfluss im Vergleich zur normalen Differenzierung schneller 66
22 abläuft. Die α-mhc-menge hat mit rel. E. einen sehr hohen Zuwachs während der Differenzierung. Expression von MyoD: Die MyoD-Menge steigt an Tag 5 der Differenzierung kontinuierlich an (max. 38,70 relative Einheiten). Dabei kommt es zu keinem signifikantem Unterschied zwischen Kontroll- und - behandelten Zellen. Expression von Mash-1: Die Induktion von Mash-1 an Tag 5 der Differenzierung fiel durch signifikant geringer aus (32 rel. E.) als in den unbehandelten Zellen mit 52 rel. E., was auf eine verringerte neuronale Differenzierung hindeutet. Insgesamt konnte gezeigt werden, dass die Expression von α-mhc, MyoD und Mash-1 beeinflusst Einfluss von auf die Expression der Glukosetransporter in P19-ECC In adulten Geweben sind die Glukosetransporter-Isoformen zellspezifisch exprimiert, wobei meist 1-2 Isoformen pro Zelltyp ausreichend sind, um den Transport von Glukose zu gewährleisten. So haben z.b. Skelett- bzw. Herzmuskelzellen 2 Isoformen (GLUT1, 4), wobei GLUT1 den basalen Glukoseumsatz transportiert, während bei Insulinstimulation zusätzlich GLUT4 aktiviert wird. Im Gegensatz dazu besitzen undifferenzierte ES- und EC-Zellen 3-4 GLUT-Isoformen (vgl. 3.1). Der Vergleich der Transkriptmengen ermöglicht es, die Expression der Isoformen während der myogenen Differenzierung zu betrachten. beeinflusst den Glukosemetabolismus und stört den Energiestoffwechsel. Diese Wirkung kann mit Veränderungen in der mrna- und Proteinexpression von GLUT1 und GLUT4 korreliert werden (Liu und Matsumura, 1995; Phillips et al., 1995). Am Stammzellmodell P19-ECC wurde deshalb untersucht, wie sich auf die Expression von GLUT1, 3, 4 und 8 während der myogenen Differenzierung auswirkt. Dazu wurde in -exponierten Zellen: a) Die mrna-mengen von GLUT1, 3, 4, 8 mittels Real-time PCR ermittelt (vgl ). b) Die Proteinmengen für GLUT1, GLUT3 und GLUT4 mittels quantitativem Western Blot untersucht (vgl ). c) Das Protein von GLUT1, GLUT3 und GLUT4 nach 5+5d myogener Differenzierung mittels Immunfluoreszenz lokalisiert (vgl ). d) Die Translokation von GLUT1 und GLUT4 wurden mittels EGFP-full-size Klone untersucht. Es wurden GLUT1 und GLUT4 ausgewählt, da diese Transporter zwischen Zytoplasma und Membran translozieren (vgl ). e) Die Phosphorylierung von Akt durch Insulin wurde mittels Western Blot mit Akt/ phospho Akt Antikörpern bestimmt (vgl ). f) Die Aufnahme von Glukose in 2d und 5d EBs wurde mittels Tritum markierter 3-OMG quantifiziert (vgl ). 67
23 1e+5 α-mhc α-mhc mrna-menge [Logarithmus rel. E.] 1e+4 1e+3 1e+2 1e+1 DMSO-Kontrolle 2 2d 5d 5+5d 5+10d myogene Differenzierung [d] 50 MyoD MyoD mrna-menge [rel. E.] DMSO-Kontrolle d 5d 5+5d 5+10d myogene Differenzierung [d] Mash-1 Mash-1 mrna-menge [rel. E.] DMSO-Kontrolle 0 2d 5d 5+5d 5+10d myogene Differenzierung [d] Abb. 36: mrna-menge von α-mhc, MyoD und Mash-1 während der myogenen Differenzierung von P19-ECC 68 Die relativen mrna-mengen wurden durch Real-time PCR in 2, 5, 5+5 und 5+10 Tagen (d) differenzierten P19-ECC ermittelt. Die Transkriptmenge in undifferenzierten Zellen wurden als Bezugspunkt der Berechnung mit 1 festgelegt. N=3 n=2; p 0,05, 2 p 0,001 Signifikanz zu P19; p 0,05, 2 p 0,001 Signifikanz zu DMSO
24 GLUT1 mrna- und Proteinexpression während der myogenen Differenzierung Die Transkriptmenge von GLUT1 ist in myogen differenzierenden Zellen verschiedener Kulturdauer annähernd konstant und vergleichbar mit dem Niveau in undifferenzierten P19- ECC (=1 rel. E.) (vgl. Abb. 37). Nur eine geringe Steigerung auf das ca. 1,5fache an 5 und 5+5d kultivierten EBs ist zu messen. Mit -behandelte Zellen zeigen signifikante Veränderungen in der GLUT1 mrna-menge, wobei die Transkriptmenge an Tag 2 der Differenzierung erhöht ist (ca. 1,5fach) und dann kontinuierlich bis auf die Hälfte abfällt. Auf der Proteinebene führt dagegen zu einer signifikanten Verringerung um ca. 20% (5+5d) und 100% (5+10d) GLUT3 mrna- und Proteinexpression während der myogenen Differenzierung GLUT3 ist ein Transporter mit hoher Glc-Affinität, der im adulten Organismus hauptsächlich in neuronalen Zellen vorkommt. Während der spontanen Differenzierung von embryonalen Stammzellen wird er differentiell exprimiert. Während der Differenzierung der P19-Zellen war dieser Transporter konstitutiv exprimiert. Die Wirkung von auf die GLUT3-Menge wurde sowohl auf mrna-(vgl. Abb. 38 A), als auch Proteinebene (vgl. Abb. 38 B) ermittelt. Dabei wird deutlich, dass zu Abweichungen in der Transkriptmenge abhängig von der Kulturphase führt. Während die Transkriptmenge im Vergleich zu undifferenzierten Zellen nach 2 Tagen stark verringert war, stieg sie dann bis Tag 5 an und hatte dort ein Maximum mit 1,698 rel. E.. Danach verringerte sich die Expression wieder bis auf 0,789 rel. E.. Bei der Zugabe von kam es zu einem signifikanten Anstieg der Expression nach 2d, aber nach 5+10d zu einer Expressionsverringerung um 0,5 rel. E.. Diese Expressionsabnahme korrelierte mit den Daten des neuronalen Markers Mash-1, dessen Expression unter -Einfluss nach 5+10d signifikant reduziert war (Kontrolle 7,58 rel. E.; 4,586 rel. E.). Auf der Proteinebene führt dagegen zu einer signifikanten Verringerung um ca %. Generell steigt die Menge des GLUT3 Proteins von Tag 2d bis 5+10d an GLUT4 mrna- und Proteinexpression während der myogenen Differenzierung Die Expression von GLUT4 unter -Einfluss wurde mittels Real-time PCR für die mrna-expresssion (vgl. Abb. 39 A) und Western Blot für die Proteinexpression (vgl. Abb. 39 B, C) untersucht. Während der ersten Phase der myogenen Differenzierung (2-5d) steigt sowohl die mrna- als auch die Proteinmenge von GLUT4 an. führt in dieser Phase zur Verringerung von GLUT4 (2d: RNA um 50%, Protein 20%). Ab Tag 5 der Differenzierung nimmt die Transkriptmenge kontinuierlich ab, wobei die Proteinmenge konstant bleibt. Nach 5+10d ist die GLUT4 mrna-menge wieder gleich den unbehandelten P19-ECC, während das GLUT4- Protein in den DMSO-differenzierten Zellen stark erhöht ist (vgl. Abb. 39 B). 69

25 A 2,0 GLUT1 mrna-menge [rel. E.] 1,6 1,2 0,8 0,4 DMSO-Kontrolle 2 0,0 2d 5d 5+5d 5+10d myogene Differenzierung [d] B GLUT1 β-aktin ECC 2d 5d 5+5d 5+10d T K T K T K T K 55kd 43kd C 1,6 DMSO-Kontrolle GLUT1 Proteinmenge [rel. E.] 1,4 1,2 1,0 0,8 0,6 2d 5d 5+5d 5+10d myogene Differenzierung [d] Abb. 37: Expression von GLUT1-mRNA (A) und -Protein (B) in myogen differenzierenden P19-ECC A: Quantifizierung der GLUT1 mrna-menge mittels Real-time PCR B: Western Blot für GLUT1 mit Aktin als Ladekontrolle C: Densiometrische Auswertung des Western Blots zum GLUT1-Proteingehalt von in behandelten und Kontroll-EBs im Vergleich zu P19-ECC. N=3 n=2; p 0,05, 2 p 0,001 Signifikanz zu P19; p 0,05, 2 p 0,001 Signifikanz zu DMSO 70
26 A 2,0 DMSO-Kontrolle GLUT3 mrna-menge [rel. E.] 1,6 1,2 0,8 0,4 2 0,0 2d 5d 5+5d 5+10d myogene Differenzierung [d] B GLUT3 β-aktin ECC 2d 5d 5+5d 5+10d T K T K T K T K 45kd 43kd C 4 DMSO-Kontrolle GLUT3 Proteinmenge [rel. E.] d 5d 5+5d 5+10d myogene Differenzierung [d] Abb. 38: GLUT3 mrna- und Proteinexpression während der myogenen Differenzierung A: Quantifizierung der GLUT3 mrna-menge mittels Real-time PCR B: Western Blot für GLUT3 mit Aktin als Ladekontrolle C: Densiometrische Auswertung des Western Blot zum GLUT3-Proteingehalt von behandelten und Kontroll-EBs im Vergleich zu P19-ECC. N=3 n=2; p 0,05, 2 p 0,001 Signifikanz zu P19; p 0,05, 2 p 0,001 Signifikanz zu DMSO 71

27 A 4 DMSO-Kontrolle GLUT4 mrna-menge [rel. E.] d 5d 5+5d 5+10d myogene Differenzierung [d] B GLUT4 2d 5d 5+5d 5+10d T K T K T K T K 54kd β-aktin 43kd C 2,0 DMSO-Kontrolle GLUT4 Proteinmenge [rel. E.] 1,5 1,0 0,5 0,0 2d 5d 5+5d 5+10d myogene Differenzierung [d] Abb. 39: GLUT4 mrna- und Proteinexpression während der myogenen Differenzierung A: Quantifizierung der GLUT4 mrna-menge mittels Real-time PCR B: Western Blot für GLUT4 mit Aktin als Ladekontrolle C: Densiometrische Auswertung des Western Blots zum GLUT4-Proteingehalt von behandelten und Kontroll-EBs im Vergleich zur 2d EB-Kontrolle. N=3 n=2; p 0,05, 2 p 0,001 Signifikanz zu P19; p 0,05, 2 p 0,001 Signifikanz zu DMSO 72
28 GLUT8 mrna-expression während der myogenen Differenzierung Die GLUT8 Transkriptmenge ist während der myogenen Differenzierung der P19-Zellen differentiell reguliert (vgl. Abb. 40). Im Vergleich zu undifferenzierten Zellen ist sie an Tag 2 erhöht (ca. 2,4 fach) und nimmt während des weiteren Differenzierungsprozesses wieder ab. An Tag 5+10 steigt sie wieder signifikant an. verändert das Expressionsprofil während der Differenzierung kaum, führt aber zur Verringerung der Transkriptmengen, insbesondere an Tag 5+10, wo die Expression um das 2,5 fache niedriger liegt als bei der Kontrolle. Zusammenfassend ist die mrna-expression von GLUT8 während der Differenzierung durch signifikant verringert, wobei sich das Expressionsprofil nicht ändert. 3,0 GLUT8 mrna-menge [rel. E.] 2,5 2,0 1,5 1,0 0,5 2 DMSO-Kontrolle 2 0,0 2d 5d 5+5d 5+10d myogene Differenzierung [d] Abb. 40: GLUT8 mrna-expression während der myogenen Differenzierung GLUT8 mrna-expression mittels Real-time PCR bezogen auf P19-ECC (=1 rel.e.). N=3 n=2; p 0,05, 2 p 0,001 Signifikanz zu P19; p 0,05, 2 p 0,001 Signifikanz zu DMSO Lokalisation von GLUT1, 3 und 4 in in vitro myogen differenzierten P19 5+5d EBs Die Lokalisation von GLUT1, 3 und 4 wurde in 5+5d differenzierten P19-ECC untersucht. Um die differenzierten Muskelzellen zu lokalisieren, wurden die myogenen EB-Zellen mittels Desmin-Antikörper markiert. Desmin ist ein Filament kardiomyogener Zellen. GLUT1 liegt in 5+5d myogen differenzierten Zellen in Zytoplasma und Membran der Zellen vor. In den -behandelten Zellen ist die GLUT1-Fluoreszenz in den Kardiomyozyten schwächer als in den Kontrollen. GLUT1 ist im Zytoplasma der Zellen kernnah detekierbar (vgl. Abb. 41 A, B). GLUT3 liegt im Gegensatz zu undifferenzierten (Membranfärbung, Daten nicht gezeigt) in den 5+5d differenzierten Zellen im Zytoplasma unmittelbar um den Zellkern (vgl. Abb. 41 C). In -behandelten Zellen ist das GLUT3-Protein auch im Zytoplasma kernnah lokalisiert, aber es nimmt eine gerichtete Lage innerhalb der Zellen um den Zellkern ein (vgl. Abb. 41 C, 73







, GLUT3 (C, D)")

 differenziert.")


29 3 Ergebnisse D). GLUT4 liegt in den 5+5d differenzierten Zellen im Kern und Zytoplasma der Kardiomyozyten vor. In den -behandelten Zellen befindet sich GLUT4 hauptsächlich im Kern (vgl. Abb. 41 E, F). Unter -Einfluss kommt es am Tag 5+5 der Differenzierung von EB-Zellen zu Kardiomyozyten zu Veränderungen der subzellulären Lokalisation der Glukosetransporterisoformen. -A GLUT1/ Desmin + B GLUT1/ Desmin GLUT1 C GLUT3/ Desmin D GLUT3/ Desmin GLUT3 E GLUT4/ Desmin F GLUT4/ Desmin GLUT4 Abb. 41: Einfluss von auf die GLUT1-, GLUT3- und GLUT4-Lokalisation in 5+5d myogen differenzierten EBs GLUT1 (A, B), GLUT3 (C, D) und GLUT4 (E, F) wurden in 5+5 Tage differenzierten Kardiomyozyten lokalisiert. Die P19-ECC wurden ohne (A, C, E) und mit (B, D, F) differenziert. Die Kardiomyozyten wurden mittels anti-desmin kolokalisiert (rot bei GLUT1, GLUT3, grün bei GLUT4). Durch Doppelmarkierung mit den spezifischen GLUT-Antikörpern wurden die Verteilung der GLUT in den Kardiomyozyten gezeigt. Die Kerne sind zur besseren Orientierung blau gefärbt, die Negativkontrollen in der rechten Ecke dargestellt. Maßstab 20µm. 74



 behandelt (Versuchsschema vgl. 2.4, 2.")


.")

. 5h P19 P19 + DMSO P19 + A B C 15h D E F G H I 25h Abb.")
30 Lokalisation von GLUT1 und GLUT4 mittels pegfp-c3 full size Klone in P19-ECC in vitro GLUT1 und GLUT4 liegen im Zytoplasma in Vesikeln vor und können in die Membran translozieren. Um Studien in vitro durchführen zu können, wurden die P19-ECC mit full-size Klonen für GLUT1 und GLUT4 transfektiert (vgl. 2.4). So konnte die Lokalisation dieser Transporter mittels des EGFP im KLSM bestimmt werden. Expression und Lokalisation von EGFP-GLUT1 in P19-ECC Die Zellen wurden mit EGFP-GLUT1 und den entsprechenden Kontrollen transfektiert und nach 4h mit (10nM) behandelt (Versuchsschema vgl. 2.4, 2.7). Die Zellen wurden für 5, 15 und 25h kultiviert und mikroskopisch ausgewertet (vgl. Abb. 42). Ca. 30% der Zellen zeigten eine grüne Fluoreszenz. Das Fluoreszenzsignal markierte in unbehandelten und DSMO-behandelten Zellen stark die Zellmembran und zu einem geringen Anteil auch das Zytoplasma. Eine deutliche Veränderung zeigt sich bei -behandelten Zellen. Hier liegt das EGFP-GLUT1 Signal in den meisten Zellen nur im Zytoplasma (vgl. Abb. 42, I). Das Signal ist punktförmig und gibt den Zellen ein gekörntes Aussehen, was vermuten lässt, dass sich EGFP-GLUT1 in zytoplasmatischen Vesikeln befindet. Der Einfluss von auf die Lokalisation des GLUT1 wurde durch eine immunhistochemische Detektion bestätigt (vgl ; Abb. 47). 5h P19 P19 + DMSO P19 + A B C 15h D E F G H I 25h Abb. 42: Lokalisation von EGFP-GLUT1 nach 5h, 15h und 25h Nach transienter Transfektion der P19-ECC mit EGFP-GLUT1 wurde die Inkubation mit für 5h (C), 15h (F) und 25h (I) durchgeführt. Unbehandelte Zellen sind in (A, D, G), DMSO Kontrollzellen in (B, E, H) abgebildet. Nach Fixierung in 4% PFA wurde die Fluoreszenz am KLSM bei 488 nm ausgewertet. Die grüne Fluoreszenz ist das EGFP markierte GLUT1. Die Zellkerne wurden mit 7-AAD rot gegengefärbt. 75


. Eine Insulinzugabe verändert die Lokalisation des EGFP-GLUT1 nicht (vgl. Abb. 43).")
 als auch in den behandelten Zellen mit dieser ungewöhnlichen Lokalisation, waren keine")



 untersucht.")
. Maßstab 20µm.")
31 Einfluss von Insulin auf die Lokalisation des EGFP-GLUT1 Die Assoziation der Glukosetransporter mit zytoplasmatischen Vesikeln führt vor allem bei Stimulation mit Insulin zur Auflösung dieser Strukturen und Ausschüttung der Transporter in die Zellmembran. Um zu untersuchen, ob es sich bei der ungewöhnlichen Lokalisation von GLUT1 nach -Zugabe um eine Speicherung in Insulin-sensitiven Vesikeln handelt, wurden die Zellen mit Insulin stimuliert (vgl ). Eine Insulinzugabe verändert die Lokalisation des EGFP-GLUT1 nicht (vgl. Abb. 43). Sowohl in den Kontrollen, in denen EGFP-GLUT in der Zellmembran ist (vgl. Abb. 43 A, B, D, E) als auch in den behandelten Zellen mit dieser ungewöhnlichen Lokalisation, waren keine Unterschiede sichtbar. Es konnte gezeigt werden, dass GLUT1 nach 25h mit 10nM nicht mehr in der Zellmembran, sondern im Zytoplasma vorliegt. Insulin hat keinen Einfluss auf die Lage des EGFP-GLUT1, der hauptsächlich in Vesikeln in Umgebung des Kernes vorliegt. P19 DMSO A B C - Insulin D E F + Insulin EGFP- Kontrolle G H I Abb. 43: Insulin-Einfluss auf die Lokalisation von EGFP-GLUT1 in P19-ECC nach 25h Die Translokation des EGFP-GLUT1 Konstruktes wurde nach 25h Inkubation mit (C), DMSO (B) und der unbehandelten Kontrolle (A) mittels Insulingabe für 2h (vgl ) untersucht. Insulininkubierten P19-ECC sind in D-F, unbehandelte EGFP-Vektor-Kontrollzellen in G, DMSO-behandelte in H und -behandelte in I abgebildet. Die Kerne sind rot gefärbt (7-AAD). Maßstab 20µm. Einfluss von α-nf auf die Lokalisation des EGFP-GLUT1 Zur Untersuchung der α-nf-wirkung wurde der AhR vor -Stimulation mit 10µM α NF blockiert. In den behandelten Zellen lag GLUT1 nach α NF Inhibierung des AhR membranständig vor (vgl. Abb. 44 A), während er perinukleär in nur mit inkubierten Zellen lokalisiert war (vgl. Abb. 44 B). 76


, wohingegen nur EGFP mit ca. 28 kd vom Kontrollplasmid translatiert wird (wie in Abb. 45 A, B Spur2).")
32 A B + α NF 10µm Abb. 44: α NF-Einfluss auf die Lokalisation von EGFP-GLUT1 in P19-ECC nach 25h Die Translokation des EGFP-GLUT1 Konstruktes wurde nach Inkubation für 2h mit α NF und anschliessender 25h Inkubation mit am Fluoreszenzmikroskop untersucht. Die behandelten Zellen sind in (A), die α NF und inkubierten in (B) dargestellt. Die Kerne sind rot gefärbt (7-AAD). Maßstab 10µm. Lokalisation von GLUT4 in P19-ECC mittels EGFP-GLUT4 Überprüfung der Translation des GLUT4-EGFP: Nach Transfektion der P19-Zellen mit dem GLUT4-EGFP bzw. dem EGFP-Vektor (Kontrolle) wurde nach 4h das Zelllysat mittels Western Blot untersucht (vgl. Abb. 45). Bei der Translation eines GLUT4-EGFP- Fusionsproteins muss ein dimäres Protein von ca. 73 kd entstehen (wie in Abb. 45 A, B Spur1 gezeigt), wohingegen nur EGFP mit ca. 28 kd vom Kontrollplasmid translatiert wird (wie in Abb. 45 A, B Spur2). Die Detektion mit einem anti-egfp-antikörper (vgl. Abb. 45 A) und einem anti-glut4-antikörper (vgl. Abb. 45 B) ergab übereinstimmende Ergebnisse, was die Translation eines vollständigen Fusionsprotein der entsprechenden Größe (GLUT4- EGFP, 72kDA) beweist. A kd M 1 2 B kd M ,2 57,9 42 GLUT4-EGFP 66,2 57,9 42 GLUT4-EGFP GLUT4 EGFP Abb. 45: Expression von GLUT4-EGFP in P19-ECC unter - und Insulin-Einfluss A: In Spur1 ist das Gesamtprotein der GLUT4-EGFP transfizierten Zellen aufgetragen, in Spur2 das Gesamtprotein der EGFP transfizierten Kontrollzellen. Durch Detektion mit dem GFP-Antikörper wurde das GLUT4-EGFP bei 73kD in Spur1 (GLUT4 Maus: 45kD plus EGFP:28kD) und das EGFP in Spur 2 bei 28kD detektiert. B: Durch Stripping des Western Blot aus A und Inkubation mit dem GLUT4 Antikörper wurde das endogene GLUT4 (54kd) und das GLUT4-EGFP detektiert. 77
.")
 Zellen durchgeführt (vgl. Abb. 46 B).")



 untersucht.")

33 Zelluläre Expression von GLUT4-EGFP: GLUT4-EGFP liegt nahe am Kern, während nur EGFP (Kontrolle) in der gesamten Zelle homogen verteilt vorliegt (vgl. Abb. 46 A: a, c). Nach Zugabe von 1µg/ml Insulin (2h) translokiert der Transporter in die Zellmembran (vgl. Abb. 46 A: b). Die Insulinstimulation wurde auch an exponierten (25h) Zellen durchgeführt (vgl. Abb. 46 B). Während vor Insulininkubation relativ viel Protein noch im Zytoplasma vorlag, translozierte der Hauptteil nach Insulin-Zugabe in die Membran. Es waren keine signifikanten Unterschiede in der Lokalisation in -exponierten und der Kontrolle (DMSO) zu erkennen, was beweist, dass die Insulin-stimulierte GLUT4-Translokation nicht beeinflusst. A a b c d DMSO B a b - Insulin c d + Insulin Abb. 46: Expression von GLUT4-EGFP in P19-ECC unter - und Insulin-Einfluss Nach transienter Transfektion der P19-ECC mit EGFP-GLUT4 wurde die Translokation von GLUT4 durch Insulin (A) und die Translokation nach 25h -Behandlung mit Insulin (B) untersucht. Nach Fixierung in 4%PFA wurde die Fluoreszenz am KLSM bei 488nm ausgewertet. Die grüne Fluoreszenz ist das EGFP markierte GLUT4. Die Zellkerne wurden mit 7-AAD rot gegengefärbt. Maßstab 8µm. A: Lokalisation von GLUT4 in unbehandelten P19-ECC (a), nach Zugabe von Insulin (2h, b), unbehandelter Kontrolle (d) und EGFP-Kontrolle (c). B: Lokalisation von GLUT4 nach 25h Inkubation (a) und nachfolgender Inkubation für 2h mit Insulin (c). Die DMSO- Kontrolle nach 25h ist in (b) und mit 2h Insulinbehandlung in (d) dargestellt. 78

 wurde die GLUT1-Lage nach -Zugabe mittels anti-glut1")

, aber in einigen Kontrollzellen auch unverändert in")

.")

.")
34 Immunhistochemische Lokalisation von GLUT1 nach Kultur mit Neben der Markierung des GLUT1-Proteins mit EGFP (vgl ; Abb. 42) wurde die GLUT1-Lage nach -Zugabe mittels anti-glut1 Antikörper in der Immunfluoreszenz untersucht. In den behandelten Zellen befindet sich GLUT1 verstärkt im Zytoplasma (vgl. Abb. 47 B, D), aber in einigen Kontrollzellen auch unverändert in der Membran (vgl. Abb. 47 F). Die Anzahl der Zellen, die GLUT1 in der Membran exprimieren, ist in exponierten Zellen deutlich geringer als in unbehandelten Zellen (vgl. Abb. 47 A, C, E). A DMSO B C D E F Abb. 47: GLUT1 Immunfluoreszenz in P19-ECC nach Inkubation Detektion von anti-glut1 (grün, 488nM) mittels KSLM in P19-ECC (grüne Fluoreszenz). Die Zellen wurden für 25h mit DMSO (A, C, E) oder 10nM (B; D, F) behandelt. Nach - Behandlung ist GLUT1 hauptsächlich im Zytoplasma (, B, D) aber auch in einigen Zellen membranständig lokalisiert (, F). Die Zellkerne sind mit 7-AAD rot gefärbt. Maßstab 20µm. 79
 wird durch spezifische Signalwege reguliert.")
35 Einfluss von auf die Phosphorylierung von Akt Die Verteilung der Glukosetransporter in die entsprechenden zellulären Kompartemente (Zellmembran, Vesikel) wird durch spezifische Signalwege reguliert. Die zytoplasmatische Phosphatase Akt ist ein Schlüsselprotein für die Translokation von GLUT4. Da eine Hemmung der Aktivierung von Akt zur Fehlsortierung der GLUT führen könnte, wurde untersucht, ob die Phosphorylierung von Akt in P19-ECC beeinflusst. Weiterhin sollte festgestellt werden, ob die Akt-Phosphorylierung durch über den AhR erfolgt. Dazu wurde eine Versuchsgruppe vor -Zugabe mit dem AhR-Inhibitor α-nf behandelt. Die Phosphorylierung von Akt wurde durch phosphospezifische Antikörper im Western Blot detektiert (vgl. Abb. 48). In allen Versuchsgruppen (DMSO,, + α-nf) kommt es durch Insulin zur Aktivierung von Akt. Die Phosphorylierung von Akt ist durch nicht beeinflusst. DMSO + Insulin - Insulin + IH DMSO +IH pakt 60 kd Akt 60 kd Abb. 48: Detektion der Phosphorylierung von Akt in P19-ECC Nach 25h Inkubation mit wurde die Phosphorylierung von Akt bestimmt. Zur Induktion der Phosphorylierung wurden die einzelnen Versuchsgruppen für 5min mit Insulin behandelt. Das 60kD große Protein wurde mittels Western Blot detektiert. Zur Kontrolle wurde nach Stripping der Membran nicht phosphoryliertes Akt bestimmt. IH steht für den AhR-Inhibitor α-nf Bestimmung der Wirkung von auf die Glukoseaufnahme in 2d und 5d P19-80 EBs Die Aufnahme von Glukose in 2d und 5d differenzierte EBs wurde durch die Messung der Inkorporation von 3 H-OMG bestimmt. Die Messwerte der einzelnen EBs hatten eine weite Streuung ( nmol/cm 3 /3min). Die relative Glukoseaufnahme des EBs wurde nach Formel [1] (vgl. 2.15) berechnet. Von den EBs einer Versuchsgruppe wurde der Mittelwert berechnet (vgl. Abb. 49). Die durchschnittliche Glukoseaufnahme eines EBs liegt bei ca nmol/cm 3 /3min, wobei keine signifikante Änderung in der untersuchten Kulturzeit gemessen wurde. führt sowohl bei 2 als auch bei 5d EBs zur Verringerung der Glc-Aufnahme. Bei 2d kultivierten EBs ist die Verringerung der Glukoseaufnahme nicht signifikant durch verändert (vgl. Abb. 49). Erst in 5d kultivierten EBs kommt es zur signifikanten Verringerung der Glukoseaufnahme um 25%.
4.2 Kokulturen Epithelzellen und Makrophagen
 Ergebnisse 4.2 Kokulturen Epithelzellen und Makrophagen Nach der eingehenden Untersuchung der einzelnen Zelllinien wurden die Versuche auf Kokulturen aus den A549-Epithelzellen und den Makrophagenzelllinien
Ergebnisse 4.2 Kokulturen Epithelzellen und Makrophagen Nach der eingehenden Untersuchung der einzelnen Zelllinien wurden die Versuche auf Kokulturen aus den A549-Epithelzellen und den Makrophagenzelllinien
3 Ergebnisse 3.1 Charakterisierung der untersuchten Melanome
 3 Ergebnisse 3.1 Charakterisierung der untersuchten Melanome Untersucht wurden insgesamt 26 Melanome, die zwischen 1991 und 1997 in der Universitätsklinik und Poliklinik für Dermatologie und Venerologie
3 Ergebnisse 3.1 Charakterisierung der untersuchten Melanome Untersucht wurden insgesamt 26 Melanome, die zwischen 1991 und 1997 in der Universitätsklinik und Poliklinik für Dermatologie und Venerologie
4.1. Herstellung des CH2/CH3-trunkierten dimerisierten anti-cd30-igg1-tnf- Fusionsproteins
 ERGEBNISSE 29 4. Ergebnisse 4.1. Herstellung des CH2/CH3-trunkierten dimerisierten anti-cd3-igg1-tnf- Fusionsproteins Im vorliegenden Immunzytokin wurden die Domänen CH2/CH3 des humanen Fc-Fragmentes durch
ERGEBNISSE 29 4. Ergebnisse 4.1. Herstellung des CH2/CH3-trunkierten dimerisierten anti-cd3-igg1-tnf- Fusionsproteins Im vorliegenden Immunzytokin wurden die Domänen CH2/CH3 des humanen Fc-Fragmentes durch
Kaninchen: 1 cagattgaggagagcaccaccatcgtcacctcgcagttggcggaggtcagcgcagccgag 60
 Ergebnisse 6 3. Ergebnisse 3. Charakterisierung der zelllinienspezifischen cdn Die Entwicklung zweier Zelllinien, des epithelialen Trophoblasten und des pluripotenten Embryoblasten, ist der erste Differenzierungsschritt
Ergebnisse 6 3. Ergebnisse 3. Charakterisierung der zelllinienspezifischen cdn Die Entwicklung zweier Zelllinien, des epithelialen Trophoblasten und des pluripotenten Embryoblasten, ist der erste Differenzierungsschritt
4.1. Herstellung, Aufreinigung und Nachweis eines CD33-spezifischen IgM-Antikörpers
 ERGEBNISSE 30 4. Ergebnisse 4.1. Herstellung, Aufreinigung und Nachweis eines CD33-spezifischen IgM-Antikörpers 4.1.1. Herstellung und Aufreinigung des Antikörpers CD33-spezifische IgM-Antikörper wurden
ERGEBNISSE 30 4. Ergebnisse 4.1. Herstellung, Aufreinigung und Nachweis eines CD33-spezifischen IgM-Antikörpers 4.1.1. Herstellung und Aufreinigung des Antikörpers CD33-spezifische IgM-Antikörper wurden
Drexler G.A. 1, Derer A 1, Dirks W.G. 2, Hable V. 3, Greubel C. 3, Burgdorfer C. 3, Dollinger G. 3, Du G. 4, Friedl A.A. 1
 Nutzung von bi-cistronischen Vektoren für die Beobachtung der Rekrutierung von Signal- und Reparaturproteinen an DNA-Schäden nach Ionen- Mikrobestrahlung durch Live-Cell Imaging Drexler G.A. 1, Derer A
Nutzung von bi-cistronischen Vektoren für die Beobachtung der Rekrutierung von Signal- und Reparaturproteinen an DNA-Schäden nach Ionen- Mikrobestrahlung durch Live-Cell Imaging Drexler G.A. 1, Derer A
Die RT-PCR Untersuchungen wurden anhand 27 Struma nodosa- (siehe Tab. 7) und 15 Adenom- (siehe Tab. 11) Präparaten durchgeführt.
 und 15 Adenom- (siehe Tab. 11) Präparaten durchgeführt.") - 22-4 Ergebnisse 4.1 RT-PCR Die RT-PCR Untersuchungen wurden anhand 27 Struma nodosa- (siehe Tab. 7) und 15 Adenom- (siehe Tab. 11) Präparaten durchgeführt. 4.1.1 Struma nodosa Histologie Fas Fas CD 97
- 22-4 Ergebnisse 4.1 RT-PCR Die RT-PCR Untersuchungen wurden anhand 27 Struma nodosa- (siehe Tab. 7) und 15 Adenom- (siehe Tab. 11) Präparaten durchgeführt. 4.1.1 Struma nodosa Histologie Fas Fas CD 97
4. ERGEBNISSE. 4.2. Untersuchung des umgelagerten IgH Gens in Hodgkinzelllinien
 36 4. ERGEBNISSE 4.1. Immunglobulin Gentranskripte in Hodgkinzelllinien Mit Hilfe der RT PCR untersuchten wir die Expression umgelagerter Ig Gene in den Hodgkinzelllinien L1236, L428, L591 und KM-H2 sowie
36 4. ERGEBNISSE 4.1. Immunglobulin Gentranskripte in Hodgkinzelllinien Mit Hilfe der RT PCR untersuchten wir die Expression umgelagerter Ig Gene in den Hodgkinzelllinien L1236, L428, L591 und KM-H2 sowie
3 Ergebnisse. 3.1 Nachweis der Expression des y + -Systems in Keratinozyten und HDMEC. 3.1.1 Nachweis auf RNA-Ebene. Ergebnisse 34
 Ergebnisse 34 3 Ergebnisse 3.1 Nachweis der Expression des y + -Systems in Keratinozyten und HDMEC 3.1.1 Nachweis auf RNA-Ebene Zum Nachweis der Transkription des y + -Systems in kutanen Zellen, wurden
Ergebnisse 34 3 Ergebnisse 3.1 Nachweis der Expression des y + -Systems in Keratinozyten und HDMEC 3.1.1 Nachweis auf RNA-Ebene Zum Nachweis der Transkription des y + -Systems in kutanen Zellen, wurden
4.1 Interaktion von Envelope Proteinen mit dem spannungsabhängigen Kalium Kanal BEC-1
 4. Ergebnisse 60 4 ERGEBNISSE 4.1 Interaktion von Envelope Proteinen mit dem spannungsabhängigen Kalium Kanal BEC-1 Retrovirale Envelope Proteine spielen eine entscheidende Rolle beim Eindringen des Virus
4. Ergebnisse 60 4 ERGEBNISSE 4.1 Interaktion von Envelope Proteinen mit dem spannungsabhängigen Kalium Kanal BEC-1 Retrovirale Envelope Proteine spielen eine entscheidende Rolle beim Eindringen des Virus
7 Anhang A. 7 Anhang A. Abbildungsverzeichnis
 7 Anhang A Abbildungsverzeichnis Abb. 1: Allgemeine chemische Struktur von PCDDs, PCDFs und TCDD... 2 Abb. 2: AhR-Signalkaskade und Blockierung des AhR durch AhRR modifiziert nach (Mimura und Fujii-Kuriyama,
7 Anhang A Abbildungsverzeichnis Abb. 1: Allgemeine chemische Struktur von PCDDs, PCDFs und TCDD... 2 Abb. 2: AhR-Signalkaskade und Blockierung des AhR durch AhRR modifiziert nach (Mimura und Fujii-Kuriyama,
β2-microglobulin deficient mice lack CD4-8+cytolytic T cells
 β2-microglobulin deficient mice lack CD4-8+cytolytic T cells Mäuse mit einem Knock-out bezüglich ß-Microglobulin sind nicht in der Lage CD4-8+ cytotoxische T-Zellen zu bilden Nature,Vol 344, 19. April
β2-microglobulin deficient mice lack CD4-8+cytolytic T cells Mäuse mit einem Knock-out bezüglich ß-Microglobulin sind nicht in der Lage CD4-8+ cytotoxische T-Zellen zu bilden Nature,Vol 344, 19. April
Dissertation zur Erlangung des akademischen Grades Doktor der Medizin (Dr. med.)
") Aus dem Institut für Anatomie und Zellbiologie. an der Martin-Luther-Universität Halle-Wittenberg (Direktor: Prof. Dr. med. Dr. agr. Bernd Fischer) und aus der Universität Göttingen Sektion Embryologie
Aus dem Institut für Anatomie und Zellbiologie. an der Martin-Luther-Universität Halle-Wittenberg (Direktor: Prof. Dr. med. Dr. agr. Bernd Fischer) und aus der Universität Göttingen Sektion Embryologie
4. Diskussion. 4.1. Polymerase-Kettenreaktion
 59 4. Diskussion Bei der Therapie maligner Tumoren stellt die Entwicklung von Kreuzresistenz gegen Zytostatika ein ernstzunehmendes Hindernis dar. Im wesentlich verantwortlich für die so genannte Multidrug
59 4. Diskussion Bei der Therapie maligner Tumoren stellt die Entwicklung von Kreuzresistenz gegen Zytostatika ein ernstzunehmendes Hindernis dar. Im wesentlich verantwortlich für die so genannte Multidrug
Marianti Manggau. Untersuchungen zu Signalwegen von SPP in humanen Keratinozyten
 Marianti Manggau Untersuchungen zu Signalwegen von SPP in humanen Keratinozyten Die Deutsche Bibliothek - CIP-Einheitsaufnahme Manggau, Marianti: Untersuchungen zu Signalwegen von SPP in humanen Keratinozyten
Marianti Manggau Untersuchungen zu Signalwegen von SPP in humanen Keratinozyten Die Deutsche Bibliothek - CIP-Einheitsaufnahme Manggau, Marianti: Untersuchungen zu Signalwegen von SPP in humanen Keratinozyten
Ergebnisse Immunhistochemie
 3.2.2.6 Immunhistochemie Zum Nachweis von Mikrometasasen wurden Antikörper gegen humanes Zytokeratin verwendet. Da alle implantierten Tumorgewebe epithelialen Ursprungs waren, konnte dieser Marker bei
3.2.2.6 Immunhistochemie Zum Nachweis von Mikrometasasen wurden Antikörper gegen humanes Zytokeratin verwendet. Da alle implantierten Tumorgewebe epithelialen Ursprungs waren, konnte dieser Marker bei
Wirkung der Blockade des Angiotensin II ATi-Rezeptors auf die Funktion und die Struktur des Herzens der Streptozotodn-diabetischen Ratte
 Wirkung der Blockade des Angiotensin II ATi-Rezeptors auf die Funktion und die Struktur des Herzens der Streptozotodn-diabetischen Ratte Inaugural-Dissertation zur Erlangung des Doktorgrades der Mathematisch-Naturwissenschaftlichen
Wirkung der Blockade des Angiotensin II ATi-Rezeptors auf die Funktion und die Struktur des Herzens der Streptozotodn-diabetischen Ratte Inaugural-Dissertation zur Erlangung des Doktorgrades der Mathematisch-Naturwissenschaftlichen
Hendrik A. Wolff, Can M. Sag, Kay Neumann, Marie-Kristin Opiela, Margret Rave-Fränk, Clemens F. Hess, Lars S. Maier, Hans Christiansen
 Ionisierende Strahlung (IR) führt über Reactive Oxygen Species (ROS) und aktivierte CaMKinase II zu akuten Veränderungen im kardialen Calciumstoffwechsel und zu akuter Schädigung isolierter Kardiomyocyten
Ionisierende Strahlung (IR) führt über Reactive Oxygen Species (ROS) und aktivierte CaMKinase II zu akuten Veränderungen im kardialen Calciumstoffwechsel und zu akuter Schädigung isolierter Kardiomyocyten
Stand von letzter Woche
 RUB ECR1 AXR1 Stand von letzter Woche E2 U? E1-like AXR1 Repressor ARF1 Proteasom AuxRE Repressor wird sehr schnell abgebaut notwendig für Auxinantwort evtl. Substrat für SCF Identifikation des SCF-Ubiquitin
RUB ECR1 AXR1 Stand von letzter Woche E2 U? E1-like AXR1 Repressor ARF1 Proteasom AuxRE Repressor wird sehr schnell abgebaut notwendig für Auxinantwort evtl. Substrat für SCF Identifikation des SCF-Ubiquitin
Die Connexine Cx33 und Cx43 werden in der Retina der Ratte tageszeitlich reguliert
 Aus dem Institut für Funktionelle und Klinische Anatomie der Universitätsmedizin der Johannes Gutenberg-Universität Mainz Die Connexine Cx33 und Cx43 werden in der Retina der Ratte tageszeitlich reguliert
Aus dem Institut für Funktionelle und Klinische Anatomie der Universitätsmedizin der Johannes Gutenberg-Universität Mainz Die Connexine Cx33 und Cx43 werden in der Retina der Ratte tageszeitlich reguliert
3.3 Wt1 ist notwendig für die Entwicklung der Milz
 3.3 Wt1 ist notwendig für die Entwicklung der Milz Während der Embryogenese wird WT1 in verschiedenen Organen exprimiert. Dazu gehören Nieren, Gonaden, die stromalen Zellen der Milz sowie mesotheliale
3.3 Wt1 ist notwendig für die Entwicklung der Milz Während der Embryogenese wird WT1 in verschiedenen Organen exprimiert. Dazu gehören Nieren, Gonaden, die stromalen Zellen der Milz sowie mesotheliale
3.2.1 Neurogeneserate der magnetfeldbehandelten Tiere aus restriktiver Haltung
 Eigene Untersuchungen 25 3.2 Ergebnisse 3.2.1 Neurogeneserate der magnetfeldbehandelten Tiere aus restriktiver Haltung Untersucht wurde, ob die Magnetfeldbehandlung mit 1, 8, 12, 29 und 5 Hz einen Einfluss
Eigene Untersuchungen 25 3.2 Ergebnisse 3.2.1 Neurogeneserate der magnetfeldbehandelten Tiere aus restriktiver Haltung Untersucht wurde, ob die Magnetfeldbehandlung mit 1, 8, 12, 29 und 5 Hz einen Einfluss
4.6. MR-tomographische Untersuchung an Leber, Milz und Knochenmark
 4.6. MR-tomographische Untersuchung an Leber, Milz und Knochenmark Die folgenden Darstellungen sollen einen Überblick über das Signalverhalten von Leber, Milz und Knochenmark geben. Die Organe wurden zusammen
4.6. MR-tomographische Untersuchung an Leber, Milz und Knochenmark Die folgenden Darstellungen sollen einen Überblick über das Signalverhalten von Leber, Milz und Knochenmark geben. Die Organe wurden zusammen
Ergebnisse. I. Identifizierung intrazellulärer Interaktionspartner. Ergebnisse I. Identifizierung intrazellulärer Interaktionspartner
 Ergebnisse I. Identifizierung intrazellulärer Interaktionspartner Es sollten mithilfe des Yeast-Two-Hybrid-Systems Proteine identifiziert werden, die mit der zytoplasmatischen Domäne von CEACAM1-4L aus
Ergebnisse I. Identifizierung intrazellulärer Interaktionspartner Es sollten mithilfe des Yeast-Two-Hybrid-Systems Proteine identifiziert werden, die mit der zytoplasmatischen Domäne von CEACAM1-4L aus
Medizinische Immunologie. Vorlesung 6 Effektormechanismen
 Medizinische Immunologie Vorlesung 6 Effektormechanismen Effektormechanismen Spezifische Abwehrmechanismen Effektormechanismen der zellulären Immunantwort - allgemeine Prinzipien - CTL (zytotoxische T-Lymphozyten)
Medizinische Immunologie Vorlesung 6 Effektormechanismen Effektormechanismen Spezifische Abwehrmechanismen Effektormechanismen der zellulären Immunantwort - allgemeine Prinzipien - CTL (zytotoxische T-Lymphozyten)
Übung 8. 1. Zellkommunikation. Vorlesung Bio-Engineering Sommersemester 2008. Kapitel 4. 4
 Bitte schreiben Sie Ihre Antworten direkt auf das Übungsblatt. Falls Sie mehr Platz brauchen verweisen Sie auf Zusatzblätter. Vergessen Sie Ihren Namen nicht! Abgabe der Übung bis spätestens 05. 05. 08
Bitte schreiben Sie Ihre Antworten direkt auf das Übungsblatt. Falls Sie mehr Platz brauchen verweisen Sie auf Zusatzblätter. Vergessen Sie Ihren Namen nicht! Abgabe der Übung bis spätestens 05. 05. 08
Ergebnisse 32. Peptid 1- Peptid 2- Peptid 1- Peptid 2- Sepharose Sepharose Sepharose Sepharose 1/I 1/II 2/I 2/II
 Ergebnisse 32 4 Ergebnisse 4.1 Nachweis des ORF1-Proteins p40 Das erste offene Leseraster (ORF1) eines L1-Retrotransposons kodiert für ein 40 kd Protein (p40-protein). Dieses Protein hat eine Chaperone-Funktion
Ergebnisse 32 4 Ergebnisse 4.1 Nachweis des ORF1-Proteins p40 Das erste offene Leseraster (ORF1) eines L1-Retrotransposons kodiert für ein 40 kd Protein (p40-protein). Dieses Protein hat eine Chaperone-Funktion
Johannes Gutenberg-Universität Mainz Fachbereich Biologie, Institut für Zoologie Leitung: Prof. Dr. Wolfrum Datum: 15. April 2005
 Johannes Gutenberg-Universität Mainz Fachbereich Biologie, Institut für Zoologie Leitung: Prof. Dr. Wolfrum Datum: 15. April 2005 F1 Molekulare Zoologie Teil II: Expressionsanalyse Proteinexpressionsanalyse
Johannes Gutenberg-Universität Mainz Fachbereich Biologie, Institut für Zoologie Leitung: Prof. Dr. Wolfrum Datum: 15. April 2005 F1 Molekulare Zoologie Teil II: Expressionsanalyse Proteinexpressionsanalyse
Schlussbericht zur Studienwoche Biologie und Medizin vom 17. 23. März 2013
 Schlussbericht zur Studienwoche Biologie und Medizin vom 17. 23. März 2013 Projekt: Zellbiologie: Expression und Reinigung der onkogenen Kinase Abl an der Ecole Polytechnique Fédérale de Lausanne, EPFL
Schlussbericht zur Studienwoche Biologie und Medizin vom 17. 23. März 2013 Projekt: Zellbiologie: Expression und Reinigung der onkogenen Kinase Abl an der Ecole Polytechnique Fédérale de Lausanne, EPFL
Charakterisierung von Tumor initiierenden Zellen mit randomisierten Phagenpeptidbanken
 Charakterisierung von Tumor initiierenden Zellen mit randomisierten Phagenpeptidbanken S. Adebahr 1, M. Henke 1, R. Mertelsmann 2, G. Niedermann 1, M. Trepel 2,3 1 Klinik für Strahlenheilkunde, Universitätsklinikum
Charakterisierung von Tumor initiierenden Zellen mit randomisierten Phagenpeptidbanken S. Adebahr 1, M. Henke 1, R. Mertelsmann 2, G. Niedermann 1, M. Trepel 2,3 1 Klinik für Strahlenheilkunde, Universitätsklinikum
Metabolische Veränderungen und Zelltod in neuralen Zellen. durch Advanced Glycation Endproducts -
 Metabolische Veränderungen und Zelltod in neuralen Zellen durch Advanced Glycation Endproducts - Implikationen für die Pathogenese der Alzheimer schen Demenz Dissertation zur Erlangung des naturwissenschaftlichen
Metabolische Veränderungen und Zelltod in neuralen Zellen durch Advanced Glycation Endproducts - Implikationen für die Pathogenese der Alzheimer schen Demenz Dissertation zur Erlangung des naturwissenschaftlichen
Stammzellenmanipulation. Stammzellen können in Zellkultur manipuliert werden
 Stammzellenmanipulation Hämatopoietische Stammzellen können gebraucht werden um kranke Zellen mit gesunden zu ersetzen (siehe experiment bestrahlte Maus) Epidermale Stammzellpopulationen können in Kultur
Stammzellenmanipulation Hämatopoietische Stammzellen können gebraucht werden um kranke Zellen mit gesunden zu ersetzen (siehe experiment bestrahlte Maus) Epidermale Stammzellpopulationen können in Kultur
Übung 11 Genregulation bei Prokaryoten
 Übung 11 Genregulation bei Prokaryoten Konzepte: Differentielle Genexpression Positive Genregulation Negative Genregulation cis-/trans-regulation 1. Auf welchen Ebenen kann Genregulation stattfinden? Definition
Übung 11 Genregulation bei Prokaryoten Konzepte: Differentielle Genexpression Positive Genregulation Negative Genregulation cis-/trans-regulation 1. Auf welchen Ebenen kann Genregulation stattfinden? Definition
Entwicklung und Validierung von multiplex real-time RT-PCR assays in der Virusdiagnostik
 Entwicklung und Validierung von multiplex real-time RT-PCR assays in der Virusdiagnostik Bernd Hoffmann, Klaus R. Depner, Horst Schirrmeier & Martin Beer Friedrich-Loeffler-Institut Greifswald-Insel Riems
Entwicklung und Validierung von multiplex real-time RT-PCR assays in der Virusdiagnostik Bernd Hoffmann, Klaus R. Depner, Horst Schirrmeier & Martin Beer Friedrich-Loeffler-Institut Greifswald-Insel Riems
3.2 Analyse des Signaltransduktionsweges, über den Hyaluronsäure die Aktivierung des MMP-9-Gens vermittelt
 3.2 Analyse des Signaltransduktionsweges, über den Hyaluronsäure die Aktivierung des MMP-9-Gens vermittelt 3.2.1 Hyaluronsäure vermittelt die transkriptionelle Aktivierung des MMP-9-Gens und nicht die
3.2 Analyse des Signaltransduktionsweges, über den Hyaluronsäure die Aktivierung des MMP-9-Gens vermittelt 3.2.1 Hyaluronsäure vermittelt die transkriptionelle Aktivierung des MMP-9-Gens und nicht die
Die Rolle des NLRP3- Inflammasom in der Pathogenese der idiopathischen pulmonalen Fibrose und Sarkoidose
 Aus der Medizinischen Universitätsklinik und Poliklinik Abteilung für Pneumologie der Albert-Ludwigs-Universität Freiburg im Breisgau Die Rolle des NLRP3- Inflammasom in der Pathogenese der idiopathischen
Aus der Medizinischen Universitätsklinik und Poliklinik Abteilung für Pneumologie der Albert-Ludwigs-Universität Freiburg im Breisgau Die Rolle des NLRP3- Inflammasom in der Pathogenese der idiopathischen
Abbildungsverzeichnis
 I INHALTSVERZEICHNIS Abkürzungen VI Abbildungsverzeichnis VIII I. Einleitung 1. Neurone und Axonwachstum 1 2. Oligodendrozyten und Myelin 3 3. Das Proteolipid Protein (PLP) 6 4. Mutationen im PLP-Gen und
I INHALTSVERZEICHNIS Abkürzungen VI Abbildungsverzeichnis VIII I. Einleitung 1. Neurone und Axonwachstum 1 2. Oligodendrozyten und Myelin 3 3. Das Proteolipid Protein (PLP) 6 4. Mutationen im PLP-Gen und
Autismus. autism spectrum disorders
 Autismus autism spectrum disorders Index Was ist Autismus? Wissenschaftliche Grundlagen Phelan-McDermid-Syndrome (PMDS) Pharmakologische Tests Zusammenfassung Schlussfolgerungen Methoden Elektrophysiologische
Autismus autism spectrum disorders Index Was ist Autismus? Wissenschaftliche Grundlagen Phelan-McDermid-Syndrome (PMDS) Pharmakologische Tests Zusammenfassung Schlussfolgerungen Methoden Elektrophysiologische
Entwicklungs /gewebespezifische Genexpression. Coexpression funktional überlappender Gene
 Übung 11 Genregulation bei Prokaryoten Konzepte: Entwicklungs /gewebespezifische Genexpression Coexpression funktional überlappender Gene Positive Genregulation Negative Genregulation cis /trans Regulation
Übung 11 Genregulation bei Prokaryoten Konzepte: Entwicklungs /gewebespezifische Genexpression Coexpression funktional überlappender Gene Positive Genregulation Negative Genregulation cis /trans Regulation
EINFLUSS DER ZELLMEMBRAN UND EINIGER ZELLAKTOREN AUF DEN ALTERUNGSPROZESS
 EINFLUSS DER ZELLMEMBRAN UND EINIGER ZELLAKTOREN AUF DEN ALTERUNGSPROZESS Der Kern steuert die Proteinsynthese durch Synthese der Boten-RNA (mrna) gemäß den von der DNA gelieferten Informationen Die mrna
EINFLUSS DER ZELLMEMBRAN UND EINIGER ZELLAKTOREN AUF DEN ALTERUNGSPROZESS Der Kern steuert die Proteinsynthese durch Synthese der Boten-RNA (mrna) gemäß den von der DNA gelieferten Informationen Die mrna
Untersuchungen zur differenziellen Genexpression im ZNS von Noradrenalintransporter-Knockout- und Wildtyp-Mäusen
 Untersuchungen zur differenziellen Genexpression im ZNS von Noradrenalintransporter-Knockout- und Wildtyp-Mäusen Inaugural-Dissertation zur Erlangung des Doktorgrades (Dr. rer. nat.) der Mathematisch-Naturwissenschaftlichen
Untersuchungen zur differenziellen Genexpression im ZNS von Noradrenalintransporter-Knockout- und Wildtyp-Mäusen Inaugural-Dissertation zur Erlangung des Doktorgrades (Dr. rer. nat.) der Mathematisch-Naturwissenschaftlichen
Die Entwicklung von B Lymphozyten
 Die Entwicklung von B Lymphozyten Die Entwicklung von B Lymphozyten Übersicht Gliederung I II III IV Stammzellen im Knochenmark VDJ Rekombination Positive Selektion Negative Selektion I Stammzellen im
Die Entwicklung von B Lymphozyten Die Entwicklung von B Lymphozyten Übersicht Gliederung I II III IV Stammzellen im Knochenmark VDJ Rekombination Positive Selektion Negative Selektion I Stammzellen im
Doppelmarkierung im CISH Her2 und Topo2A
 Doppelmarkierung im CISH Her2 und Topo2A Sonja Urdl und Sigurd Lax Institut für Pathologie LKH Graz West sonja.urdl@lkh-grazwest.at; sigurd.lax@lkh-grazwest.at In Situ Hybridisierung (ISH) Molekularbiologische
Doppelmarkierung im CISH Her2 und Topo2A Sonja Urdl und Sigurd Lax Institut für Pathologie LKH Graz West sonja.urdl@lkh-grazwest.at; sigurd.lax@lkh-grazwest.at In Situ Hybridisierung (ISH) Molekularbiologische
6. Induktion der Galactosidase von Escherichia coli
 Johannes Gutenberg-Universität Mainz Institut für Mikrobiologie und Weinforschung FI-Übung: Identifizierung, Wachstum und Regulation (WS 2004/05) Sebastian Lux Datum: 19.1.2005 6. Induktion der Galactosidase
Johannes Gutenberg-Universität Mainz Institut für Mikrobiologie und Weinforschung FI-Übung: Identifizierung, Wachstum und Regulation (WS 2004/05) Sebastian Lux Datum: 19.1.2005 6. Induktion der Galactosidase
Einfluss von Hypoxie auf die Interaktion von Leukämieblasten mit Knochenmark-Stromazellen
 Dr. rer. nat. Jasmin Wellbrock und Prof. Dr. Walter Fiedler Projektbericht an die Alfred & Angelika Gutermuth Stiftung Einfluss von Hypoxie auf die Interaktion von Leukämieblasten mit Knochenmark-Stromazellen
Dr. rer. nat. Jasmin Wellbrock und Prof. Dr. Walter Fiedler Projektbericht an die Alfred & Angelika Gutermuth Stiftung Einfluss von Hypoxie auf die Interaktion von Leukämieblasten mit Knochenmark-Stromazellen
4.1 Größe und Zusammensetzung der Toc-Komplexe in Arabidopsis thaliana
 4.1 Größe und Zusammensetzung der Toc-Komplexe in Arabidopsis thaliana Der aus Erbse isolierte Toc-Kernkomplex besteht aus den zwei Rezeptorproteinen Toc159 und Toc34, sowie der Translokationspore Toc75
4.1 Größe und Zusammensetzung der Toc-Komplexe in Arabidopsis thaliana Der aus Erbse isolierte Toc-Kernkomplex besteht aus den zwei Rezeptorproteinen Toc159 und Toc34, sowie der Translokationspore Toc75
Angewandte Zellbiologie
 Angewandte Zellbiologie Bt-BZ01: Pflanzenzellen als Bioreaktoren 8 CP (VL + Pr) MENDEL (VL) HÄNSCH (Pr) Bt-BZ02: Zellbiologie der Tiere I 8 CP (VL + Pr) BUCHBERGER WINTER (VL) VAUTI (Pr) Bt-BZ03: Zellbiologie
Angewandte Zellbiologie Bt-BZ01: Pflanzenzellen als Bioreaktoren 8 CP (VL + Pr) MENDEL (VL) HÄNSCH (Pr) Bt-BZ02: Zellbiologie der Tiere I 8 CP (VL + Pr) BUCHBERGER WINTER (VL) VAUTI (Pr) Bt-BZ03: Zellbiologie
!!! DIE!EPITHELIAL*MESENCHYMALE! TRANSITION!IN!LUNGENKREBSZELLEN!! JESSICA!ROHRBACH,!GYMNASIUM!KÖNIZ5LEBERMATT,KÖNIZ!
 DIEEPITHELIAL*MESENCHYMALE TRANSITIONINLUNGENKREBSZELLEN JESSICAROHRBACH,GYMNASIUMKÖNIZ5LEBERMATT,KÖNIZ CAMILLAGALANTE,BÜNDNERKANTONSSCHULE,CHUR SUPERVISION:SVENJAGROENEVELD,EPFL Die epithelial- mesenchymale
DIEEPITHELIAL*MESENCHYMALE TRANSITIONINLUNGENKREBSZELLEN JESSICAROHRBACH,GYMNASIUMKÖNIZ5LEBERMATT,KÖNIZ CAMILLAGALANTE,BÜNDNERKANTONSSCHULE,CHUR SUPERVISION:SVENJAGROENEVELD,EPFL Die epithelial- mesenchymale
Das Paper von heute. Samuel Grimm & Jan Kemna
 Das Paper von heute Samuel Grimm & Jan Kemna Bisheriges Modell Was bereits bekannt war - TIR1 ist an Auxinantwort (Zellteilung, Elongation, Differenzierung) beteiligt, im selben Signalweg wie AXR1 - TIR1
Das Paper von heute Samuel Grimm & Jan Kemna Bisheriges Modell Was bereits bekannt war - TIR1 ist an Auxinantwort (Zellteilung, Elongation, Differenzierung) beteiligt, im selben Signalweg wie AXR1 - TIR1
A. Sak, S. Grehl, M. Engelhard, A. Wierlemann, HP. Kaelberlah, P. Erichsen, C. Pöttgen, M. Groneberg, M. Stuschke
 γ-h2ax Foci Induktion in Lymphocten des peripheren Bluts von Tumorpatienten: in vivo und in vitro Interaktion von Cisplatin mit Doppelstrangbruch-Signalen ionisierender Strahlen A. Sak, S. Grehl, M. Engelhard,
γ-h2ax Foci Induktion in Lymphocten des peripheren Bluts von Tumorpatienten: in vivo und in vitro Interaktion von Cisplatin mit Doppelstrangbruch-Signalen ionisierender Strahlen A. Sak, S. Grehl, M. Engelhard,
Das Zytoskelett. Struktur der Intermediärfilamente. Polymerisation/ Depolymerisation. Einteilung der Intermediärfilamente
 Das Zytoskelett Struktur der Intermediärfilamente Polymerisation/ Depolymerisation Einteilung der Intermediärfilamente Zell-Zell- und Zell-Matrix-Verbindungen Einteilung der Zytoskelettsysteme Aktin-Mikrofilamente
Das Zytoskelett Struktur der Intermediärfilamente Polymerisation/ Depolymerisation Einteilung der Intermediärfilamente Zell-Zell- und Zell-Matrix-Verbindungen Einteilung der Zytoskelettsysteme Aktin-Mikrofilamente
1. Einleitung S. 1 1.1. Zelladhäsionsmoleküle S. 1 1.2. Die Immunglobulin-Superfamilie S. 2. 1.2.1. Das neurale Zelladhäsionsmolekül NCAM S.
 Inhaltsverzeichnis 1. Einleitung S. 1 1.1. Zelladhäsionsmoleküle S. 1 1.2. Die Immunglobulin-Superfamilie S. 2 1.2.1. Das neurale Zelladhäsionsmolekül NCAM S. 4 1.2.1.1. Molekulare Struktur von NCAM S.
Inhaltsverzeichnis 1. Einleitung S. 1 1.1. Zelladhäsionsmoleküle S. 1 1.2. Die Immunglobulin-Superfamilie S. 2 1.2.1. Das neurale Zelladhäsionsmolekül NCAM S. 4 1.2.1.1. Molekulare Struktur von NCAM S.
Die Embryonalentwicklung: Schlüssel zum Verständnis von Stammzellen
 Modul 1-2 Die Embryonalentwicklung: Schlüssel zum Verständnis von Stammzellen Wenn aus einer befruchteten Eizelle (Zygote) durch Zellteilungen viele Zellen entstehen, die sich reorganisieren, spezialisieren,
Modul 1-2 Die Embryonalentwicklung: Schlüssel zum Verständnis von Stammzellen Wenn aus einer befruchteten Eizelle (Zygote) durch Zellteilungen viele Zellen entstehen, die sich reorganisieren, spezialisieren,
Inhaltsverzeichnis. 1. Einleitung... 9. 2. Material und Methoden... 32
 Inhaltsverzeichnis 1. Einleitung... 9 1.1 Posttranslationale Modifikationen... 10 1.2 N-Glykosylierung... 11 1.3 O-Glykosylierung... 12 1.4 Veränderungen der Glykosylierung bei der Tumorentstehung... 13
Inhaltsverzeichnis 1. Einleitung... 9 1.1 Posttranslationale Modifikationen... 10 1.2 N-Glykosylierung... 11 1.3 O-Glykosylierung... 12 1.4 Veränderungen der Glykosylierung bei der Tumorentstehung... 13
Phylogenetisches Praktikum im ITZ 08.02.2010-19.02.2010
 Phylogenetisches Praktikum im ITZ 08.02.2010-19.02.2010 Zellzyklus- und neuronale Gene in Trichoplax adhaerens S. Sagasser Praktikanten: Patrick Reinke und Jan Kleveman Betreuerin: Karolin von der Chevallerie
Phylogenetisches Praktikum im ITZ 08.02.2010-19.02.2010 Zellzyklus- und neuronale Gene in Trichoplax adhaerens S. Sagasser Praktikanten: Patrick Reinke und Jan Kleveman Betreuerin: Karolin von der Chevallerie
4.5. Ergebnisse der Expression und Aufreinigung von rekombinanten Proteinen aus Escherichia coli
 4.5. Ergebnisse der Expression und Aufreinigung von rekombinanten Proteinen aus Escherichia coli Für die Auswertung des ELISAs wurden Positivkontrollen benötigt. Dazu wurde einem Teil der Tiere subcutan
4.5. Ergebnisse der Expression und Aufreinigung von rekombinanten Proteinen aus Escherichia coli Für die Auswertung des ELISAs wurden Positivkontrollen benötigt. Dazu wurde einem Teil der Tiere subcutan
Protokoll Praktikum für Humanbiologie Studenten
 Protokoll Praktikum für Humanbiologie Studenten Teil A: Charakterisierung der Auswirkungen von γ Interferon auf die Protein und mrna Mengen in humanen A549 Lungenepithelzellen. Studentenaufgaben Tag 1
Protokoll Praktikum für Humanbiologie Studenten Teil A: Charakterisierung der Auswirkungen von γ Interferon auf die Protein und mrna Mengen in humanen A549 Lungenepithelzellen. Studentenaufgaben Tag 1
Psoriasin is a major Escherichia coli-cidal factor of the female genital tract
 Psoriasin is a major Escherichia coli-cidal factor of the female genital tract Mildner M, Stichenwirth M, Abtin A, Eckhart L, Sam C, Gläser R, Schröder JM, Gmeiner R, Mlitz V, Pammer J, Geusau A, Tschachler
Psoriasin is a major Escherichia coli-cidal factor of the female genital tract Mildner M, Stichenwirth M, Abtin A, Eckhart L, Sam C, Gläser R, Schröder JM, Gmeiner R, Mlitz V, Pammer J, Geusau A, Tschachler
Zelldifferenzierung und Morphogenese
 Zelldifferenzierung und Morphogenese Fragestellung: Wie ist es möglich, daß sich aus einer einzigen Zelle ein hochkomplexes Lebewesen mit hohem Differenzierungsgrad entwickeln kann? weniger gut verstanden
Zelldifferenzierung und Morphogenese Fragestellung: Wie ist es möglich, daß sich aus einer einzigen Zelle ein hochkomplexes Lebewesen mit hohem Differenzierungsgrad entwickeln kann? weniger gut verstanden
APP-GFP/Fluoreszenzmikroskop. Aufnahmen neuronaler Zellen, mit freund. Genehmigung von Prof. Stefan Kins, TU Kaiserslautern
 Über die Herkunft von Aβ42 und Amyloid-Plaques Heute ist sicher belegt, dass sich die amyloiden Plaques aus einer Vielzahl an Abbaufragmenten des Amyloid-Vorläufer-Proteins (amyloid-precursor-protein,
Über die Herkunft von Aβ42 und Amyloid-Plaques Heute ist sicher belegt, dass sich die amyloiden Plaques aus einer Vielzahl an Abbaufragmenten des Amyloid-Vorläufer-Proteins (amyloid-precursor-protein,
Inhaltsverzeichnis. 1 Einleitung... 1
 Inhaltsverzeichnis 1 Einleitung... 1 1.1 G-Protein-gekoppelte Rezeptoren... 3 1.1.1 Klassifizierung und Struktur von G-Protein-gekoppelten Rezeptoren... 6 1.1.2 Purinerge Rezeptoren... 12 1.1.3 Pharmazeutische
Inhaltsverzeichnis 1 Einleitung... 1 1.1 G-Protein-gekoppelte Rezeptoren... 3 1.1.1 Klassifizierung und Struktur von G-Protein-gekoppelten Rezeptoren... 6 1.1.2 Purinerge Rezeptoren... 12 1.1.3 Pharmazeutische
NFATc2 ist ein Schalter der T-Zell-Rezeptor-abhängigen Aktivierung in humanen CD4 + Th-Zellen
 NFATc2 ist ein Schalter der T-Zell-Rezeptor-abhängigen Aktivierung in humanen CD4 + Th-Zellen Dissertation zur Erlangung des akademischen Grades des Doktors der Naturwissenschaften (Dr. rer. nat) eingereicht
NFATc2 ist ein Schalter der T-Zell-Rezeptor-abhängigen Aktivierung in humanen CD4 + Th-Zellen Dissertation zur Erlangung des akademischen Grades des Doktors der Naturwissenschaften (Dr. rer. nat) eingereicht
Entdeckungen unter der Schädeldecke. Jean-Marc Fritschy Institut für Pharmakologie und Toxikologie
 Entdeckungen unter der Schädeldecke Jean-Marc Fritschy Institut für Pharmakologie und Toxikologie Inhalt 1. GFP, das Wunderprotein 2. Die Nervenzellen bei der Arbeit beobachten 3. Nervenzellen mit Licht
Entdeckungen unter der Schädeldecke Jean-Marc Fritschy Institut für Pharmakologie und Toxikologie Inhalt 1. GFP, das Wunderprotein 2. Die Nervenzellen bei der Arbeit beobachten 3. Nervenzellen mit Licht
HER2-Diagnostik. Ein Leitfaden für Brustkrebs-Patientinnen
 HER2-Diagnostik Ein Leitfaden für Brustkrebs-Patientinnen Was ist HER2? HER2 vielfach auch als erbb2 oder HER2/neu bezeichnet ist ein Eiweiß bzw. Proteinbaustein an der Oberfläche von Zellen (Rezeptor).
HER2-Diagnostik Ein Leitfaden für Brustkrebs-Patientinnen Was ist HER2? HER2 vielfach auch als erbb2 oder HER2/neu bezeichnet ist ein Eiweiß bzw. Proteinbaustein an der Oberfläche von Zellen (Rezeptor).
RESOLUTION OIV/OENO 427/2010 KRITERIEN FÜR METHODEN ZUR QUANTIFIZIERUNG VON POTENTIELL ALLERGENEN RÜCKSTÄNDEN EIWEISSHALTIGER SCHÖNUNGSMITTEL IM WEIN
 RESOLUTION OIV/OENO 427/2010 KRITERIEN FÜR METHODEN ZUR QUANTIFIZIERUNG VON POTENTIELL ALLERGENEN RÜCKSTÄNDEN EIWEISSHALTIGER SCHÖNUNGSMITTEL IM WEIN Die GENERALVERSAMMLUNG, unter Berücksichtigung des
RESOLUTION OIV/OENO 427/2010 KRITERIEN FÜR METHODEN ZUR QUANTIFIZIERUNG VON POTENTIELL ALLERGENEN RÜCKSTÄNDEN EIWEISSHALTIGER SCHÖNUNGSMITTEL IM WEIN Die GENERALVERSAMMLUNG, unter Berücksichtigung des
Zellbiologie: BSc Arbeiten 15/16
 Zellbiologie: BSc Arbeiten 15/16 Lehrstuhl Zellbiologie: Arbeitsgruppen Prof. Benedikt Kost Prof. Georg Kreimer PD Michael Lebert Slot Zeitraum Anzahl Plätze Semester 1 24.08.15 09.10.15 4 Ferien 2 09.11.15
Zellbiologie: BSc Arbeiten 15/16 Lehrstuhl Zellbiologie: Arbeitsgruppen Prof. Benedikt Kost Prof. Georg Kreimer PD Michael Lebert Slot Zeitraum Anzahl Plätze Semester 1 24.08.15 09.10.15 4 Ferien 2 09.11.15
Western Blot. Zuerst wird ein Proteingemisch mit Hilfe einer Gel-Elektrophorese aufgetrennt.
 Western Blot Der Western Blot ist eine analytische Methode zum Nachweis bestimmter Proteine in einer Probe. Der Nachweis erfolgt mit spezifischen Antikörpern, die das gesuchte Protein erkennen und daran
Western Blot Der Western Blot ist eine analytische Methode zum Nachweis bestimmter Proteine in einer Probe. Der Nachweis erfolgt mit spezifischen Antikörpern, die das gesuchte Protein erkennen und daran
4. Quantitative Analyse der Ligand-Bindungsstudien
 4. Quantitative Analyse der Ligand-Bindungsstudien Im folgenden apitel sollen die grundlegenden analytischen Methoden zur Interpretation der experimentell gewonnenen Bindungsdaten vorgestellt werden. ie
4. Quantitative Analyse der Ligand-Bindungsstudien Im folgenden apitel sollen die grundlegenden analytischen Methoden zur Interpretation der experimentell gewonnenen Bindungsdaten vorgestellt werden. ie
1 Einleitung. 1 Einleitung. 1.1 Zielsetzung
 1.1 Zielsetzung 1 Einleitung Embryonale Zellen zeichnen sich durch spezifische Eigenschaften bei Wachstum, Differenzierung und Stoffwechsel aus. Untersuchungen an adulten Zellen oder vordifferenzierten
1.1 Zielsetzung 1 Einleitung Embryonale Zellen zeichnen sich durch spezifische Eigenschaften bei Wachstum, Differenzierung und Stoffwechsel aus. Untersuchungen an adulten Zellen oder vordifferenzierten
Somatischer Kerntransfer und zelluläre Therapien: Möglichkeiten zum Gendoping im Spitzensport
 39 Somatischer Kerntransfer und zelluläre Therapien: Möglichkeiten zum Gendoping im Spitzensport Heiner Niemann Der Begriff Klon stammt aus dem griechischen klonos und bedeutet Ast oder Zweig. Heute wird
39 Somatischer Kerntransfer und zelluläre Therapien: Möglichkeiten zum Gendoping im Spitzensport Heiner Niemann Der Begriff Klon stammt aus dem griechischen klonos und bedeutet Ast oder Zweig. Heute wird
Immunologische Techniken. Gelpräzipitationsreaktionen 1. Immundiffusion 2. Immunelektrophorese
 Immunologische Techniken Gelpräzipitationsreaktionen 1. Immundiffusion 2. Immunelektrophorese Direkte und indirekte Hämagglutination Direkte Hämagglutination: wenig empfindlich Indirekte Hämagglutination:
Immunologische Techniken Gelpräzipitationsreaktionen 1. Immundiffusion 2. Immunelektrophorese Direkte und indirekte Hämagglutination Direkte Hämagglutination: wenig empfindlich Indirekte Hämagglutination:
Bestimmung der Phenolaseaktivität in Kartoffelpreßsaft
 Paul Elsinghorst, Jürgen Gäb Ergebnisprotokoll Versuch 22 Bestimmung der Phenolaseaktivität in Kartoffelpreßsaft 1. Stichworte Phenolasen (Monophenolasen, Diphenolasen, natürliche Bedeutung) Messmethoden
Paul Elsinghorst, Jürgen Gäb Ergebnisprotokoll Versuch 22 Bestimmung der Phenolaseaktivität in Kartoffelpreßsaft 1. Stichworte Phenolasen (Monophenolasen, Diphenolasen, natürliche Bedeutung) Messmethoden
Molekulare Mechanismen der Signaltransduktion. 06 - Kartierung des AXR1 Gens + early auxin-induced genes Folien: http://tinyurl.
 Molekulare Mechanismen der Signaltransduktion 06 - Kartierung des AXR1 Gens + early auxin-induced genes Folien: http://tinyurl.com/modul-mms bisheriges Modell auxin auxin AXR1 auxin response AXR1 potentieller
Molekulare Mechanismen der Signaltransduktion 06 - Kartierung des AXR1 Gens + early auxin-induced genes Folien: http://tinyurl.com/modul-mms bisheriges Modell auxin auxin AXR1 auxin response AXR1 potentieller
Das Reagenz speziell für die Transfektion von Säugerzellen mit sirna und mirna
 METAFECTENE SI + Das Reagenz speziell für die Transfektion von Säugerzellen mit sirna und mirna Bestellinformationen, MSDS, Publikationen und Anwendungsbeispiele unter www.biontex.com Produkt Katalognummer
METAFECTENE SI + Das Reagenz speziell für die Transfektion von Säugerzellen mit sirna und mirna Bestellinformationen, MSDS, Publikationen und Anwendungsbeispiele unter www.biontex.com Produkt Katalognummer
3.1 Die Kultur normaler humaner Bronchialepithelzellen. 3.1.2 Die selektive Gewinnung und Kultivierung normaler humaner Bronchialepithelzellen
 3 Ergebnisse 3.1 Die Kultur normaler humaner Bronchialepithelzellen 3.1.1 Übersicht der gewonnenen Resektate Von 3 Patienten, die sich ab Juni 1997 bis Juni 1999 einem thoraxchirurgischen Eingriff in der
3 Ergebnisse 3.1 Die Kultur normaler humaner Bronchialepithelzellen 3.1.1 Übersicht der gewonnenen Resektate Von 3 Patienten, die sich ab Juni 1997 bis Juni 1999 einem thoraxchirurgischen Eingriff in der
Regulation der Genexpression: regulierbare Promotoren, Proteine und sirna
 Regulation der Genexpression: regulierbare Promotoren, Proteine und sirna Biochemie Praktikum Christian Brendel, AG Grez Ebenen der Genregulation in Eukaryoten Cytoplasma DNA Zellkern Introns Exons Chromatin
Regulation der Genexpression: regulierbare Promotoren, Proteine und sirna Biochemie Praktikum Christian Brendel, AG Grez Ebenen der Genregulation in Eukaryoten Cytoplasma DNA Zellkern Introns Exons Chromatin
Grundlegende Methoden der Molekularen Biologie, AD I. Prof. Dr. Albert Duschl
 Grundlegende Methoden der Molekularen Biologie, AD I Prof. Dr. Albert Duschl Molekulare Mechanismen Themen für die erste Vorlesung: Signalwege, am Beispiel von Jak/STAT Immunpräzipitation, Immunfluoreszenz
Grundlegende Methoden der Molekularen Biologie, AD I Prof. Dr. Albert Duschl Molekulare Mechanismen Themen für die erste Vorlesung: Signalwege, am Beispiel von Jak/STAT Immunpräzipitation, Immunfluoreszenz
Heterologe Expression einer funktionellen Domäne des nikotinischen Acetylcholinrezeptors
 Heterologe Expression einer funktionellen Domäne des nikotinischen Acetylcholinrezeptors Inauguraldissertation zur Erlangung der Doktorwürde am Fachbereich Biologie/Chemie/Pharmazie der Freien Universität
Heterologe Expression einer funktionellen Domäne des nikotinischen Acetylcholinrezeptors Inauguraldissertation zur Erlangung der Doktorwürde am Fachbereich Biologie/Chemie/Pharmazie der Freien Universität
Pathohysiologie und Androgendeprivationtstherapie(ADT) des kastrationsresistenten Prostatakarzinoms
 des kastrationsresistenten Prostatakarzinoms") Pathohysiologie und Androgendeprivationtstherapie(ADT) des kastrationsresistenten Prostatakarzinoms KASTRATIONSRESISTENTES PROSTATAKARZINOM (CRPC) Patienten sprechen im Allgemeinen gut auf eine ADT an.
Pathohysiologie und Androgendeprivationtstherapie(ADT) des kastrationsresistenten Prostatakarzinoms KASTRATIONSRESISTENTES PROSTATAKARZINOM (CRPC) Patienten sprechen im Allgemeinen gut auf eine ADT an.
Gentherapie. Grüne Gentechnologie
 Gentherapie Grüne Gentechnologie Definition Mit Gentherapie bezeichnet man das Einfügen von Genen in Zellen eines Individuums zur Behandlung von Erbkrankheiten bzw. Gendefekten Durch die Einführung soll
Gentherapie Grüne Gentechnologie Definition Mit Gentherapie bezeichnet man das Einfügen von Genen in Zellen eines Individuums zur Behandlung von Erbkrankheiten bzw. Gendefekten Durch die Einführung soll
Familienanamnese, Tumorlokalisation, Tumorabrenzung, Tumorgröße, TNM-Stadien
 Prognosefaktoren beim Die Prognose dieser Studie untersucht die Dauer des rezidivfreien Intervalls und die Gesamtüberlebenszeit. Folgende Prognosefaktoren wurden berücksichtigt: 1.klinische Daten 2. histopathologischer
Prognosefaktoren beim Die Prognose dieser Studie untersucht die Dauer des rezidivfreien Intervalls und die Gesamtüberlebenszeit. Folgende Prognosefaktoren wurden berücksichtigt: 1.klinische Daten 2. histopathologischer
B-Zellentwicklung. Grundlagen der Immunologie 5. Semester - Dienstags Uhr Ruhr-Universität Bochum, HMA 20 HEV.
 Grundlagen der Immunologie 5. Semester - Dienstags 11.15 Uhr Ruhr-Universität Bochum, HMA 20 B-Zellentwicklung Albrecht Bufe www.ruhr-uni-bochum.de/homeexpneu T und B Zellen zirkulieren unablässig durch
Grundlagen der Immunologie 5. Semester - Dienstags 11.15 Uhr Ruhr-Universität Bochum, HMA 20 B-Zellentwicklung Albrecht Bufe www.ruhr-uni-bochum.de/homeexpneu T und B Zellen zirkulieren unablässig durch
SCRIPTUM HIGH PRECISE
 Nur für Forschungszwecke Datenblatt Artikel BS.50.020 = 20 Reaktionen x 50 µl Artikel BS.50.100 = 100 Reaktionen x 50 µl Artikel BS.50. 500 = 500 Reaktionen x 50 µl Haltbarkeit: 12 Monate Lagerung bei
Nur für Forschungszwecke Datenblatt Artikel BS.50.020 = 20 Reaktionen x 50 µl Artikel BS.50.100 = 100 Reaktionen x 50 µl Artikel BS.50. 500 = 500 Reaktionen x 50 µl Haltbarkeit: 12 Monate Lagerung bei
"Stammzellen - Alleskönner zwischen Wissenschaft und Ethik"
 "Stammzellen - Alleskönner zwischen Wissenschaft und Ethik" Jörg Klug Justus-Liebig-Universität Gießen 2. Science Bridge Seminar 19. November 2010 Wo kommt der Name her? A. megalocephala C. elegans Edmund
"Stammzellen - Alleskönner zwischen Wissenschaft und Ethik" Jörg Klug Justus-Liebig-Universität Gießen 2. Science Bridge Seminar 19. November 2010 Wo kommt der Name her? A. megalocephala C. elegans Edmund
Aus dem Alltag in der Molekularbiologie. Dr. Andreas Bergthaler
 Aus dem Alltag in der Molekularbiologie Schulvortrag, 11.1.2010 A3rak4ve Gründe für eine Karriere Im Op4malfall: in der Forschung kann man selbst bes,mmen, was man erforscht. arbeitet man mit schlauen
Aus dem Alltag in der Molekularbiologie Schulvortrag, 11.1.2010 A3rak4ve Gründe für eine Karriere Im Op4malfall: in der Forschung kann man selbst bes,mmen, was man erforscht. arbeitet man mit schlauen
Immunofluoreszenz-Markierung an kultivierten adhärenten Säugerzellen. Methanol-Fixierung
 Immunofluoreszenz-Markierung an kultivierten adhärenten Säugerzellen Methanol-Fixierung 2 Materialien Pinzetten Für das Handling der Zellen ist es empfehlenswert Pinzetten mit sehr feiner Spitze zu nutzen.
Immunofluoreszenz-Markierung an kultivierten adhärenten Säugerzellen Methanol-Fixierung 2 Materialien Pinzetten Für das Handling der Zellen ist es empfehlenswert Pinzetten mit sehr feiner Spitze zu nutzen.
Versuch 6.14 ph-abhängigkeit eines Indikators am Beispiel Thymolblau
 Versuch 6.14 ph-abhängigkeit eines Indikators am Beispiel Thymolblau Einleitung Lösungen mit verschiedenen ph-werten von stark sauer bis stark basisch werden mit gleich viel Thymolblau-Lösung versetzt.
Versuch 6.14 ph-abhängigkeit eines Indikators am Beispiel Thymolblau Einleitung Lösungen mit verschiedenen ph-werten von stark sauer bis stark basisch werden mit gleich viel Thymolblau-Lösung versetzt.
3. EIGENE UNTERSUCHUNGEN
 3.2. Ergebnisse 3.2.1. Probenentnahme Von 100 Rotfüchsen waren 78 Tiere mit Schildzecken und 41 mit Flöhen befallen. Bei 15 Füchsen konnte weder ein Zecken- noch ein Flohbefall festgestellt werden. Beim
3.2. Ergebnisse 3.2.1. Probenentnahme Von 100 Rotfüchsen waren 78 Tiere mit Schildzecken und 41 mit Flöhen befallen. Bei 15 Füchsen konnte weder ein Zecken- noch ein Flohbefall festgestellt werden. Beim
Etablierung, Validierung und praktische Anwendung einer multiplex real-time RT-PCR zum Nachweis des Rabbit Haemorrhagic Disease Virus
 Aus dem Institut für Virologie der Tierärztlichen Hochschule Hannover und dem Institut für Virusdiagnostik des Friedrich-Loeffler-Instituts, Insel Riems Etablierung, Validierung und praktische Anwendung
Aus dem Institut für Virologie der Tierärztlichen Hochschule Hannover und dem Institut für Virusdiagnostik des Friedrich-Loeffler-Instituts, Insel Riems Etablierung, Validierung und praktische Anwendung
2. Bachelorarbeit. Bachelorstudiengang Molekulare Biotechnologie Fachhochschule Campus Wien
 Etablierung eines zellulären in vitro Assays zur Bestimmung der Angiotensin II Aktivität basierend auf der Expression von Luziferase durch die Aktivierung von NF-κB 2. Bachelorarbeit Bachelorstudiengang
Etablierung eines zellulären in vitro Assays zur Bestimmung der Angiotensin II Aktivität basierend auf der Expression von Luziferase durch die Aktivierung von NF-κB 2. Bachelorarbeit Bachelorstudiengang
Biologie für Mediziner
 Biologie für Mediziner - Zellbiologie 1 - Prof. Dr. Reiner Peters Institut für Medizinische Physik und Biophysik/CeNTech Robert-Koch-Strasse 31 Tel. 0251-835 6933, petersr@uni-muenster.de Dr. Martin Kahms
Biologie für Mediziner - Zellbiologie 1 - Prof. Dr. Reiner Peters Institut für Medizinische Physik und Biophysik/CeNTech Robert-Koch-Strasse 31 Tel. 0251-835 6933, petersr@uni-muenster.de Dr. Martin Kahms
ENZYME. Teil 1: Grundlagen und Substratbestimmungen
 ENZYME Teil 1: Grundlagen und Substratbestimmungen Metastabiler Zustand Beispiel: Glucose-6-Phosphat + H 2 O [Glc6P] [H 2 0] K = = 1.135 x 10 [Glc] [Pi] -3 Gleichgewicht stark auf Seite von Glc + Pi Glucose
ENZYME Teil 1: Grundlagen und Substratbestimmungen Metastabiler Zustand Beispiel: Glucose-6-Phosphat + H 2 O [Glc6P] [H 2 0] K = = 1.135 x 10 [Glc] [Pi] -3 Gleichgewicht stark auf Seite von Glc + Pi Glucose
Wirkungen auf die Blut-Hirn-Schranke, DNA-Schädigung
 Wirkungen auf die Blut-Hirn-Schranke, DNA-Schädigung Dr. Monika Asmuß Bundesamt für Strahlenschutz Mobilfunk und Gesundheit BfS-Informationsveranstaltung, 25. Juni 2009, München 1 Blut-Hirn-Schranke (BHS)
Wirkungen auf die Blut-Hirn-Schranke, DNA-Schädigung Dr. Monika Asmuß Bundesamt für Strahlenschutz Mobilfunk und Gesundheit BfS-Informationsveranstaltung, 25. Juni 2009, München 1 Blut-Hirn-Schranke (BHS)
Melanopsin und die innere Uhr
 Hintergrund Melanopsin und die innere Uhr Benjamin Schuster-Böcker, Betreuer: Dr. Rita Rosenthal Zirkadianer Rhythmus Praktisch alle höheren Lebewesen folgen einem Aktivitätsrhythmus, der dem Wechsel von
Hintergrund Melanopsin und die innere Uhr Benjamin Schuster-Böcker, Betreuer: Dr. Rita Rosenthal Zirkadianer Rhythmus Praktisch alle höheren Lebewesen folgen einem Aktivitätsrhythmus, der dem Wechsel von
Entwicklungsbiologie Master-Module. (bzw. 9 + 3)
") Entwicklungsbiologie Master-Module (bzw. 9 + 3) Entwicklungsbiologie Master-Module Block 3 Ebio I Evolution von Musterbildung und Gametogenese Mikroinjektion von Tribolium-Embryonen zur Proteinlokalisierung
Entwicklungsbiologie Master-Module (bzw. 9 + 3) Entwicklungsbiologie Master-Module Block 3 Ebio I Evolution von Musterbildung und Gametogenese Mikroinjektion von Tribolium-Embryonen zur Proteinlokalisierung
Praktikum Physik. Protokoll zum Versuch: Oberflächenspannung. Durchgeführt am Gruppe X
 Praktikum Physik Protokoll zum Versuch: Oberflächenspannung Durchgeführt am 02.02.2012 Gruppe X Name 1 und Name 2 (abc.xyz@uni-ulm.de) (abc.xyz@uni-ulm.de) Betreuer: Wir bestätigen hiermit, dass wir das
Praktikum Physik Protokoll zum Versuch: Oberflächenspannung Durchgeführt am 02.02.2012 Gruppe X Name 1 und Name 2 (abc.xyz@uni-ulm.de) (abc.xyz@uni-ulm.de) Betreuer: Wir bestätigen hiermit, dass wir das
Musterlösung - Übung 5 Vorlesung Bio-Engineering Sommersemester 2008
 Aufgabe 1: Prinzipieller Ablauf der Proteinbiosynthese a) Erklären Sie folgende Begriffe möglichst in Ihren eigenen Worten (1 kurzer Satz): Gen Nukleotid RNA-Polymerase Promotor Codon Anti-Codon Stop-Codon
Aufgabe 1: Prinzipieller Ablauf der Proteinbiosynthese a) Erklären Sie folgende Begriffe möglichst in Ihren eigenen Worten (1 kurzer Satz): Gen Nukleotid RNA-Polymerase Promotor Codon Anti-Codon Stop-Codon
Allgemeine Produktbeschreibung Spezifisches IgE
 Allgemeine Produktbeschreibung Spezifisches IgE Produkt- und Verfahrensbeschreibung Der Nachweis von spezifischem IgE in Serum oder Plasma basiert auf dem Prinzip eines Enzymimmunoassays zur quantitativen
Allgemeine Produktbeschreibung Spezifisches IgE Produkt- und Verfahrensbeschreibung Der Nachweis von spezifischem IgE in Serum oder Plasma basiert auf dem Prinzip eines Enzymimmunoassays zur quantitativen
1.3 Begründe kurz, ob mit Hilfe der fünf in der vereinfachten ID-Card verwendeten Marker aus Aufgabe 1.1 das Geschlecht mit bestimmt werden kann.
 Übungsaufgaben zum DNA-Fingerprinter 1/6 1. In der EU wird die Einführung einer ID-Card (Nachfolger des Personalausweises, Entwurf s. Abb. 1) diskutiert, die neben Foto, äußeren Merkmalen und Fingerabdruck
Übungsaufgaben zum DNA-Fingerprinter 1/6 1. In der EU wird die Einführung einer ID-Card (Nachfolger des Personalausweises, Entwurf s. Abb. 1) diskutiert, die neben Foto, äußeren Merkmalen und Fingerabdruck
Wissenschaftliche Angestellte, Laborleitung Geburtsdatum, Nationalität: 28.04.1973, deutsch
 Forschungsbericht über das Projekt Einfluss der TLR3-Aktivierung des retinalen Pigmentepithels auf das Verhalten von Makrophagen, gefördert durch die DOG im Rahmen der Forschungsförderung innovativer wissenschaftlicher
Forschungsbericht über das Projekt Einfluss der TLR3-Aktivierung des retinalen Pigmentepithels auf das Verhalten von Makrophagen, gefördert durch die DOG im Rahmen der Forschungsförderung innovativer wissenschaftlicher