Der Leukozyten Rezeptor Komplex des Bankivahuhns
|
|
|
- Marielies Schubert
- vor 7 Jahren
- Abrufe
Transkript
1 Aus dem Institut für Physiologie, Physiologische Chemie und Tierernährung des Veterinärwissenschaftlichen Departments der Tierärztlichen Fakultät der Ludwig-Maximilians-Universität München Vorstand: Univ.-Prof. Dr. M. Stangassinger Arbeit angefertigt unter der Leitung von Univ.-Prof. Dr. T. Göbel Der Leukozyten Rezeptor Komplex des Bankivahuhns Inaugural-Dissertation zur Erlangung der tiermedizinischen Doktorwürde der Tierärztlichen Fakultät der Ludwig-Maximilians-Universität München von Katharina Lochner aus Peißenberg München 2010
2 Gedruckt mit Genehmigung der Tierärztlichen Fakultät der Ludwig-Maximilians-Universität München Dekan: Univ.-Prof. Dr. Braun Berichterstatter: Univ.-Prof. Dr. Göbel Koreferent/en: Univ.-Prof. Dr. Dr. Erhard Prof. Dr. Kaltner Univ.-Prof. Dr. Matis Priv.-Doz. Dr. Rinder Tag der Promotion: 24. Juli 2010
3 Für meine Mutter
4 Inhaltsverzeichnis Inhaltsverzeichnis 1 Einleitung Literatur Ig-ähnliche Rezeptoren LRC ILT KIR und GPVI LAIR FcαRI (CD89) und NKp LRC des Huhns Publikation Diskussion Zusammenfassung Summary Literaturverzeichnis Persönliche Daten...27 I
5 Abkürzungsverzeichnis Abkürzungsverzeichnis AS Aminosäuren BAC artifizielles Chromosom (engl. Bacterial Artificial Chromosome) CEACAM karzinoembryonisches Antigen-abhängiges Zell-Adhäsionsmolekül (engl. Carcinoembryonic Antigen-related Cell Adhesion Molecule) CHIR Hühner Ig-ähnlicher Rezeptor (engl. Chicken Ig-like Receptor) FcαR Fcα Rezeptor FcεR Fcε Rezeptor GPVI Glycoprotein VI Ig Immunglobulin IgSF Immunglobulin-Superfamilie ILT Ig-ähnliches Transkript ITAM Immunrezeptor Tyrosin-basierendes aktivierendes Motiv ITIM Immunrezeptor Tyrosin-basierendes inhibierendes Motiv KIR Killerzellen Ig-ähnlicher Rezeptor LAIR Leucozyten assoziierter Ig-ähnlicher Rezeptor LILR Leukozyten Ig-ähnlicher Rezeptor LRC Leukozyten Rezeptor Komplex (engl. Leukocyte Receptor Complex) LSL Lohmann Leghorn (engl. Lohmann selected Leghorn) MHC Haupthistokompatibilitätskomplex (engl. Major Histocompatibility Complex) MIR Monozyten/Makrophagen Ig- ähnlicher Rezeptor NITR neuer Immun-Typ Rezeptor NK-Zelle natürliche Killer-Zelle NKC natürlicher Killer Gen-Komplex (engl. Natural Killer Gene-Complex) NKp natürliches Killerprotein PIR gepaarter Ig- ähnlicher Rezeptor (engl. Paired Ig-like Receptor) RJF Bankivahuhn (engl. Red Jungle Fowl) SH2 Src Homologe 2 SHP SH2-beinhaltende Proteintyrosinphosphatase SIGLEC Sialinsäure-bindendes Ig-ähnliches Lektin (engl. Sialic acid binding Iglike Lectins) ITSM Immunrezeptor Tyrosin-basierendes Wechselmotiv (engl. Immune Receptor Tyrosin-based Switch Motif) II
6 Einleitung 1 Einleitung Leukozyten werden durch entgegen gesetzte Signalwege reguliert. Die Interaktion von Oberflächenrezeptoren mit ihren Liganden löst eine Hemmung oder Aktivierung der Zellantwort aus. Zwei große Gruppen strukturell unterschiedlicher Moleküle sind für die Vermittlung dieser Regulation verantwortlich. Dies sind die Mitglieder der C- Typ Lektin-Superfamilie im natürlichen Killer Gen-Komplex (NKC) [1], beim Mensch auf Chromosom 12p13.1 [2], und die der Immunglobulin-Superfamilie (IgSF) im Leukozyten Rezeptor Komplex (LRC) auf Chromosom 19q13.4 [3]. Der LRC des Menschen umfasst 1Mb und beinhaltet 45 verschiedene Gensequenzen, von denen 30 als Immunglobulin (Ig) -ähnliche Rezeptorgene identifiziert wurden [4,5]. Es sind dies die Familien der Ig-ähnliche Transkripte (ILT), Killerzellen Ig-ähnliche Rezeptoren (KIR), und Leukozyten assoziierten Ig-ähnliche Rezeptoren (LAIR) sowie das Glycoprotein VI (GPVI), das natürliche Killerprotein (NKp) 46 und der Fcα Rezeptor (FcαR) I [4]. Am zentromeren Ende des Komplexes werden neue Gene kodiert, die sogenannten LENG, die einen anderen Ursprung und strukturell keinen Bezug zu Ig-ähnlichen Rezeptoren haben [4]. Weiter zentromer des eigentlichen LRC liegen Gene die verwandte Moleküle kodieren. Dieser ganze Bereich wird deshalb als erweiterter (extended) LRC bezeichnet und beinhaltet zusätzliche Ig-ähnliche Genfamilien wie die karzinoembryonischen Antigen-abhängigen Zell-Adhäsionsmoleküle (CEACAM) und die Sialinsäure-bindenden Ig-ähnlichen Lektine (SIGLEC). CEACAM spielen eine Rolle bei verschiedenen Vorgängen, wie Angiogenese, Tumorentwicklung, Insulinregulation und Apoptose. Des weiteren gibt es unter ihnen Rezeptoren für Pathogene wie Neisseria, Haemophilus, Escherichia und Salmonellen sowie den Maus Hepatitis Virus, welche die Bindung an CEACAM zur Transzytose und Immunmodulation verwenden [6,7]. Wie der Name schon sagt, sind SIGLEC Rezeptoren für bestimmte Zuckerreste. Sie kommen als Adhäsionsmoleküle auf verschiedenen Leukozyten vor wie z.b. Siglec-1 (CD169) auf Makrophagen [8] und Siglec-2 (CD22) auf B-Zellen [9]. Die Ig-ähnlichen Rezeptoren des LRC kommen in drei Varianten vor. Entweder sind sie mit Immunrezeptor Tyrosin-basierenden inhibierenden Motiven (ITIM) ausgestattet und wirken somit hemmend auf die Zelle oder haben durch bestimmte Aminosäuren (AS) die Möglichkeit Adaptermoleküle mit Immunrezeptor Tyrosin- 1
7 Einleitung basierenden aktivierenden Motiven (ITAM) [10] zu binden, welche die Zelle aktivieren. Zusätzlich gibt es Rezeptoren, die über hemmende und aktivierende Eigenschaften in Kombination verfügen, was diese potentiell bifunktional macht. Eine zum menschlichen LRC homologe Region liegt bei der Maus auf Chromosom 7, wobei keine den KIR entsprechenden Gene vorhanden sind [4]. Auch in anderen Tierarten konnten homologe Genfamilien nachgewiesen werden. So wurden z.b. in verschiedenen Knochenfischen neue Immun-Typ Rezeptoren (NITR) gefunden [11], die funktionelle Homologe der KIR und ILT sind [12]. Untersuchungen an Ciona intestinalis geben Hinweise darauf, dass der LRC der Wirbeltiere aus einer einzelnen Vorläuferregion entstanden ist, die sich nach zwei Duplikationen des gesamten Genoms in eine Tetrade von Genloci aufgeteilt hat, und die als verwandte Region in den modernen Tunicaten zu finden ist [13]. Beim Huhn findet sich ein dem LRC homologer Bereich auf Mikrochromosom 31. Dieser Genort enthält zahlreiche Vertreter der Hühner Ig-ähnlichen Rezeptoren (CHIR), einer hochpolymorphen Genfamilie. Wie es für Mitglieder der IgSF charakteristisch ist, gibt es hemmende, aktivierende und bifunktionale CHIR [14]. Ein erster Versuch die genomische Organisation des LRC des Huhns näher zu beschreiben wurde anhand von artifiziellen Chromosomen (BAC) -Klonen eines Huhns der Linie Lohmann Leghorn (LSL) vorgenommen [15]. Die Untersuchung des LRC beim Bankivahuhn (RJF) ist ein weiterer Schritt zur Aufklärung der Bedeutung dieses genomischen Bereiches und der hier kodierten Rezeptoren. 2
8 Literatur 2 Literatur 2.1 Ig-ähnliche Rezeptoren Ig-ähnliche Rezeptoren sind Mitglieder der IgSF, welche aus vielen strukturell ähnlichen Rezeptoren besteht [16,17] und an der Regulierung des Immunsystem von Vertebraten beteiligt ist [18,19]. Diese Rezeptoren sind auf der Zelloberfläche exprimierte Proteine und haben mit anderen Mitgliedern der IgSF mehrere Eigenschaften gemein z.b. das Vorhandensein von spezifischen Ig-Domänen, Disulfidbrücken zwischen den Ketten, abwechselnde hydrophobe und hydrophile Reste und die für eine spezielle Faltung wichtigen strukturellen Besonderheiten [20,21]. Ig-ähnliche Rezeptoren haben eine oder mehrere extrazelluläre Ig-Domänen, einen transmembranen sowie einen langen oder kurzen zytoplasmatischen Anteil [4]. Die extrazelluläre Region besteht aus einer verschiedenen Anzahl von Ig-Domänen. Eine Ig-Domäne setzt sich aus zwei β-faltblättern zusammen, welche durch Disulfidbrücken stabilisiert werden. Je nach Anzahl der β-ketten, aus denen die Faltblätter bestehen, und der Länge der Sequenzen zwischen den Disulfidbrücken unterscheidet man V-, C1- und C2-Domänen. In antigenerkennenden Proteinen wie dem T-Zellrezeptor kommen die variablen V-Domänen, in Haupthistokompatibilitätskomplex (MHC) -Proteinen C1-Domänen und in Igähnlichen Rezeptoren nur C2-Domänen vor [20]. Aktivierende und inhibierende Rezeptoren im LRC unterscheiden sich in ihren transmembranen und zytoplasmatischen Anteilen. Aktivierende Rezeptoren besitzen einen kurzen zytoplasmatischen Teil und eine positiv geladene AS (Arginin oder Lysin) in der transmembranen Domäne, die an den negativ geladenen transmembranen Aspartatrest eines Adaptormoleküls binden kann, welches ITAM besitzt. Je nach Rezeptor handelt es sich dabei um Assoziierung mit DAP12 [22], der γ-kette des FcεR oder CD3ζ [23]. Ein ITAM besteht aus einer Folge von vier AS, wobei Tyrosin und Leucin den Rahmen für zwei beliebige AS bilden (YxxL). Diese Abfolge wiederholt sich getrennt durch sechs bis acht AS [24]. Die Peptide der ITAM interagieren mit den Tyrosinkinasen Syk oder ZAP-70, welche weitere Kaskaden zur Zellaktivierung auslösen [25]. Die inhibitorischen Rezeptoren haben keine positive Ladung im transmembranen Bereich sondern einen langen zytoplasmatischen Teil mit ein bis vier ITIM. Dieses Motiv ist eine Sequenzfolge von sechs AS, wobei die erste Isoleucin, Valin, Leucin 3
9 Literatur oder Serin sein kann. Es folgt Tyrosin auf eine beliebige AS und nach zwei wiederum beliebigen AS bildet Valin oder Leucin den Abschluß (I/V/L/SxYxxL/V) [26]. Durch Ligandenbindung werden die Tyrosinreste im ITIM phosphoryliert und können dann zytoplasmatische Phosphatasen mit einer Src homology 2 (SH2)-Domäne binden, welche schließlich durch Dephosphorylierung aktivierende Kaskaden unterbrechen [25]. Abb. 1: Schematische Darstellung des strukturellen Aufbaus der Ig-ähnlichen Rezeptoren mit extrazellulären Ig-Domänen (hier beispielhaft zwei) stabilisiert durch Disulfidbrücken (SS). Der inhibitorische Rezeptor (links) mit ITIM (hier beispielhaft zwei). Der aktivierende Rezeptor (rechts) mit einer positiv geladenen AS im transmembranen Bereich, durch die eine Assoziierung mit einem homodimeren ITAM-haltigen Adaptermolekül eingegangen werden kann. Abbildung entnommen aus der Dissertation von B. Viertlböck [27]. 2.2 LRC Der LRC, der beim Mensch auf Chromosom 19q13.4 [3] und bei der Maus auf Chromosom 7 zu finden ist, erstreckt sich über 1Mb und beinhaltet 30 Ig-ähnliche Rezeptorgene [4,5]. Basierend auf genetischer Organisation und Struktur können diese in vier Gruppen eingeteilt werden. Die erste Gruppe bilden die ILT, die zweite besteht aus KIR und GPVI, eine dritte Gruppe stellen LAIR dar und als viertes können NKp46 und der Rezeptor für FcαRI einander zugeordnet werden [4]. Bei ihnen allen kommen nur extrazelluläre Ig-Domänen vom C2-Typ vor [20]. 4
10 Literatur ILT ILT, auch bezeichnet als Leukozyten Ig-ähnliche Rezeptoren (LILR) oder Monozyten/Makrophagen Ig-ähnliche Rezeptoren (MIR), sind potente Modulatoren von Immunantworten wie auch die homologen gepaarten Ig-ähnlichen Rezeptoren (PIR) der Maus. Die ILT-Familie von 13 Genen liegt in zwei spiegelverkehrt duplizierten Klustern vor und ist durch ein Paar LAIR-Gene und die unabhängige Genfamilie der LENG getrennt [1,28]. Jeder der beiden Kluster umfasst 150 kb der Gensequenz zentromer zur KIR-Familie [4]. Es gibt sechs Gene für potentiell aktivierende (LILRA), fünf Gene für hemmende (LILRB) Rezeptoren und zwei Pseudogene [5]. ILT werden auf allen Zellen der myeloiden Linie sowie auf den plasmazytotischen Dentritischen Zellen [29] teilweise auf NK-Zellen, T-Zellen, Granulozyten und LILRB1 auf B-Zellen exprimiert. ILT besitzen vier extrazytoplasmatische Ig-Domänen, mit Ausnahme von ILT3, das nur zwei hat. Hemmende Rezeptoren verfügen über vier ITIM, mit denen sie die SH2- beinhaltende Proteintyrosinphosphatase (SHP) -1 [30] rekrutieren. Diese unterbricht durch Phosphorylierung des carboxyterminalen Tyrosinrestes in Src-Kinasen deren katalytische Aktivität [31]. Aktivierende ILT haben Arginin im transmembranen Bereich und interagieren mit der γ-kette des Adaptermolekül FcεRI [32]. Die einzig bekannten Liganden für einige ILT sind MHC-Klasse I-Moleküle [19]. ILT binden an die stärker konservierte α3-untereinheit. Betrachtet man die genomische Organisation und Homologie mit murinen PIR scheint sich der ILT-Genort vor dem KIR-Kluster entwickelt zu haben [4]. Im Gegensatz zu KIR haben die ILT begrenzte allelische Varianz [33] und konservierte Genanzahl zwischen Haplotypen mit dem Sonderfall des funktionellen LILRA3, der in manchen Individuen fehlt [1,28]. ILT Genen entsprechen die PIR Gene der Maus. Sie sind nah miteinander verwandt und könnten von gemeinsame Vorfahren abstammen [4] KIR und GPVI KIR-Gene liegen im LRC zwischen dem Gen für den FcαR und den ILT-Genen [34]. Diese Genfamilie beansprucht ca. 140 kb und besteht aus maximal 16 Mitgliedern und zwei Pseudogenen [4]. KIR werden auf Natürlichen Killer (NK) -Zellen sowie auf bestimmten T-Zellpopulationen exprimiert [5] und binden MHC-Klasse I-Moleküle und zwar an den hochpolymorphen α1- und α2-untereinheiten [35]. 5
11 Literatur KIR besitzen zwei oder drei extrazelluläre Ig-Domänen, wonach sie entweder als KIR2D oder KIR3D bezeichnet werden. Es gibt sowohl hemmende als auch aktivierende Varianten. Deren Namen wird aufgrund der unterschiedlichen Länge des intrazellulären Anteils bei hemmenden Rezeptoren ein L für das englische long (z.b. KIR2DL) und bei aktivierenden ein S für das englische short (z.b. KIR2DS) hinzugefügt. KIR2DL4 bildet eine Ausnahme, da er sowohl hemmende als auch aktivierende Eigenschaften hat [36]. Die aktivierenden Rezeptoren verfügen über ein geladenes Lysin im transmembranen Teil und assoziieren mit Adaptermolekül DAP12, welches ITAM besitzt. Sie lösen in letzter Konsequenz Zytotoxizität und/oder Zytokin-Freisetzung aus [22]. Die hemmenden KIR können mit den ITIM SHP-1 und SHP-2 aktivieren und die Zytotoxizität verhindern [37]. Hemmende Rezeptoren unterbrechen intrazelluläre Aktivierung, wenn MHC-Klasse I korrekt auf der Zielzelle exprimiert wird [38]. KIR können in Haplotypguppe A und B eingeteilt werden [39]. Gruppe A besteht generell aus einfacheren Rezeptoren, die meistens nur hemmende KIR darstellen während die Rezeptoren der Gruppe B im allgemeinen variabler sind und potentiell aktivierende KIR enthalten [5]. Im LRC der Maus fehlen zu den KIR homologe Gene. Das Gen für den Kollagenrezeptor GPVI liegt neben dem Gen für NKp46 am telomeren Ende des LRC, und ein homologes Gen kommt im LRC der Maus vor [4]. Dieser aktivierende Rezeptor hat zwei extrazytoplasmatische Ig-Domänen, ein positv geladenes Arginin im transmembranen Bereich und assoziiert mit der γ-kette des FcεRI. GPVI wird auf Thrombozyten von Mensch und Maus exprimiert [40] LAIR Die homologen LAIR Gene 1 und 2 kodieren angrenzend an den ILT-Kluster in unterschiedlicher transkriptionaler Orientierung [28]. LAIR-1 besitzt zwei extrazelluläre Ig-C2-Domänen und zwei ITIM im zytoplasmatischen Anteil. LAIR-2 ist zwar zu 84% homolog, besitzt aber keinen transmembranen Teil und scheint somit ein löslicher Rezeptor zu sein. LAIR-1 wird auf fast allen Immunzellen exprimiert und hemmt z.b. die zytotoxische Aktivität von T-Zellen durch Quervernetzung mit CD3 [41], oder die Kalziummobilisierung in B- Zellen durch Interaktion mit dem B-Zell Rezeptor [42]. Neben dem aktivierenden GPVI [43] ist LAIR-1 der zweite Kollagen-bindende Rezeptor im LRC. In vitro kann mit ihm die Aktivierung von primären Immunzellen gehemmt werden. 6
12 Literatur LAIR-2 bindet auch Kollagen und könnte als löslicher Rezeptor durch die Besetzung von Kollagenbindungsstellen LAIR-1 negativ regulieren [44]. Der ILT/LAIR Locus hat eine ausgedehnte inverse Duplikation durchgemacht. Da Mäusen das LAIR-2 Gen fehlt, fand diese Duplikation wohl eher später in der Evolution statt [17,45] FcαRI (CD89) und NKp46 Das FcαRI Gen befindet sich telomer des KIR-Klusters [46]. Der Rezeptor besitzt zwei extrazelluläre Ig-Domänen, ein positiv geladenes Arginin im transmembranen Teil und einen kurzen zytoplasmatischen Rest. Er ist somit ein aktivierender Rezeptor und interagiert mit dem Adaptermolekül FcεR [47]. FcαRI wird auf auf Monozyten, Makrophagen und Granulozyten exprimiert [48] und bindet IgA. Hierbei unterscheidet er nicht zwischen IgA-Subtyp 1 und 2 sowie der sekretierten bzw. monomeren oder polymeren Form [49,50]. Das NKp46-Gen liegt im LRC neben dem Gen für FcαRI und ist als Homolog im LRC der Maus vorhanden [4]. Auch dieses Protein besitzt zwei extrazelluläre Ig- Domänen einen kurzen zytoplasmatischen Rest ohne Signalmotive und transmembran einen positiv geladenen Argininrest durch den der Rezeptor mit den ITAM besitzenden Adaptermolekülen CD3ζ und FcεR assoziiert. NKp46 ist ein wichtiger Rezeptor zur Erkennung und Eliminierung von tumor- und virusinfizierten Zellen, er bindet z.b. Hämagglutinin des Influenza Virus [51,52]. Abb. 2-1: Organisation des LRC auf dem menschlichen Chromosom 19q13.4. Der Komplex ist durch vier Multigenfamilien charakterisiert, die Ig-ähnliche Rezeptoren kodieren. Erstens die Familie der ILT (auch bezeichnet als LILR)(orange), zweitens die Familie der KIR und GPVI (lila), drittens die Familie der LAIR (grün), viertens FcαR und NKp46 (rosa). Die neuen LENG-Gene (blau) sind strukturell nicht mit den Ig-ähnlichen Rezeptoren verwandt. Abbildung modifiziert nach Martin et al. [4] 7
13 Literatur 2.3 LRC des Huhns Eine zum menschlichen LRC homologe Region ist im Hühnergenom auf Mikrochromosom 31 lokalisiert. An diesem Genort befindet sich die Familie der CHIR-Gene [53,14]. CHIR gehören zur IgSF und besitzen wie alle Mitglieder dieser Genfamilie nur Ig-C2-Domänen. Man kann die CHIR in drei große Gruppen einteilen: aktivierende (CHIRA1, 2), hemmende (CHIRB1-6) und bifunktionale (CHIRAB1-3) Rezeptoren. Die hemmenden CHIRB besitzen einen langen zytoplasmatischen Teil mit zwei ITIM, während die aktivierenden CHIRA nur einen kurzen zytoplasmatischen Rest haben. Sie verfügen aber über eine positiv geladene AS im transmembranen Bereich, um mit Adaptermolekülen zu assoziieren. Die bifunktionalen Rezeptoren CHIRAB schließlich weisen sowohl hemmende als auch aktivierende Eigenschaften auf [14]. CHIRA, CHIRB und CHIRAB3 haben extrazellulär zwei Ig-Domänen, welche als Ig1- Domäne distal und Ig2-Domäne proximal der Zellmembran bezeichnet werden. Dagegen besitzen CHIRAB1 und CHIRAB2 nur eine Ig1-Domäne. Vergleicht man die Ig1- und Ig2-Domäne eines CHIR, so besteht hier nur ca % Identität der AS, während beim Vergleich von Ig1-Domänen bzw. Ig2-Domänen verschiedener CHIR eine Übereinstimmung der AS von ca % vorliegt. Sowohl CHIRA als auch CHIRAB besitzen eine positiv geladene AS im transmembranen Teil, entweder Arginin oder Lysin, in charakteristischer Position, was die Möglichkeit bietet mit einem Adaptermolekül zu assoziieren. So bindet z.b. CHIRA2 an die γ-kette des Adaptermoleküls FcεRI, welches aktivierende ITAM besitzt [14]. Alle CHIR außer CHIRAB3 weisen außerdem transmembran einen konservierten Cysteinrest nahe der extrazellulären Domäne auf, der eine Möglichkeit zur kovalenten Disulfidbrückenbindung darstellt. Hierdurch kann z.b. eine Dimerisation stattfinden, wie für CHIRB2 gezeigt werden konnte [54,14]. Dabei ist das Cystein in der transmembranen α-helix gegenüber der geladenen AS positioniert. Diese Stellung gibt Grund zur Annahme, dass eine cysteinmediierte Disulfidbrückenbindung die Assoziierung mit einem Adaptermolekül über die positiv geladene AS nicht stört. Im Gegensatz dazu fehlt CHIRA1 das konservierte Arginin oder Lysin. Dieser Rezeptor besitzt aber ein positiv geladenes Histidin 16 AS vom konservierten Cystein entfernt [55]. Diese Lokalisierung bedeutet, dass Cystein und Histidin auf der gleichen Seite der α-helix liegen und somit aufgrund der räumlichen Nähe entweder 8
14 Literatur eine cysteinmediierte Bindung oder eine Assoziierung mit einem Adaptermolekül erfolgen kann [14]. Während CHIRB1 - CHIRB4 zwei intrazytoplasmatische ITIM besitzen [54], wurde bei CHIRB5 ein amino/nh 2 -terminales ITIM und ein carboxyterminales Immunorezeptor Tyrosin-basierendes Wechselmotiv (ITSM) gefunden. Beim ITSM handelt es sich um eine Modifikation des ITIM, in der die erste AS durch ein Threonin ersetzt ist (TxYxxL/V). Im CHIRAB1 findet man transmembran ein positiv geladenes Arginin und im CHIRAB2 ein Lysin. CHIRAB1 und CHIRAB2 besitzen einen langen zytoplasmatischen Teil mit einem carboxyterminalen ITIM. Nähere Untersuchung der Position des NH 2 -terminalen ITIM bei diesen Rezeptoren ergab in der AS-Abfolge einen Wechsel zum Methionin, drei AS vom konservierten Tyrosin entfernt. Diese Anordnung bildet ein YxxM Motiv, welches in aktivierenden Signalmolekülen wie DAP10 zu finden ist [56]. CHIRAB3 besitzt ein ITIM und ein ITSM zusammen mit einem positiven transmembranen Rest [14]. Außer CHIRAB2, der nur auf Makrophagen der Linie BM-2 nachgewiesen wurde, werden CHIR auf verschiedenen Leukozyten wie reifen B-Zellen, Makrophagen, Monozyten und NK-Zellen exprimiert [14,57]. Über die Liganden der CHIR ist noch nicht viel bekannt. Erst vor kurzem wurde ein CHIRAB mit einer einzelnen extrazellulären Ig-Domäne (CHIRAB1) als hochaffiner FcY-Rezeptor identifiziert und als CHIR-AB1 bezeichnet. Dieser Rezeptor vereint Eigenschaften verschiedener Säuger Fc-Rezeptoren, wie die Expression auf B- Zellen, Monozyten und NK-Zellen und seine potentielle Bifunktionalität. Allerdings binden nicht alle CHIRAB mit einer Ig-Domäne an IgY. Die Rezeptoren die mit IgY interagieren, unterscheiden sich von denen die keine Bindung eingehen in der Position bestimmter AS der extrazellulären Ig-Domäne [57,58]. 9

15 Literatur 10 Abb. 2-2: Eigenschaften der IgY-Bindungsstelle. A Ribbon Diagram der Kristallstruktur des CHIR- AB1, erstellt mithilfe der Datei 2vsd. der Proteindatenbank. Die β-stränge a, b und e sind blau, die Stränge c, c, f, g und g grün dargestellt. Durch Mutationsanalysen identifizierte AS, die mit IgY interagieren, sind durch rote Striche verbunden. Abbildung modifiziert nach Viertlböck et al. [57]. B Vergleich der Ig1-Domäne der CHIRAB1-Rezeptoren des RJF unter Anwendung von Clustal W. Rot unterlegt sind die von Viertlböck et al. [57] identifizierten AS, die für die IgY-Bindung entscheidend sind. Grau unterlegt sind die Rezeptoren, welche potenziell in der Lage sind IgY zu binden. Aus Southern Blot Analysen mit DNA aus einzelnen Tieren von vier verschiedenen Linien und aus der Untersuchung des Genoms vom LSL [15] geht hervor, dass der LRC beim Huhn hochpolymorph ist, und es sich bei den CHIR um eine Genfamilie mit einer sehr großen Anzahl an Mitgliedern handelt [14]. L P R P S L S L H P S Q G V S L G D T V T L R C H L P R M A A W V Q L W L N G T L R F N K E K D K E Q D A A E F S F A V T N L E D A G T Y Q C R Y Q V S E P L W T S N Q S D P V E L V L T H L S L Q F D L V... Q L E M H M.. I H K..... K K.. V K.. R.. K K.... K.... M N M H K K K.. R.. K K L P F.. S... D R.. K K H M.. N T..... L E T K.. R.. K K H A.. M. E N T..... H.. S T T K.. R.. K K H A.. M. E N T..... H.. S T K.. R.. K K L P R P S L S L H P S Q G V S L G D T V T L R C H L P R M A A W V Q L W L N G T L R F N K E K D K E Q D A A E F S F A V T N L E D A G T Y Q C R Y Q V S E P L W T S N Q S D P V E L V L T H L S L Q F D L V... Q L E M H M.. I H K..... K K.. V K.. R.. K K.... K.... M N M H K K K.. R.. K K L P F.. S... D R.. K K H M.. N T..... L E T K.. R.. K K H A.. M. E N T..... H.. S T T K.. R.. K K H A.. M. E N T..... H.. S T K.. R.. K K Majority Ig1 RFJ CHIRAB1/01 Ig1 RJF CHIRAB1/03 Ig1 RJF CHIRAB1/04 Ig1 RJF CHIRAB1/06 Ig1 RJF CHIRAB1/08 Ig1 RJF CHIRAB1/11 Ig1 RJF CHIRAB1/13 Ig1 RJF CHIRAB1/14 Ig1 RJF CHIRAB1/02 Ig1 RJF CHIRAB1/05 Ig1 RJF CHIRAB1/07 Ig1 RJF CHIRAB1/09 Ig1 RJF CHIRAB1/10 Ig1 RJF CHIRAB1/12 Majority Ig1 RFJ CHIRAB1/01 Ig1 RJF CHIRAB1/03 Ig1 RJF CHIRAB1/04 Ig1 RJF CHIRAB1/06 Ig1 RJF CHIRAB1/08 Ig1 RJF CHIRAB1/11 Ig1 RJF CHIRAB1/13 Ig1 RJF CHIRAB1/14 Ig1 RJF CHIRAB1/02 Ig1 RJF CHIRAB1/05 Ig1 RJF CHIRAB1/07 Ig1 RJF CHIRAB1/09 Ig1 RJF CHIRAB1/10 Ig1 RJF CHIRAB1/12 L P R P S L S L H P S Q G V S L G D T V T L R C H L P R M A A W V Q L W L N G T L R F N K E K D K E Q D A A E F S F A V T N L E D A G T Y Q C R Y Q V S E P L W T S N Q S D P V E L V L T H L S L Q F D L V... Q L E M H M.. I H K..... K K.. V K.. R.. K K.... K.... M N M H K K K.. R.. K K L P F.. S... D R.. K K H M.. N T..... L E T K.. R.. K K H A.. M. E N T..... H.. S T T K.. R.. K K H A.. M. E N T..... H.. S T K.. R.. K K L P R P S L S L H P S Q G V S L G D T V T L R C H L P R M A A W V Q L W L N G T L R F N K E K D K E Q D A A E F S F A V T N L E D A G T Y Q C R Y Q V S E P L W T S N Q S D P V E L V L T H L S L Q F D L V... Q L E M H M.. I H K..... K K.. V K.. R.. K K.... K.... M N M H K K K.. R.. K K L P F.. S... D R.. K K H M.. N T..... L E T K.. R.. K K H A.. M. E N T..... H.. S T T K.. R.. K K H A.. M. E N T..... H.. S T K.. R.. K K Majority Ig1 RFJ CHIRAB1/01 Ig1 RJF CHIRAB1/03 Ig1 RJF CHIRAB1/04 Ig1 RJF CHIRAB1/06 Ig1 RJF CHIRAB1/08 Ig1 RJF CHIRAB1/11 Ig1 RJF CHIRAB1/13 Ig1 RJF CHIRAB1/14 Ig1 RJF CHIRAB1/02 Ig1 RJF CHIRAB1/05 Ig1 RJF CHIRAB1/07 Ig1 RJF CHIRAB1/09 Ig1 RJF CHIRAB1/10 Ig1 RJF CHIRAB1/12 Majority Ig1 RFJ CHIRAB1/01 Ig1 RJF CHIRAB1/03 Ig1 RJF CHIRAB1/04 Ig1 RJF CHIRAB1/06 Ig1 RJF CHIRAB1/08 Ig1 RJF CHIRAB1/11 Ig1 RJF CHIRAB1/13 Ig1 RJF CHIRAB1/14 Ig1 RJF CHIRAB1/02 Ig1 RJF CHIRAB1/05 Ig1 RJF CHIRAB1/07 Ig1 RJF CHIRAB1/09 Ig1 RJF CHIRAB1/10 Ig1 RJF CHIRAB1/12 L P R P S L S L H P S Q G V S L G D T V T L R C H L P R M A A W V Q L W L N G T L R F N K E K D K E Q D A A E F S F A V T N L E D A G T Y Q C R Y Q V S E P L W T S N Q S D P V E L V L T H L S L Q F D L V... Q L E M H M.. I H K..... K K.. V K.. R.. K K.... K.... M N M H K K K.. R.. K K L P F.. S... D R.. K K H M.. N T..... L E T K.. R.. K K H A.. M. E N T..... H.. S T T K.. R.. K K H A.. M. E N T..... H.. S T K.. R.. K K L P R P S L S L H P S Q G V S L G D T V T L R C H L P R M A A W V Q L W L N G T L R F N K E K D K E Q D A A E F S F A V T N L E D A G T Y Q C R Y Q V S E P L W T S N Q S D P V E L V L T H L S L Q F D L V... Q L E M H M.. I H K..... K K.. V K.. R.. K K.... K.... M N M H K K K.. R.. K K L P F.. S... D R.. K K H M.. N T..... L E T K.. R.. K K H A.. M. E N T..... H.. S T T K.. R.. K K H A.. M. E N T..... H.. S T K.. R.. K K Majority Ig1 RFJ CHIRAB1/01 Ig1 RJF CHIRAB1/03 Ig1 RJF CHIRAB1/04 Ig1 RJF CHIRAB1/06 Ig1 RJF CHIRAB1/08 Ig1 RJF CHIRAB1/11 Ig1 RJF CHIRAB1/13 Ig1 RJF CHIRAB1/14 Ig1 RJF CHIRAB1/02 Ig1 RJF CHIRAB1/05 Ig1 RJF CHIRAB1/07 Ig1 RJF CHIRAB1/09 Ig1 RJF CHIRAB1/10 Ig1 RJF CHIRAB1/12 Majority Ig1 RFJ CHIRAB1/01 Ig1 RJF CHIRAB1/03 Ig1 RJF CHIRAB1/04 Ig1 RJF CHIRAB1/06 Ig1 RJF CHIRAB1/08 Ig1 RJF CHIRAB1/11 Ig1 RJF CHIRAB1/13 Ig1 RJF CHIRAB1/14 Ig1 RJF CHIRAB1/02 Ig1 RJF CHIRAB1/05 Ig1 RJF CHIRAB1/07 Ig1 RJF CHIRAB1/09 Ig1 RJF CHIRAB1/10 Ig1 RJF CHIRAB1/12 L P R P S L S L H P S Q G V S L G D T V T L R C H L P R M A A W V Q L W L N G T L R F N K E K D K E Q D A A E F S F A V T N L E D A G T Y Q C R Y Q V S E P L W T S N Q S D P V E L V L T H L S L Q F D L V... Q L E M H M.. I H K..... K K.. V K.. R.. K K.... K.... M N M H K K K.. R.. K K L P F.. S... D R.. K K H M.. N T..... L E T K.. R.. K K H A.. M. E N T..... H.. S T T K.. R.. K K H A.. M. E N T..... H.. S T K.. R.. K K L P R P S L S L H P S Q G V S L G D T V T L R C H L P R M A A W V Q L W L N G T L R F N K E K D K E Q D A A E F S F A V T N L E D A G T Y Q C R Y Q V S E P L W T S N Q S D P V E L V L T H L S L Q F D L V... Q L E M H M.. I H K..... K K.. V K.. R.. K K.... K.... M N M H K K K.. R.. K K L P F.. S... D R.. K K H M.. N T..... L E T K.. R.. K K H A.. M. E N T..... H.. S T T K.. R.. K K H A.. M. E N T..... H.. S T K.. R.. K K Majority Ig1 RFJ CHIRAB1/01 Ig1 RJF CHIRAB1/03 Ig1 RJF CHIRAB1/04 Ig1 RJF CHIRAB1/06 Ig1 RJF CHIRAB1/08 Ig1 RJF CHIRAB1/11 Ig1 RJF CHIRAB1/13 Ig1 RJF CHIRAB1/14 Ig1 RJF CHIRAB1/02 Ig1 RJF CHIRAB1/05 Ig1 RJF CHIRAB1/07 Ig1 RJF CHIRAB1/09 Ig1 RJF CHIRAB1/10 Ig1 RJF CHIRAB1/12 Majority Ig1 RFJ CHIRAB1/01 Ig1 RJF CHIRAB1/03 Ig1 RJF CHIRAB1/04 Ig1 RJF CHIRAB1/06 Ig1 RJF CHIRAB1/08 Ig1 RJF CHIRAB1/11 Ig1 RJF CHIRAB1/13 Ig1 RJF CHIRAB1/14 Ig1 RJF CHIRAB1/02 Ig1 RJF CHIRAB1/05 Ig1 RJF CHIRAB1/07 Ig1 RJF CHIRAB1/09 Ig1 RJF CHIRAB1/10 Ig1 RJF CHIRAB1/12 L P R P S L S L H P S Q G V S L G D T V T L R C H L P R M A A W V Q L W L N G T L R F N K E K D K E Q D A A E F S F A V T N L E D A G T Y Q C R Y Q V S E P L W T S N Q S D P V E L V L T H L S L Q F D L V... Q L E M H M.. I H K..... K K.. V K.. R.. K K.... K.... M N M H K K K.. R.. K K L P F.. S... D R.. K K H M.. N T..... L E T K.. R.. K K H A.. M. E N T..... H.. S T T K.. R.. K K H A.. M. E N T..... H.. S T K.. R.. K K L P R P S L S L H P S Q G V S L G D T V T L R C H L P R M A A W V Q L W L N G T L R F N K E K D K E Q D A A E F S F A V T N L E D A G T Y Q C R Y Q V S E P L W T S N Q S D P V E L V L T H L S L Q F D L V... Q L E M H M.. I H K..... K K.. V K.. R.. K K.... K.... M N M H K K K.. R.. K K L P F.. S... D R.. K K H M.. N T..... L E T K.. R.. K K H A.. M. E N T..... H.. S T T K.. R.. K K H A.. M. E N T..... H.. S T K.. R.. K K Majority Ig1 RFJ CHIRAB1/01 Ig1 RJF CHIRAB1/03 Ig1 RJF CHIRAB1/04 Ig1 RJF CHIRAB1/06 Ig1 RJF CHIRAB1/08 Ig1 RJF CHIRAB1/11 Ig1 RJF CHIRAB1/13 Ig1 RJF CHIRAB1/14 Ig1 RJF CHIRAB1/02 Ig1 RJF CHIRAB1/05 Ig1 RJF CHIRAB1/07 Ig1 RJF CHIRAB1/09 Ig1 RJF CHIRAB1/10 Ig1 RJF CHIRAB1/12 Majority Ig1 RFJ CHIRAB1/01 Ig1 RJF CHIRAB1/03 Ig1 RJF CHIRAB1/04 Ig1 RJF CHIRAB1/06 Ig1 RJF CHIRAB1/08 Ig1 RJF CHIRAB1/11 Ig1 RJF CHIRAB1/13 Ig1 RJF CHIRAB1/14 Ig1 RJF CHIRAB1/02 Ig1 RJF CHIRAB1/05 Ig1 RJF CHIRAB1/07 Ig1 RJF CHIRAB1/09 Ig1 RJF CHIRAB1/10 Ig1 RJF CHIRAB1/12 L P R P S L S L H P S Q G V S L G D T V T L R C H L P R M A A W V Q L W L N G T L R F N K E K D K E Q D A A E F S F A V T N L E D A G T Y Q C R Y Q V S E P L W T S N Q S D P V E L V L T H L S L Q F D L V... Q L E M H M.. I H K..... K K.. V K.. R.. K K.... K.... M N M H K K K.. R.. K K L P F.. S... D R.. K K H M.. N T..... L E T K.. R.. K K H A.. M. E N T..... H.. S T T K.. R.. K K H A.. M. E N T..... H.. S T K.. R.. K K L P R P S L S L H P S Q G V S L G D T V T L R C H L P R M A A W V Q L W L N G T L R F N K E K D K E Q D A A E F S F A V T N L E D A G T Y Q C R Y Q V S E P L W T S N Q S D P V E L V L T H L S L Q F D L V... Q L E M H M.. I H K..... K K.. V K.. R.. K K.... K.... M N M H K K K.. R.. K K L P F.. S... D R.. K K H M.. N T..... L E T K.. R.. K K H A.. M. E N T..... H.. S T T K.. R.. K K H A.. M. E N T..... H.. S T K.. R.. K K Majority Ig1 RFJ CHIRAB1/01 Ig1 RJF CHIRAB1/03 Ig1 RJF CHIRAB1/04 Ig1 RJF CHIRAB1/06 Ig1 RJF CHIRAB1/08 Ig1 RJF CHIRAB1/11 Ig1 RJF CHIRAB1/13 Ig1 RJF CHIRAB1/14 Ig1 RJF CHIRAB1/02 Ig1 RJF CHIRAB1/05 Ig1 RJF CHIRAB1/07 Ig1 RJF CHIRAB1/09 Ig1 RJF CHIRAB1/10 Ig1 RJF CHIRAB1/12 Majority Ig1 RFJ CHIRAB1/01 Ig1 RJF CHIRAB1/03 Ig1 RJF CHIRAB1/04 Ig1 RJF CHIRAB1/06 Ig1 RJF CHIRAB1/08 Ig1 RJF CHIRAB1/11 Ig1 RJF CHIRAB1/13 Ig1 RJF CHIRAB1/14 Ig1 RJF CHIRAB1/02 Ig1 RJF CHIRAB1/05 Ig1 RJF CHIRAB1/07 Ig1 RJF CHIRAB1/09 Ig1 RJF CHIRAB1/10 Ig1 RJF CHIRAB1/12 L P R P S L S L H P S Q G V S L G D T V T L R C H L P R M A A W V Q L W L N G T L R F N K E K D K E Q D A A E F S F A V T N L E D A G T Y Q C R Y Q V S E P L W T S N Q S D P V E L V L T H L S L Q F D L V... Q L E M H M.. I H K..... K K.. V K.. R.. K K.... K.... M N M H K K K.. R.. K K L P F.. S... D R.. K K H M.. N T..... L E T K.. R.. K K H A.. M. E N T..... H.. S T T K.. R.. K K H A.. M. E N T..... H.. S T K.. R.. K K L P R P S L S L H P S Q G V S L G D T V T L R C H L P R M A A W V Q L W L N G T L R F N K E K D K E Q D A A E F S F A V T N L E D A G T Y Q C R Y Q V S E P L W T S N Q S D P V E L V L T H L S L Q F D L V... Q L E M H M.. I H K..... K K.. V K.. R.. K K.... K.... M N M H K K K.. R.. K K L P F.. S... D R.. K K H M.. N T..... L E T K.. R.. K K H A.. M. E N T..... H.. S T T K.. R.. K K H A.. M. E N T..... H.. S T K.. R.. K K Majority Ig1 RFJ CHIRAB1/01 Ig1 RJF CHIRAB1/03 Ig1 RJF CHIRAB1/04 Ig1 RJF CHIRAB1/06 Ig1 RJF CHIRAB1/08 Ig1 RJF CHIRAB1/11 Ig1 RJF CHIRAB1/13 Ig1 RJF CHIRAB1/14 Ig1 RJF CHIRAB1/02 Ig1 RJF CHIRAB1/05 Ig1 RJF CHIRAB1/07 Ig1 RJF CHIRAB1/09 Ig1 RJF CHIRAB1/10 Ig1 RJF CHIRAB1/12 Majority Ig1 RFJ CHIRAB1/01 Ig1 RJF CHIRAB1/03 Ig1 RJF CHIRAB1/04 Ig1 RJF CHIRAB1/06 Ig1 RJF CHIRAB1/08 Ig1 RJF CHIRAB1/11 Ig1 RJF CHIRAB1/13 Ig1 RJF CHIRAB1/14 Ig1 RJF CHIRAB1/02 Ig1 RJF CHIRAB1/05 Ig1 RJF CHIRAB1/07 Ig1 RJF CHIRAB1/09 Ig1 RJF CHIRAB1/10 Ig1 RJF CHIRAB1/12 L P R P S L S L H P S Q G V S L G D T V T L R C H L P R M A A W V Q L W L N G T L R F N K E K D K E Q D A A E F S F A V T N L E D A G T Y Q C R Y Q V S E P L W T S N Q S D P V E L V L T H L S L Q F D L V... Q L E M H M.. I H K..... K K.. V K.. R.. K K.... K.... M N M H K K K.. R.. K K L P F.. S... D R.. K K H M.. N T..... L E T K.. R.. K K H A.. M. E N T..... H.. S T T K.. R.. K K H A.. M. E N T..... H.. S T K.. R.. K K L P R P S L S L H P S Q G V S L G D T V T L R C H L P R M A A W V Q L W L N G T L R F N K E K D K E Q D A A E F S F A V T N L E D A G T Y Q C R Y Q V S E P L W T S N Q S D P V E L V L T H L S L Q F D L V... Q L E M H M.. I H K..... K K.. V K.. R.. K K.... K.... M N M H K K K.. R.. K K L P F.. S... D R.. K K H M.. N T..... L E T K.. R.. K K H A.. M. E N T..... H.. S T T K.. R.. K K H A.. M. E N T..... H.. S T K.. R.. K K Majority Ig1 RFJ CHIRAB1/01 Ig1 RJF CHIRAB1/03 Ig1 RJF CHIRAB1/04 Ig1 RJF CHIRAB1/06 Ig1 RJF CHIRAB1/08 Ig1 RJF CHIRAB1/11 Ig1 RJF CHIRAB1/13 Ig1 RJF CHIRAB1/14 Ig1 RJF CHIRAB1/02 Ig1 RJF CHIRAB1/05 Ig1 RJF CHIRAB1/07 Ig1 RJF CHIRAB1/09 Ig1 RJF CHIRAB1/10 Ig1 RJF CHIRAB1/12 Majority Ig1 RFJ CHIRAB1/01 Ig1 RJF CHIRAB1/03 Ig1 RJF CHIRAB1/04 Ig1 RJF CHIRAB1/06 Ig1 RJF CHIRAB1/08 Ig1 RJF CHIRAB1/11 Ig1 RJF CHIRAB1/13 Ig1 RJF CHIRAB1/14 Ig1 RJF CHIRAB1/02 Ig1 RJF CHIRAB1/05 Ig1 RJF CHIRAB1/07 Ig1 RJF CHIRAB1/09 Ig1 RJF CHIRAB1/10 Ig1 RJF CHIRAB1/12 A B





16 Publikation 3 Publikation 11





17 Publikation 12
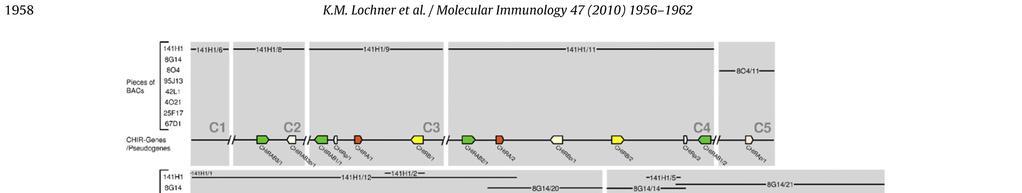
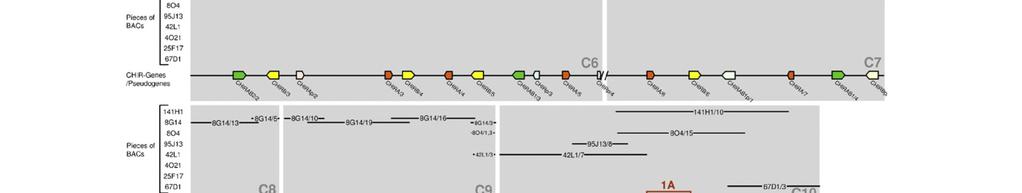
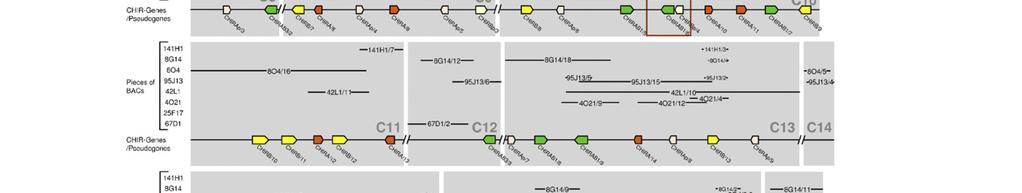
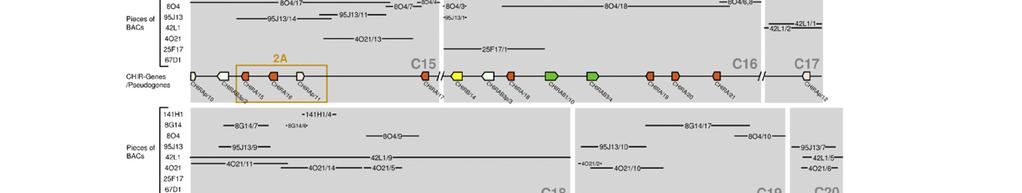


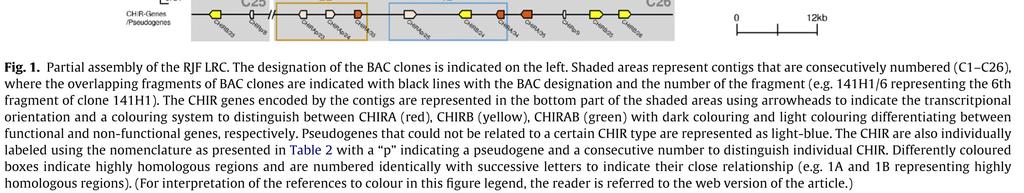
18 Publikation 13

19 Publikation 14

20 Publikation 15







21 Publikation 16



22 Publikation 17
23 Diskussion 4 Diskussion Das Hühnergenom-Sequenzierungsprojekt und die Erstellung der Wageningen Hühner-BAC-Bibliothek sind wichtige Schritte, um die genomische Organisation der einzelnen Chromosomen und die Genstrukturen des Huhns zu erfassen. Bei der Untersuchung der Chromosomen des Huhns wurde festgestellt, dass die 33 Mikrochromosomen ca. ein Drittel der genomischen DNA darstellen. Sie sind aber so genreich, dass sie vermutlich doppelt so viele Gene beinhalten wie die sechs Makrochromosomen [1]. Allerdings konnten noch nicht alle Bereiche annotiert werden, so wie beispielsweise das Mikrochromosom 31. Mithilfe von Fluoreszenz-insitu-Hybridisierung konnte gezeigt werden, dass sowohl der LRC Marker ROSO264 [53] als auch eine CHIR-DNA Probe auf Mikrochromosom 31 hybridisieren. Das verdeutlichte, dass dieses Mikrochromosom den LRC des Huhns beinhaltet und aufgrund des starken Signals hier viele CHIR zu finden sind [14]. Erstmals wurde 2006 versucht, anhand der BAC-Bibliothek aus dem Genom des LSL-Huhns die Sequenzstücke und Gene des Hühner-LRC zu ordnen. Dabei fanden Laun et al. in sieben BAC-Klonen, welche insgesamt 550kb umfassen, über 100 CHIR [15]. Um die bessere Erfassung dieses Genortes und seiner Bedeutung voranzutreiben, wurde in der vorliegenden Arbeit versucht, die entsprechenden BAC- Sequenzen des Genoms eines einzelnen ingezüchteten RJF zu ordnen, um die Sequenzen und die Gene des LRC dieses Tieres näher zu untersuchen. Das Problem ist, dass die acht zur Verfügung stehenden BAC-Klone aus bis zu 21 ungeordneten Stücken bestehen. Trotz Verlängerung dieser Teilsequenzen mit Hilfe der Trace Archive Datenbank des National Center for Biotechnology Information und zusätzlichen Sequenzstücken des Washington University Genome Center, konnte keine durchgehende Sequenz erstellt werden. Bisherige Untersuchungen ergaben, dass die Mitglieder der CHIR-Genfamilie sehr zahlreich und verschiedenartig, aber auch in vielen Sequenzabschnitten identisch sind [14,15]. Diese Tatsache trägt entscheidend zu dem Problem bei die Sequenzstücke einander zuzuordnen und die Annotation der Gene in diesem Bereich vorzunehmen. Ebenso wie im LSL-Huhn [15] konnten im Genom des RJF in 875kb über 100 CHIR identifiziert werden. Vergleicht man diese mit den homologen Regionen anderer Wirbeltiere, dann fällt beim Huhn die große Anzahl an verschiedenartigen Mitgliedern dieser IgSF auf. So beinhaltet z.b. der LRC des Menschen, der 1Mb umfasst, nur 30 18
24 Diskussion Gene der IgSF [4] und in marinen Fleischfressern (Robben und Seelöwen) findet man sogar nur einen einzigen, schwach polymorphen KIR [59]. Als Liganden für ILT und KIR im menschlichen LRC wurden humane- Leukozytenantigen (HLA) Klasse I-Moleküle identifiziert. Die Familien interagieren allerdings mit unterschiedlichen Epitopen des HLA-Klasse I-Molekül. So binden die Mitglieder der KIR an die hochpolymorphen α1- und α2-domänen, wobei die Entstehung verschiedenartiger, polymorpher KIR wahrscheinlich von Vorteil ist um diese Bindung zu optimieren. Dagegen gehen ILT eine Bindung mit der höher konservierten α3-domäne des HLA-Proteins ein, was ein Grund dafür sein könnte, dass bei diesen Rezeptoren weniger Polymorphismus festzustellen ist [4]. Warum die Vielfalt der CHIR so groß ist, ist noch nicht bekannt und hängt auch mit dem bis jetzt spärlichen Wissen über ihre Liganden zusammen. Sogar innerhalb einer Rezeptorsubgruppe wie den CHIRAB1 bindet nur ein Teil an IgY, die sogenannten CHIR-AB1-ähnlichen Rezeptoren. Andere CHIRAB1 weisen durch die Varianz nur weniger AS keine IgY-Bindung auf und haben evtl. andere Liganden [60]. Trotz der großen Ähnlichkeit der einzelnen CHIRAB1-Proteinsequenzen des RJF, die phylogenetisch sehr nah beieinander liegen, sind die Unterschiede von einzelnen AS so entscheidend, dass von 14 identifizierten CHIRAB1 nur acht Rezeptoren potenziell IgY binden können (siehe Abb. 2-2). Weiter fällt auf, dass es sich bei allen erfassten Genen in diesem Bereich ausschließlich um CHIR handelt. Eine Erklärung hierfür könnte die enorme genomische Instabilität an diesem Genort sein. Diese stellt für die CHIR scheinbar einen Vorteil dar, wird aber für die meisten anderer Gene eher von großem Nachteil sein. Aufgrund einer solchen Instabilität können unabhängige Gene, die sich auf dem gleichen chromosomalen Abschnitt befinden, mit beeinflusst werden. So wird z.b. vermutet, dass der Selektionsdruck auf den FcαR der Maus auch eine Deletion des benachbarten KIR Locus verursacht hat [61]. In 875kb, die sich in 26 Contigs aufteilen, wurden insgesamt 130 CHIR Gene gefunden und 46 von ihnen meist aufgrund mehrerer Eigenschaften als Pseudogene identifiziert, wie fehlende Exons, Leserahmenverschiebung und zu frühe Stoppcodons. Allerdings liegen einiger dieser Pseudogene nah an Contigenden und könnten nach Ergänzung durch neue Sequenzstücke durchaus funktionsfähige Gene repräsentieren. Von den 84 funktionsfähigen Genen kodieren 35 für aktivierende Rezeptoren. Frühere Untersuchungen geben Hinweise darauf, dass der 19
Immunbiologie. Teil 3
 Teil 3 Haupthistokompatibilitätskomplex (1): - es gibt einen grundlegenden Unterschied, wie B-Lymphozyten und T-Lymphozyten ihr relevantes Antigen erkennen - B-Lymphozyten binden direkt an das komplette
Teil 3 Haupthistokompatibilitätskomplex (1): - es gibt einen grundlegenden Unterschied, wie B-Lymphozyten und T-Lymphozyten ihr relevantes Antigen erkennen - B-Lymphozyten binden direkt an das komplette
Signaltransduktion durch Zell-Zell und Zell-Matrix Kontakte
 Signaltransduktion durch Zell-Zell und Zell-Matrix Kontakte - Integrine als zentrale Adhäsionsrezeptoren - - Focal Adhesion Kinase (FAK) als zentrales Signalmolekül - Regulation von Zellfunktionen durch
Signaltransduktion durch Zell-Zell und Zell-Matrix Kontakte - Integrine als zentrale Adhäsionsrezeptoren - - Focal Adhesion Kinase (FAK) als zentrales Signalmolekül - Regulation von Zellfunktionen durch
Medizinische Immunologie. Vorlesung 6 Effektormechanismen
 Medizinische Immunologie Vorlesung 6 Effektormechanismen Effektormechanismen Spezifische Abwehrmechanismen Effektormechanismen der zellulären Immunantwort - allgemeine Prinzipien - CTL (zytotoxische T-Lymphozyten)
Medizinische Immunologie Vorlesung 6 Effektormechanismen Effektormechanismen Spezifische Abwehrmechanismen Effektormechanismen der zellulären Immunantwort - allgemeine Prinzipien - CTL (zytotoxische T-Lymphozyten)
Die TILR-Familie des Truthuhns
 Aus dem Veterinärwissenschaftlichen Department der Tierärztlichen Fakultät der Ludwig-Maximilians-Universität München Arbeit angefertigt unter der Leitung von Univ.-Prof. Dr. T. Göbel Die TILR-Familie
Aus dem Veterinärwissenschaftlichen Department der Tierärztlichen Fakultät der Ludwig-Maximilians-Universität München Arbeit angefertigt unter der Leitung von Univ.-Prof. Dr. T. Göbel Die TILR-Familie
Unterrichtsmaterialien in digitaler und in gedruckter Form. Auszug aus: Einführung in die Immunbiologie. Das komplette Material finden Sie hier:
 Unterrichtsmaterialien in digitaler und in gedruckter Form Auszug aus: Einführung in die Immunbiologie Das komplette Material finden Sie hier: School-Scout.de S 2 M 2 Das Immunsystem eine Übersicht Das
Unterrichtsmaterialien in digitaler und in gedruckter Form Auszug aus: Einführung in die Immunbiologie Das komplette Material finden Sie hier: School-Scout.de S 2 M 2 Das Immunsystem eine Übersicht Das
Immunologie. Entwicklung der T- und B- Lymphozyten. Vorlesung 4: Dr. Katja Brocke-Heidrich. Die Entwicklung der T-Lymphozyten
 Immunologie Vorlesung 4: Entwicklung der T- und B- Lymphozyten T-Zellen entwickeln sich im Thymus B-Zellen entwickeln sich im Knochenmark (engl. bone marrow, aber eigentlich nach Bursa fabricius) Dr. Katja
Immunologie Vorlesung 4: Entwicklung der T- und B- Lymphozyten T-Zellen entwickeln sich im Thymus B-Zellen entwickeln sich im Knochenmark (engl. bone marrow, aber eigentlich nach Bursa fabricius) Dr. Katja
Aufgabe 5 (Supersekundärstruktur)
") Aufgabe 5 (Supersekundärstruktur) Fragestellung Bei der Untereinheit des Arthropodenhämocyanins aus Limulus polyphemus werden folgende Fragestellungen untersucht: - Welche Supersekundärstrukturen gibt
Aufgabe 5 (Supersekundärstruktur) Fragestellung Bei der Untereinheit des Arthropodenhämocyanins aus Limulus polyphemus werden folgende Fragestellungen untersucht: - Welche Supersekundärstrukturen gibt
Duale Reihe Medizinische Mikrobiologie
 Duale Reihe Medizinische Mikrobiologie von Herbert Hof, Rüdiger Dörries erweitert, überarbeitet Duale Reihe Medizinische Mikrobiologie Hof / Dörries schnell und portofrei erhältlich bei beck-shop.de DIE
Duale Reihe Medizinische Mikrobiologie von Herbert Hof, Rüdiger Dörries erweitert, überarbeitet Duale Reihe Medizinische Mikrobiologie Hof / Dörries schnell und portofrei erhältlich bei beck-shop.de DIE
T-Lymphozyten. T-Lymphozyten erkennen spezifisch nur zell- ständige Antigene (Proteine!) und greifen sie direkt an. verantwortlich.
 und greifen sie direkt an. verantwortlich.") T-Lymphozyten T-Lymphozyten erkennen spezifisch nur zell- ständige Antigene (Proteine!) und greifen sie direkt an. Sie sind für die zellvermittelte Immunität verantwortlich. Antigenerkennung B Zellen erkennen
T-Lymphozyten T-Lymphozyten erkennen spezifisch nur zell- ständige Antigene (Proteine!) und greifen sie direkt an. Sie sind für die zellvermittelte Immunität verantwortlich. Antigenerkennung B Zellen erkennen
Aufgabe 2: (Aminosäuren)
") Aufgabe 2: (Aminosäuren) Aufgabenstellung Die 20 Aminosäuren (voller Name, 1- und 3-Buchstaben-Code) sollen identifiziert und mit RasMol grafisch dargestellt werden. Dann sollen die AS sinnvoll nach ihren
Aufgabe 2: (Aminosäuren) Aufgabenstellung Die 20 Aminosäuren (voller Name, 1- und 3-Buchstaben-Code) sollen identifiziert und mit RasMol grafisch dargestellt werden. Dann sollen die AS sinnvoll nach ihren
Proteinogene Aminosäuren. Unpolare, aliphatische Seitenketten Monoaminomonocarbonsäuren
 Proteinogene Aminosäuren Unpolare, aliphatische Seitenketten Monoaminomonocarbonsäuren Proteinogene Aminosäuren Unpolare, heterozyklische Seitenkette Monoaminomonocarbonsäuren Proteinogene Aminosäuren
Proteinogene Aminosäuren Unpolare, aliphatische Seitenketten Monoaminomonocarbonsäuren Proteinogene Aminosäuren Unpolare, heterozyklische Seitenkette Monoaminomonocarbonsäuren Proteinogene Aminosäuren
Grundlagen der Immunologie
 Grundlagen der Immunologie 9. Vorlesung Die zentrale (thymische) T-Zell-Entwicklung Alle Blutzellen stammen von der multipotenten hämatopoetischen Stammzelle des Knochenmarks Figure 1-3 Zentrale Lymphozytenentwicklung
Grundlagen der Immunologie 9. Vorlesung Die zentrale (thymische) T-Zell-Entwicklung Alle Blutzellen stammen von der multipotenten hämatopoetischen Stammzelle des Knochenmarks Figure 1-3 Zentrale Lymphozytenentwicklung
UAufgabe 12: (evolutiv konservierte Aminosäuren)
") UAufgabe 12: (evolutiv konservierte Aminosäuren) Aufgabenstellung Wählen Sie zur Darstellung evolutiv konservierter Aminosäure-Positionen in "1lla" eine ihnen sinnvoll erscheinende Anfärbung. Exportieren
UAufgabe 12: (evolutiv konservierte Aminosäuren) Aufgabenstellung Wählen Sie zur Darstellung evolutiv konservierter Aminosäure-Positionen in "1lla" eine ihnen sinnvoll erscheinende Anfärbung. Exportieren
Bei Einbeziehung von neun Allelen in den Vergleich ergibt sich eine Mutation in 38 Generationen (350:9); das entspricht ca. 770 Jahren.
; das entspricht ca. 770 Jahren.") 336 DNA Genealogie Das Muster genetischer Variationen im Erbgut einer Person - zu gleichen Teilen von Mutter und Vater ererbt - definiert seine genetische Identität unveränderlich. Neben seiner Identität
336 DNA Genealogie Das Muster genetischer Variationen im Erbgut einer Person - zu gleichen Teilen von Mutter und Vater ererbt - definiert seine genetische Identität unveränderlich. Neben seiner Identität
Inhalte unseres Vortrages
 Inhalte unseres Vortrages Vorstellung der beiden paper: Germ line transmission of a disrupted ß2 mirkroglobulin gene produced by homologous recombination in embryonic stem cells ß2 Mikroglobulin deficient
Inhalte unseres Vortrages Vorstellung der beiden paper: Germ line transmission of a disrupted ß2 mirkroglobulin gene produced by homologous recombination in embryonic stem cells ß2 Mikroglobulin deficient
Hemmung der Enzym-Aktivität
 Hemmung der Enzym-Aktivität Substrat Kompetitiver Inhibitor Enzym Enzym Substrat Nichtkompetitiver Inhibitor Irreversibler Inhibitor Enzym Enzym Enzym - Kinetik Michaelis Menten Gleichung Lineweaver -
Hemmung der Enzym-Aktivität Substrat Kompetitiver Inhibitor Enzym Enzym Substrat Nichtkompetitiver Inhibitor Irreversibler Inhibitor Enzym Enzym Enzym - Kinetik Michaelis Menten Gleichung Lineweaver -
Praktikum Biochemie B.Sc. Water Science WS Enzymregulation. Marinja Niggemann, Denise Schäfer
 Praktikum Biochemie B.Sc. Water Science WS 2011 Enzymregulation Marinja Niggemann, Denise Schäfer Regulatorische Strategien 1. Allosterische Wechselwirkung 2. Proteolytische Aktivierung 3. Kovalente Modifikation
Praktikum Biochemie B.Sc. Water Science WS 2011 Enzymregulation Marinja Niggemann, Denise Schäfer Regulatorische Strategien 1. Allosterische Wechselwirkung 2. Proteolytische Aktivierung 3. Kovalente Modifikation
B-Zellentwicklung. Grundlagen der Immunologie 5. Semester - Dienstags Uhr Ruhr-Universität Bochum, HMA 20 HEV.
 Grundlagen der Immunologie 5. Semester - Dienstags 11.15 Uhr Ruhr-Universität Bochum, HMA 20 B-Zellentwicklung Albrecht Bufe www.ruhr-uni-bochum.de/homeexpneu T und B Zellen zirkulieren unablässig durch
Grundlagen der Immunologie 5. Semester - Dienstags 11.15 Uhr Ruhr-Universität Bochum, HMA 20 B-Zellentwicklung Albrecht Bufe www.ruhr-uni-bochum.de/homeexpneu T und B Zellen zirkulieren unablässig durch
AUFGABENSAMMLUNG Lösungen. Variabilität von Antikörpern 1
 Variabilität von Antikörpern 1 Rezeptoren bzw. Antikörper eines noch undifferenzierten B-Lymphocyten: a) Schreiben Sie die Anzahl der variablen Exons je Chromosom auf. b) Berechnen Sie die mögliche Anzahl
Variabilität von Antikörpern 1 Rezeptoren bzw. Antikörper eines noch undifferenzierten B-Lymphocyten: a) Schreiben Sie die Anzahl der variablen Exons je Chromosom auf. b) Berechnen Sie die mögliche Anzahl
Transplantation. Univ.-Prof. Dr. Albert Duschl
 Transplantation Univ.-Prof. Dr. Albert Duschl Grafting Transplantationen sind sehr artifiziell, sie sind aber medizinisch wichtig und wir können daraus etwas über das Immunsystem lernen. Zwei Fragen drängen
Transplantation Univ.-Prof. Dr. Albert Duschl Grafting Transplantationen sind sehr artifiziell, sie sind aber medizinisch wichtig und wir können daraus etwas über das Immunsystem lernen. Zwei Fragen drängen
Selbstmoleküle HLA oder MHC
 Grundlagen der Immunologie 5. Semester - Dienstags 11.15 Uhr Ruhr-Universität Bochum, HMA 20 MHC, Immunologische Synapse, T-Zell-Programmierung und Transplantation Albrecht Bufe www.ruhr-uni-bochum.de/homeexpneu
Grundlagen der Immunologie 5. Semester - Dienstags 11.15 Uhr Ruhr-Universität Bochum, HMA 20 MHC, Immunologische Synapse, T-Zell-Programmierung und Transplantation Albrecht Bufe www.ruhr-uni-bochum.de/homeexpneu
Erratum zur Diplomarbeit von Andreas Bender. Korrekturverzeichnis: Kapitel 3.2.2, Seiten 39-46
 Erratum zur Diplomarbeit von Andreas Bender Korrekturverzeichnis: Kapitel 3.2.2, Seiten 39-46 3.2.2. Einzelne Aminosäuren in N-terminalen Abschnitten In den Abbildungen 16-22 sind die relativen Aminosäurehäufigkeiten
Erratum zur Diplomarbeit von Andreas Bender Korrekturverzeichnis: Kapitel 3.2.2, Seiten 39-46 3.2.2. Einzelne Aminosäuren in N-terminalen Abschnitten In den Abbildungen 16-22 sind die relativen Aminosäurehäufigkeiten
Zytokine und Zytokinrezeptoren
 Zytokine: Mediatoren der interzellulären Kommunikation Zytokine und Zytokinrezeptoren - kleine, lösliche GP - beeinflussen Verhalten anderer Zellen - regulieren Wachstum und Differenzierung - Zytokinrezeptoren
Zytokine: Mediatoren der interzellulären Kommunikation Zytokine und Zytokinrezeptoren - kleine, lösliche GP - beeinflussen Verhalten anderer Zellen - regulieren Wachstum und Differenzierung - Zytokinrezeptoren
Die Rolle von KIR- und HLA-Klasse I-Polymorphismen für die Funktion von Natürlichen Killerzellen in gesunden und hämatologisch erkrankten Personen
 Die Rolle von KIR- und HLA-Klasse I-Polymorphismen für die Funktion von Natürlichen Killerzellen in gesunden und hämatologisch erkrankten Personen Kumulative Inaugural-Dissertation zur Erlangung des Doktorgrades
Die Rolle von KIR- und HLA-Klasse I-Polymorphismen für die Funktion von Natürlichen Killerzellen in gesunden und hämatologisch erkrankten Personen Kumulative Inaugural-Dissertation zur Erlangung des Doktorgrades
Grundlagen der Immunologie
 Grundlagen der Immunologie 16. Vorlesung Immunglobulinvermittelte humorale Effektormechanismen Haupstadien der adaptiven Immunantwort Antigenerkennung Aktivierung, Differenzierung Effektorfunktionen Humorale
Grundlagen der Immunologie 16. Vorlesung Immunglobulinvermittelte humorale Effektormechanismen Haupstadien der adaptiven Immunantwort Antigenerkennung Aktivierung, Differenzierung Effektorfunktionen Humorale
Vertiefendes Seminar zur Vorlesung Biochemie
 Vertiefendes Seminar zur Vorlesung Biochemie 31.10.2014 Proteine: Struktur Gerhild van Echten-Deckert Fon. +49-228-732703 Homepage: http://www.limes-institut-bonn.de/forschung/arbeitsgruppen/unit-3/abteilung-van-echten-deckert/abt-van-echten-deckert-startseite/
Vertiefendes Seminar zur Vorlesung Biochemie 31.10.2014 Proteine: Struktur Gerhild van Echten-Deckert Fon. +49-228-732703 Homepage: http://www.limes-institut-bonn.de/forschung/arbeitsgruppen/unit-3/abteilung-van-echten-deckert/abt-van-echten-deckert-startseite/
B-Zell-vermittelte Immunantwort
 Rolle der humoralen Immunantwort B-Zell-vermittelte Immunantwort Extrazelluläre Erreger: humorale Immunantwort AK Plasmazellen: B-Effektorzellen Reifung der B-Zellen: Knochenmark reife, naive B-Zellen
Rolle der humoralen Immunantwort B-Zell-vermittelte Immunantwort Extrazelluläre Erreger: humorale Immunantwort AK Plasmazellen: B-Effektorzellen Reifung der B-Zellen: Knochenmark reife, naive B-Zellen
Chemokin-Rezeptor-Modulation während der T-Zell-Aktivierung: Untersuchung zur Funktion der GTPase Ras
 Aus dem Institut für Immunologie der Universität Heidelberg Geschäftsführender Direktor: Herr Prof. Dr. med. Stefan Meuer Chemokin-Rezeptor-Modulation während der T-Zell-Aktivierung: Untersuchung zur Funktion
Aus dem Institut für Immunologie der Universität Heidelberg Geschäftsführender Direktor: Herr Prof. Dr. med. Stefan Meuer Chemokin-Rezeptor-Modulation während der T-Zell-Aktivierung: Untersuchung zur Funktion
Matthias Birnstiel. Allergien. Modul. Medizinisch wissenschaftlicher Lehrgang CHRISANA. Wissenschaftliche Lehrmittel, Medien, Aus- und Weiterbildung
 Matthias Birnstiel Modul Allergien Medizinisch wissenschaftlicher Lehrgang CHRISANA Wissenschaftliche Lehrmittel, Medien, Aus- und Weiterbildung Inhaltsverzeichnis des Moduls Allergien Immunsystem und
Matthias Birnstiel Modul Allergien Medizinisch wissenschaftlicher Lehrgang CHRISANA Wissenschaftliche Lehrmittel, Medien, Aus- und Weiterbildung Inhaltsverzeichnis des Moduls Allergien Immunsystem und
3 Ergebnisse. 3.1 Die Speckling-Domäne von Wt1 umfaßt die Aminosäuren
 3 Ergebnisse 3.1 Die Speckling-Domäne von Wt1 umfaßt die Aminosäuren 76-120 Die Existenz verschiedener Isoformen von WT1 ist unter anderem auf die Verwendung einer für die Aminosäuren KTS kodierenden alternativen
3 Ergebnisse 3.1 Die Speckling-Domäne von Wt1 umfaßt die Aminosäuren 76-120 Die Existenz verschiedener Isoformen von WT1 ist unter anderem auf die Verwendung einer für die Aminosäuren KTS kodierenden alternativen
8. Translation. Konzepte: Translation benötigt trnas und Ribosomen. Genetischer Code. Initiation - Elongation - Termination
 8. Translation Konzepte: Translation benötigt trnas und Ribosomen Genetischer Code Initiation - Elongation - Termination 1. Welche Typen von RNAs gibt es und welches sind ihre Funktionen? mouse human bacteria
8. Translation Konzepte: Translation benötigt trnas und Ribosomen Genetischer Code Initiation - Elongation - Termination 1. Welche Typen von RNAs gibt es und welches sind ihre Funktionen? mouse human bacteria
Aufgabe 6 (Quartärstruktur)
") Aufgabe 6 (Quartärstruktur) Fragestellung Folgende Fragestellungen sollen beim Hämocyanin-Hexamer von Limulus polyphemus (1lla_Hexamer.pdb) untersucht werden: - Welche Symmetrien sind erkennbar? - Wenn
Aufgabe 6 (Quartärstruktur) Fragestellung Folgende Fragestellungen sollen beim Hämocyanin-Hexamer von Limulus polyphemus (1lla_Hexamer.pdb) untersucht werden: - Welche Symmetrien sind erkennbar? - Wenn
Alien Invasion I. Univ.-Prof. Dr. Albert Duschl
 Alien Invasion I Univ.-Prof. Dr. Albert Duschl Bakterien und wir Bakterien sind ein normaler und notwendiger Teil unserer Umwelt. Unser Körper enthält 10 14 Bakterien, aber nur 10 13 Eukaryontenzellen.
Alien Invasion I Univ.-Prof. Dr. Albert Duschl Bakterien und wir Bakterien sind ein normaler und notwendiger Teil unserer Umwelt. Unser Körper enthält 10 14 Bakterien, aber nur 10 13 Eukaryontenzellen.
Antigen Receptors and Accessory Molecules of T Lymphocytes
 Immunologie II Antigen Receptors and Accessory Molecules of T Lymphocytes Chapter 7 - Cellular and Molecular Immunology, Abbas-Lichtman-Pillai 6th edition Leslie Saurer Institut für Pathologie (Prof. Christoph
Immunologie II Antigen Receptors and Accessory Molecules of T Lymphocytes Chapter 7 - Cellular and Molecular Immunology, Abbas-Lichtman-Pillai 6th edition Leslie Saurer Institut für Pathologie (Prof. Christoph
Aminosäuren. Seitenkette. -Kohlenstoffatom. Karboxilgruppe. Aminogruppe
 Proteine Aminosäuren 16 Seitenkette -Kohlenstoffatom Aminogruppe Karboxilgruppe Die Gruppen der Aminosäuren 17 Bildung der Peptidbindung Die strukturellen Ebenen der Proteine 18 Primär Struktur Aminosäuer
Proteine Aminosäuren 16 Seitenkette -Kohlenstoffatom Aminogruppe Karboxilgruppe Die Gruppen der Aminosäuren 17 Bildung der Peptidbindung Die strukturellen Ebenen der Proteine 18 Primär Struktur Aminosäuer
Hämophilie Symposium, März , Reitter Sylvia
 Hämophilie Symposium, März 2010 FVIII-Gen liegt auf Xq28 (langer Arm des X-Chromosoms) x Hämophilie Erbgang I Hämophilie Erbgang II FVIII-Gen besteht aus 26 Exons mit 186 Kilobasenpaaren (kb); Exon 14
Hämophilie Symposium, März 2010 FVIII-Gen liegt auf Xq28 (langer Arm des X-Chromosoms) x Hämophilie Erbgang I Hämophilie Erbgang II FVIII-Gen besteht aus 26 Exons mit 186 Kilobasenpaaren (kb); Exon 14
7. Fragentyp A Wozu benötigt der reife Erythrozyt Glucose? A) zur Hämsynthese aus Succinyl-CoA B ) zur Herstellung von Ketonkörpern C) zur Bildung von
 zur Hämsynthese aus Succinyl-CoA B ) zur Herstellung von Ketonkörpern C) zur Bildung von") 1. Fragertyp D Welche der folgenden Aussagen über die Superfamilie der Immunglobuline trifft/treffen zu 1. Antikörpermoleküle gehören dazu 2. MHC- Moleküle gehören dazu 3. T-Zellrezeptoren gehören dazu
1. Fragertyp D Welche der folgenden Aussagen über die Superfamilie der Immunglobuline trifft/treffen zu 1. Antikörpermoleküle gehören dazu 2. MHC- Moleküle gehören dazu 3. T-Zellrezeptoren gehören dazu
Komponenten und Aufbau des Immunsystems Initiation von Immunantworten. lymphatische Organe. Erkennungsmechanismen. Lymphozytenentwicklung
 Komponenten und Aufbau des Immunsystems Initiation von Immunantworten lymphatische Organe Erkennungsmechanismen Lymphozytenentwicklung Entstehung und Verlauf adaptiver Immunantworten 159 Basale Aufgabe
Komponenten und Aufbau des Immunsystems Initiation von Immunantworten lymphatische Organe Erkennungsmechanismen Lymphozytenentwicklung Entstehung und Verlauf adaptiver Immunantworten 159 Basale Aufgabe
The Arabidopsis F-box protein TIR1 is an auxin receptor. Von Stefan Kepinski & Ottoline Leyser
 The Arabidopsis F-box protein TIR1 is an auxin receptor Von Stefan Kepinski & Ottoline Leyser Bekanntes Modell Was war bekannt? In der Zwischenzeit gefunden: - ABP1 kann große Mengen Auxin binden und ist
The Arabidopsis F-box protein TIR1 is an auxin receptor Von Stefan Kepinski & Ottoline Leyser Bekanntes Modell Was war bekannt? In der Zwischenzeit gefunden: - ABP1 kann große Mengen Auxin binden und ist
Hemmung der Enzym-Aktivität
 Enzym - Inhibitoren Wie wirkt Penicillin? Wie wirkt Aspirin? Welche Rolle spielt Methotrexat in der Chemotherapie? Welche Wirkstoffe werden gegen HIV entwickelt? Hemmung der Enzym-Aktivität Substrat Kompetitiver
Enzym - Inhibitoren Wie wirkt Penicillin? Wie wirkt Aspirin? Welche Rolle spielt Methotrexat in der Chemotherapie? Welche Wirkstoffe werden gegen HIV entwickelt? Hemmung der Enzym-Aktivität Substrat Kompetitiver
Natürliche Killerzellen. Welche Rolle spielen Sie in der Immunabwehr?
 6. Februar 2013 Natürliche Killerzellen Welche Rolle spielen Sie in der Immunabwehr? Dr. med. Volker von Baehr Institut für Medizinische Diagnostik Berlin, Nicolaistraße 22, 12247 Berlin +49 3077001-220,
6. Februar 2013 Natürliche Killerzellen Welche Rolle spielen Sie in der Immunabwehr? Dr. med. Volker von Baehr Institut für Medizinische Diagnostik Berlin, Nicolaistraße 22, 12247 Berlin +49 3077001-220,
Grundlagen der Immunologie
 Grundlagen der Immunologie 12. Vorlesung Das Komplementsystem Humorale Effektor-Mechanismen Immungloulin-vermittelte spezifische Komplement-vermittelte - angeorene Entdeckung des Komplementsystems 1890:
Grundlagen der Immunologie 12. Vorlesung Das Komplementsystem Humorale Effektor-Mechanismen Immungloulin-vermittelte spezifische Komplement-vermittelte - angeorene Entdeckung des Komplementsystems 1890:
Aminosäuren - Proteine
 Aminosäuren - Proteine ÜBERBLICK D.Pflumm KSR / MSE Aminosäuren Überblick Allgemeine Formel für das Grundgerüst einer Aminosäure Carboxylgruppe: R-COOH O Aminogruppe: R-NH 2 einzelnes C-Atom (α-c-atom)
Aminosäuren - Proteine ÜBERBLICK D.Pflumm KSR / MSE Aminosäuren Überblick Allgemeine Formel für das Grundgerüst einer Aminosäure Carboxylgruppe: R-COOH O Aminogruppe: R-NH 2 einzelnes C-Atom (α-c-atom)
Monoklonale Antikörper sind Antikörper, immunologisch aktive Proteine, die von einer auf einen einzigen B-Lymphozyten zurückgehenden Zelllinie
 Monoklonale AK Monoklonale Antikörper sind Antikörper, immunologisch aktive Proteine, die von einer auf einen einzigen B-Lymphozyten zurückgehenden Zelllinie (Zellklon) produziert werden und die sich gegen
Monoklonale AK Monoklonale Antikörper sind Antikörper, immunologisch aktive Proteine, die von einer auf einen einzigen B-Lymphozyten zurückgehenden Zelllinie (Zellklon) produziert werden und die sich gegen
Y Y. Natürliche (Angeborene) Immunität. Spezifische (erworbene) Immunität. Bakterien. Lymphozyt. T-Lymphozyten. EPITHELIALE Barriere PHAGOZYTEN
 Immunität. Spezifische (erworbene) Immunität. Bakterien. Lymphozyt. T-Lymphozyten. EPITHELIALE Barriere PHAGOZYTEN") Natürliche (Angeborene) Immunität Bakterien Spezifische (erworbene) Immunität B-Lymphozyten EPITHELIALE Barriere Knochenmark Y Y Y Y Y PHAGOZYTEN Stammzelle kleiner Lymphoblasten Effektor- Lymphozyt mechanismen
Natürliche (Angeborene) Immunität Bakterien Spezifische (erworbene) Immunität B-Lymphozyten EPITHELIALE Barriere Knochenmark Y Y Y Y Y PHAGOZYTEN Stammzelle kleiner Lymphoblasten Effektor- Lymphozyt mechanismen
Immunbiologie. Teil 4
 Teil 4 Funktion der lymphatischen Organe: - es werden drei Arten von lymphatischen Organen unterschieden: - primäre, sekundäre und tertiäre lymphatische Organe - die Organe unterscheiden sich in ihrer
Teil 4 Funktion der lymphatischen Organe: - es werden drei Arten von lymphatischen Organen unterschieden: - primäre, sekundäre und tertiäre lymphatische Organe - die Organe unterscheiden sich in ihrer
Eukaryontische DNA-Bindedomänen
 1. Viele eukaryotische (und auch prokaryotische) Transkriptionsfaktoren besitzen eine DNA-bindende Domäne, die an eine ganz bestimmte DNA- Sequenz binden kann. Aufgrund von Ähnlichkeiten in der Struktur
1. Viele eukaryotische (und auch prokaryotische) Transkriptionsfaktoren besitzen eine DNA-bindende Domäne, die an eine ganz bestimmte DNA- Sequenz binden kann. Aufgrund von Ähnlichkeiten in der Struktur
Angeborene und erworbene Immunantwort
 Molekulare Mechanismen der Pathogenese bei Infektionskrankheiten Angeborene und erworbene Immunantwort Hans-Georg Kräusslich Abteilung Virologie, Hygiene Institut INF 324, 4.OG http://www.virology-heidelberg.de
Molekulare Mechanismen der Pathogenese bei Infektionskrankheiten Angeborene und erworbene Immunantwort Hans-Georg Kräusslich Abteilung Virologie, Hygiene Institut INF 324, 4.OG http://www.virology-heidelberg.de
Pharmazeutische Biologie Grundlagen der Biochemie
 Pharmazeutische Biologie Grundlagen der Biochemie Prof. Dr. Theo Dingermann Institut für Pharmazeutische Biologie Goethe-Universität Frankfurt Dingermann@em.uni-frankfurt.de Aminosäure... chirale Moleküle
Pharmazeutische Biologie Grundlagen der Biochemie Prof. Dr. Theo Dingermann Institut für Pharmazeutische Biologie Goethe-Universität Frankfurt Dingermann@em.uni-frankfurt.de Aminosäure... chirale Moleküle
Stammbaum der Photorezeptoren
 Das Farbensehen der Vögel, Arbeitsmaterial 2 - Evolution des Farbensehens Stammbaum der Photorezeptoren AB 2-1 Arbeitsaufträge: 1. Vervollständigen Sie den Stammbaum der Photorezeptoren, indem Sie folgende
Das Farbensehen der Vögel, Arbeitsmaterial 2 - Evolution des Farbensehens Stammbaum der Photorezeptoren AB 2-1 Arbeitsaufträge: 1. Vervollständigen Sie den Stammbaum der Photorezeptoren, indem Sie folgende
INAUGURAL DISSERTATION. zur Erlangung der Doktorwürde der Naturwissenschaftlich-Mathematischen Gesamtfakultät der Ruprecht - Karls - Universität
 INAUGURAL DISSERTATION zur Erlangung der Doktorwürde der Naturwissenschaftlich-Mathematischen Gesamtfakultät der Ruprecht - Karls - Universität Heidelberg vorgelegt von Diplom- Biologin Bernadett Müller
INAUGURAL DISSERTATION zur Erlangung der Doktorwürde der Naturwissenschaftlich-Mathematischen Gesamtfakultät der Ruprecht - Karls - Universität Heidelberg vorgelegt von Diplom- Biologin Bernadett Müller
Um eine aktive Auseinandersetzung mit dem Thema Genomsequenzierung und die Diskussion dazu zu fördern, eignet sich die Methode Your DNA in a Box.
 Your DNA in a Box Gruppenaufgabe Um eine aktive Auseinandersetzung mit dem Thema Genomsequenzierung und die Diskussion dazu zu fördern, eignet sich die Methode Your DNA in a Box. Ziel der Übung ist es,
Your DNA in a Box Gruppenaufgabe Um eine aktive Auseinandersetzung mit dem Thema Genomsequenzierung und die Diskussion dazu zu fördern, eignet sich die Methode Your DNA in a Box. Ziel der Übung ist es,
und Reinstruktur Die Sekundärstruktur ist die Anordnung der Aminosäurenkette, wobei man in zwei Arten unterscheidet: o Faltblatt- oder β- Struktur
 Aufbau von Proteinen insbesondere unter Betrachtung der Primär-,, Sekundär-,, Tertiär-,, Quartär-,, Supersekundär- und Reinstruktur Primärstruktur Unter Primärstruktur versteht man in der Biochemie die
Aufbau von Proteinen insbesondere unter Betrachtung der Primär-,, Sekundär-,, Tertiär-,, Quartär-,, Supersekundär- und Reinstruktur Primärstruktur Unter Primärstruktur versteht man in der Biochemie die
Biologie I/B: Klassische und molekulare Genetik, molekulare Grundlagen der Entwicklung Theoretische Übungen SS 2016
 Biologie I/B: Klassische und molekulare Genetik, molekulare Grundlagen der Entwicklung Theoretische Übungen SS 2016 Fragen für die Übungsstunde 4 (20.06. 24.06.) Regulation der Transkription II, Translation
Biologie I/B: Klassische und molekulare Genetik, molekulare Grundlagen der Entwicklung Theoretische Übungen SS 2016 Fragen für die Übungsstunde 4 (20.06. 24.06.) Regulation der Transkription II, Translation
Wie erkennen die Immunzellen ein. Antigene. 2.1 Mustererkennungsrezeptoren
 2 Wie erkennen die Immunzellen ein Antigen? Im Zentrum der Abbildung 2.1 sieht man drei Bakterienstämme mit verschiedenen Antigenen auf ihrer Oberfläche. Leicht kann man zwei Typen von Oberflächenmolekülen
2 Wie erkennen die Immunzellen ein Antigen? Im Zentrum der Abbildung 2.1 sieht man drei Bakterienstämme mit verschiedenen Antigenen auf ihrer Oberfläche. Leicht kann man zwei Typen von Oberflächenmolekülen
Untersuchungen zur Zytokinexpression von Patienten mit fortgeschrittenen gynäkologischen Tumoren und Tumoren im Kopf-Hals-Bereich
 Aus der Universitätsklinik und Poliklinik für Strahlentherapie an der Martin-Luther-Universität Halle-Wittenberg (Komm.Direktorin: Frau PD Dr. med. Gabriele Hänsgen) Untersuchungen zur Zytokinexpression
Aus der Universitätsklinik und Poliklinik für Strahlentherapie an der Martin-Luther-Universität Halle-Wittenberg (Komm.Direktorin: Frau PD Dr. med. Gabriele Hänsgen) Untersuchungen zur Zytokinexpression
Molekulare Mechanismen der Signaltransduktion. 06 - Kartierung des AXR1 Gens + early auxin-induced genes Folien: http://tinyurl.
 Molekulare Mechanismen der Signaltransduktion 06 - Kartierung des AXR1 Gens + early auxin-induced genes Folien: http://tinyurl.com/modul-mms bisheriges Modell auxin auxin AXR1 auxin response AXR1 potentieller
Molekulare Mechanismen der Signaltransduktion 06 - Kartierung des AXR1 Gens + early auxin-induced genes Folien: http://tinyurl.com/modul-mms bisheriges Modell auxin auxin AXR1 auxin response AXR1 potentieller
Antikörper. Prof. Dr. Albert Duschl
 Antikörper Prof. Dr. Albert Duschl Proteins reloaded Mit keinem Protein werden Sie im Studium so viel zu tun bekommen wie mit Antikörpern. Antikörper (= Immunglobuline) sind Proteine des Immunsystems die
Antikörper Prof. Dr. Albert Duschl Proteins reloaded Mit keinem Protein werden Sie im Studium so viel zu tun bekommen wie mit Antikörpern. Antikörper (= Immunglobuline) sind Proteine des Immunsystems die
CD5 + B-Lymphozyten und Immunglobulin-Leichtketten bei HIV-Infizierten - Korrelation mit klinischen und immunologischen Parametern
 Aus der 2. Klinik für Innere Medizin des Städtischen Klinikums St. Georg Leipzig (Leiter: Prof. Dr. med. Bernhard R. Ruf) CD5 + B-Lymphozyten und Immunglobulin-Leichtketten bei HIV-Infizierten - Korrelation
Aus der 2. Klinik für Innere Medizin des Städtischen Klinikums St. Georg Leipzig (Leiter: Prof. Dr. med. Bernhard R. Ruf) CD5 + B-Lymphozyten und Immunglobulin-Leichtketten bei HIV-Infizierten - Korrelation
Das Komplementsystem. Effektorwirkungen. Der humorale Effektorapparat des angeborenen Immunsystems
 Das Komplementsystem Der humorale Effektorapparat des angeborenen Immunsystems Hitzeempfindlicher Bestandteil des Plasmas Hitzebehandlung (56 ºC): Inaktivierung der bakteriziden Wirkung des Plasmas Ergänzt
Das Komplementsystem Der humorale Effektorapparat des angeborenen Immunsystems Hitzeempfindlicher Bestandteil des Plasmas Hitzebehandlung (56 ºC): Inaktivierung der bakteriziden Wirkung des Plasmas Ergänzt
Funktion von Proteinen:
 Funktion von Proteinen: Transport und Speicherung Beispiel Myoglobin / Hämoglobin Myoglobin Hämoglobin Globin-Faltung ist konserviert Häm-Gruppe Cofaktoren, Coenzyme & Prosthetische Gruppen Cofaktor wird
Funktion von Proteinen: Transport und Speicherung Beispiel Myoglobin / Hämoglobin Myoglobin Hämoglobin Globin-Faltung ist konserviert Häm-Gruppe Cofaktoren, Coenzyme & Prosthetische Gruppen Cofaktor wird
In dieser Doktorarbeit wird eine rezeptorvermittelte Signalkaskade für Thrombin
 Diskussion -33-4. Diskussion In dieser Doktorarbeit wird eine rezeptorvermittelte Signalkaskade für Thrombin beschrieben, die zur Differenzierung von neonatalen glatten Gefäßmuskelzellen führt. Thrombin
Diskussion -33-4. Diskussion In dieser Doktorarbeit wird eine rezeptorvermittelte Signalkaskade für Thrombin beschrieben, die zur Differenzierung von neonatalen glatten Gefäßmuskelzellen führt. Thrombin
Alien Invasion II. Univ.-Prof. Dr. Albert Duschl
 Alien Invasion II Univ.-Prof. Dr. Albert Duschl Das Mantra RISK = HAZARD x EXPOSURE Das ist die grundlegende Formel für Risk Assessment. No hazard (commensal bacteria), high exposure = No risk High hazard,
Alien Invasion II Univ.-Prof. Dr. Albert Duschl Das Mantra RISK = HAZARD x EXPOSURE Das ist die grundlegende Formel für Risk Assessment. No hazard (commensal bacteria), high exposure = No risk High hazard,
Genregulation bei Eukaryoten II
 Genregulation bei Eukaryoten II Aktivierung und Repression der Transkription erfolgen durch Protein-Protein-Wechselwirkungen Protein-Protein-Wechselwirkungen spielen bei der Genregulation der Eukaryoten
Genregulation bei Eukaryoten II Aktivierung und Repression der Transkription erfolgen durch Protein-Protein-Wechselwirkungen Protein-Protein-Wechselwirkungen spielen bei der Genregulation der Eukaryoten
4.1. Herstellung, Aufreinigung und Nachweis eines CD33-spezifischen IgM-Antikörpers
 ERGEBNISSE 30 4. Ergebnisse 4.1. Herstellung, Aufreinigung und Nachweis eines CD33-spezifischen IgM-Antikörpers 4.1.1. Herstellung und Aufreinigung des Antikörpers CD33-spezifische IgM-Antikörper wurden
ERGEBNISSE 30 4. Ergebnisse 4.1. Herstellung, Aufreinigung und Nachweis eines CD33-spezifischen IgM-Antikörpers 4.1.1. Herstellung und Aufreinigung des Antikörpers CD33-spezifische IgM-Antikörper wurden
Posttranskriptionale RNA-Prozessierung
 Posttranskriptionale RNA-Prozessierung Spaltung + Modifikation G Q Spleissen + Editing U UUU Prozessierung einer prä-trna Eukaryotische messenger-rna Cap-Nukleotid am 5 -Ende Polyadenylierung am 3 -Ende
Posttranskriptionale RNA-Prozessierung Spaltung + Modifikation G Q Spleissen + Editing U UUU Prozessierung einer prä-trna Eukaryotische messenger-rna Cap-Nukleotid am 5 -Ende Polyadenylierung am 3 -Ende
Zelluläre Reproduktion: Zellzyklus. Regulation des Zellzyklus - Proliferation
 Zelluläre Reproduktion: Zellzyklus Regulation des Zellzyklus - Proliferation Alle Zellen entstehen durch Zellteilung Der Zellzyklus kann in vier Haupt-Phasen eingeteilt werden Interphase Zellwachstum;
Zelluläre Reproduktion: Zellzyklus Regulation des Zellzyklus - Proliferation Alle Zellen entstehen durch Zellteilung Der Zellzyklus kann in vier Haupt-Phasen eingeteilt werden Interphase Zellwachstum;
Abwehr I/2I. Seminar Block 8 WS 08/09. Dr. Mag. Krisztina Szalai
 Abwehr I/2I Seminar Block 8 WS 08/09 Dr. Mag. Krisztina Szalai krisztina.szalai@meduniwien.ac.at Der zelluläre Schenkel der natürlichen Abwehr Phagozytose: Polymorphkernige Granulozyten Neutrophile Monomorphkernige
Abwehr I/2I Seminar Block 8 WS 08/09 Dr. Mag. Krisztina Szalai krisztina.szalai@meduniwien.ac.at Der zelluläre Schenkel der natürlichen Abwehr Phagozytose: Polymorphkernige Granulozyten Neutrophile Monomorphkernige
Der Zellzyklus und mögliche Störungen durch chemische Substanzen
 Teil 2 Der Zellzyklus und mögliche Störungen durch chemische Substanzen Aplasie Hemmung der Zellzyklusprogression Gesteigerte oder unkontrollierte Zellzyklusprogression Hyperplasie Karzinogenese Proliferierende
Teil 2 Der Zellzyklus und mögliche Störungen durch chemische Substanzen Aplasie Hemmung der Zellzyklusprogression Gesteigerte oder unkontrollierte Zellzyklusprogression Hyperplasie Karzinogenese Proliferierende
Personalisierte Medizin
 Personalisierte Medizin Möglichkeiten und Grenzen Prof. Dr. Friedemann Horn Universität Leipzig, Institut für Klinische Immunologie, Molekulare Immunologie Fraunhofer Institut für Zelltherapie und Immunologie
Personalisierte Medizin Möglichkeiten und Grenzen Prof. Dr. Friedemann Horn Universität Leipzig, Institut für Klinische Immunologie, Molekulare Immunologie Fraunhofer Institut für Zelltherapie und Immunologie
Inhalt. Entdeckung und allgemeine Informationen. Klassifizierung. Genom Viren untypische Gene Tyrosyl-tRNA Synthetase. Ursprung von grossen DNA Viren
 Mimivirus Inhalt Entdeckung und allgemeine Informationen Klassifizierung Genom Viren untypische Gene Tyrosyl-tRNA Synthetase Ursprung von grossen DNA Viren Entstehung von Eukaryoten Entdeckung 1992 in
Mimivirus Inhalt Entdeckung und allgemeine Informationen Klassifizierung Genom Viren untypische Gene Tyrosyl-tRNA Synthetase Ursprung von grossen DNA Viren Entstehung von Eukaryoten Entdeckung 1992 in
Ergebnisse. I. Identifizierung intrazellulärer Interaktionspartner. Ergebnisse I. Identifizierung intrazellulärer Interaktionspartner
 Ergebnisse I. Identifizierung intrazellulärer Interaktionspartner Es sollten mithilfe des Yeast-Two-Hybrid-Systems Proteine identifiziert werden, die mit der zytoplasmatischen Domäne von CEACAM1-4L aus
Ergebnisse I. Identifizierung intrazellulärer Interaktionspartner Es sollten mithilfe des Yeast-Two-Hybrid-Systems Proteine identifiziert werden, die mit der zytoplasmatischen Domäne von CEACAM1-4L aus
Inaugural-Dissertation zur Erlangung der medizinischen Doktorwürde an der Charité - Universitätsmedizin Berlin
 Charité - Universitätsmedizin Berlin Campus Benjamin Franklin Aus der Medizinischen Klinik III Direktor: Prof. Dr. med. Eckhard Thiel Unterschiede in der T-Zell-Immunität gegen die Tumorantigene EpCAM,
Charité - Universitätsmedizin Berlin Campus Benjamin Franklin Aus der Medizinischen Klinik III Direktor: Prof. Dr. med. Eckhard Thiel Unterschiede in der T-Zell-Immunität gegen die Tumorantigene EpCAM,
1. Welche Auswirkungen auf die Expression des lac-operons haben die folgenden Mutationen:
 Übung 10 1. Welche Auswirkungen auf die Expression des lac-operons haben die folgenden Mutationen: a. Eine Mutation, die zur Expression eines Repressors führt, der nicht mehr an den Operator binden kann.
Übung 10 1. Welche Auswirkungen auf die Expression des lac-operons haben die folgenden Mutationen: a. Eine Mutation, die zur Expression eines Repressors führt, der nicht mehr an den Operator binden kann.
Traditionelle und innovative Impfstoffentwicklung
 Traditionelle und innovative Impfstoffentwicklung Reingard.grabherr@boku.ac.at Traditionelle Impfstoffentwicklung Traditionelle Impfstoffentwicklung Louis Pasteur in his laboratory, painting by A. Edelfeldt
Traditionelle und innovative Impfstoffentwicklung Reingard.grabherr@boku.ac.at Traditionelle Impfstoffentwicklung Traditionelle Impfstoffentwicklung Louis Pasteur in his laboratory, painting by A. Edelfeldt
Human Papilloma Virus
 Human Papilloma Virus Gliederung Allgemeine Informationen Virusstruktur Infektion Verschiedene Arten des Infektionsverlaufs nach Infizierung mit HPV Lebenszyklus des HPV Tumorinduktion Virusstruktur Papillomaviren
Human Papilloma Virus Gliederung Allgemeine Informationen Virusstruktur Infektion Verschiedene Arten des Infektionsverlaufs nach Infizierung mit HPV Lebenszyklus des HPV Tumorinduktion Virusstruktur Papillomaviren
Antikörperstruktur, -funktion und -affinitätsreifung
 Grundlagen der Immunologie 5. Semester - Dienstags 11.15 Uhr Ruhr-Universität Bochum, HMA 20 Antikörperstruktur, -funktion und -affinitätsreifung Albrecht Bufe www.ruhr-uni-bochum.de/homeexpneu Aktivierte
Grundlagen der Immunologie 5. Semester - Dienstags 11.15 Uhr Ruhr-Universität Bochum, HMA 20 Antikörperstruktur, -funktion und -affinitätsreifung Albrecht Bufe www.ruhr-uni-bochum.de/homeexpneu Aktivierte
Testfragen zur 1. Vorlesung in Biochemie
 Testfragen zur 1. Vorlesung in Biochemie 1. Nennen Sie die zentralen Komponenten des Zwei-Komponenten-Systems 2. Auf welche Aminosäurereste werden die Phosphatgruppen übertragen? 3. Was wird bei der Chemotaxis
Testfragen zur 1. Vorlesung in Biochemie 1. Nennen Sie die zentralen Komponenten des Zwei-Komponenten-Systems 2. Auf welche Aminosäurereste werden die Phosphatgruppen übertragen? 3. Was wird bei der Chemotaxis
Abbildung 1: Schematische Darstellung der Domänenorganisation von Survivin
 Abbildung 1: Schematische Darstellung der Domänenorganisation von Survivin Survivin weist eine N-terminale BIR-Domäne (grün) und eine C-terminale α-helix (blau) auf. Das intrinsische nuclear export signal
Abbildung 1: Schematische Darstellung der Domänenorganisation von Survivin Survivin weist eine N-terminale BIR-Domäne (grün) und eine C-terminale α-helix (blau) auf. Das intrinsische nuclear export signal
Eukaryotische messenger-rna
 Eukaryotische messenger-rna Cap-Nukleotid am 5 -Ende Polyadenylierung am 3 -Ende u.u. nicht-codierende Bereiche (Introns) Spleißen von prä-mrna Viele Protein-codierende Gene in Eukaryoten sind durch nicht-codierende
Eukaryotische messenger-rna Cap-Nukleotid am 5 -Ende Polyadenylierung am 3 -Ende u.u. nicht-codierende Bereiche (Introns) Spleißen von prä-mrna Viele Protein-codierende Gene in Eukaryoten sind durch nicht-codierende
Organisation und Evolution des Genoms
 Organisation und Evolution des Genoms Organisation und Evolution des Genoms Definition Genom: vollständige DNA-Sequenz eines Organismus I. Einfachstes Genom: Prokaryoten Zwei Gruppen, evolutionär unterschiedlicher
Organisation und Evolution des Genoms Organisation und Evolution des Genoms Definition Genom: vollständige DNA-Sequenz eines Organismus I. Einfachstes Genom: Prokaryoten Zwei Gruppen, evolutionär unterschiedlicher
Expression und immunmodulatorische Funktion von HLA-G und seinen verkürzten Isoformen. in Tumorzellinien
 Expression und immunmodulatorische Funktion von HLA-G und seinen verkürzten Isoformen in Tumorzellinien Dissertation zur Erlangung des Doktorgrades der Naturwissenschaften an der Fakultät für Biologie
Expression und immunmodulatorische Funktion von HLA-G und seinen verkürzten Isoformen in Tumorzellinien Dissertation zur Erlangung des Doktorgrades der Naturwissenschaften an der Fakultät für Biologie
Protein-Protein Bindungsstellen. Lennart Heinzerling
 Protein-Protein Bindungsstellen Lennart Heinzerling 1 Worum geht es in den nächsten 45 Minuten? Auffinden von Protein- Protein Komplexen aus einer großen Menge potentieller Komplexe z.b. für -Interaction
Protein-Protein Bindungsstellen Lennart Heinzerling 1 Worum geht es in den nächsten 45 Minuten? Auffinden von Protein- Protein Komplexen aus einer großen Menge potentieller Komplexe z.b. für -Interaction
SOCS-Proteine: Die molekulare Feuerwehr bei Entzündungen
 SOCS-Proteine: Die molekulare Feuerwehr bei Entzündungen Edgar Sawatzky, Meike Egert und Maria Stec Es ist Winter. Wieder einmal ist die Hochsaison für Erkältungen angebrochen und infektiöse Viren und
SOCS-Proteine: Die molekulare Feuerwehr bei Entzündungen Edgar Sawatzky, Meike Egert und Maria Stec Es ist Winter. Wieder einmal ist die Hochsaison für Erkältungen angebrochen und infektiöse Viren und
Bioinformatik: Schnittstelle zwischen Informatik und Life-Science
 Bioinformatik: Schnittstelle zwischen Informatik und Life-Science Andreas Zendler (PD Dr.rer.nat.Dr.phil.) GI / GChACM 12. ovember 2001 Inhaltsübersicht I. Einführung II. Bioinformatik III. Industrial
Bioinformatik: Schnittstelle zwischen Informatik und Life-Science Andreas Zendler (PD Dr.rer.nat.Dr.phil.) GI / GChACM 12. ovember 2001 Inhaltsübersicht I. Einführung II. Bioinformatik III. Industrial
Untersuchung der Expression und Regulation der KIR Gene im Rahmen der Differenzierung humaner Natürlicher Killerzellen
 Untersuchung der Expression und Regulation der KIR Gene im Rahmen der Differenzierung humaner Natürlicher Killerzellen Inaugural-Dissertation zur Erlangung des Doktorgrades der der Mathematisch-Naturwissenschaftlichen
Untersuchung der Expression und Regulation der KIR Gene im Rahmen der Differenzierung humaner Natürlicher Killerzellen Inaugural-Dissertation zur Erlangung des Doktorgrades der der Mathematisch-Naturwissenschaftlichen
Enzyme SPF BCH am
 Enzyme Inhaltsverzeichnis Ihr kennt den Aufbau von Proteinen (mit vier Strukturelementen) und kennt die Kräfte, welche den Aufbau und die Funktion von Enzymen bestimmen... 3 Ihr versteht die Einteilung
Enzyme Inhaltsverzeichnis Ihr kennt den Aufbau von Proteinen (mit vier Strukturelementen) und kennt die Kräfte, welche den Aufbau und die Funktion von Enzymen bestimmen... 3 Ihr versteht die Einteilung
Th1-Th2-Zytokine bei entzündlicher Herzmuskelerkrankung
 Aus der medizinischen Klinik II, Abteilung für Kardiologie und Pulmonologie des Universitätsklinikums Benjamin Franklin der Freien Universität Berlin Direktor: Univ.-Prof. Dr. med. H.-P. Schultheiss Th1-Th2-Zytokine
Aus der medizinischen Klinik II, Abteilung für Kardiologie und Pulmonologie des Universitätsklinikums Benjamin Franklin der Freien Universität Berlin Direktor: Univ.-Prof. Dr. med. H.-P. Schultheiss Th1-Th2-Zytokine
5 Zusammenfassung. Zusammenfassung 76
 Zusammenfassung 76 5 Zusammenfassung 1. Aminosäuren sind als Bausteine der Proteine essentiell zur Aufrechterhaltung vitaler Prozesse. Eine besondere Rolle im Zellmetabolismus der Haut spielen dabei kationische
Zusammenfassung 76 5 Zusammenfassung 1. Aminosäuren sind als Bausteine der Proteine essentiell zur Aufrechterhaltung vitaler Prozesse. Eine besondere Rolle im Zellmetabolismus der Haut spielen dabei kationische
Stand von letzter Woche
 RUB ECR1 AXR1 Stand von letzter Woche E2 U? E1-like AXR1 Repressor ARF1 Proteasom AuxRE Repressor wird sehr schnell abgebaut notwendig für Auxinantwort evtl. Substrat für SCF Identifikation des SCF-Ubiquitin
RUB ECR1 AXR1 Stand von letzter Woche E2 U? E1-like AXR1 Repressor ARF1 Proteasom AuxRE Repressor wird sehr schnell abgebaut notwendig für Auxinantwort evtl. Substrat für SCF Identifikation des SCF-Ubiquitin
β2-microglobulin deficient mice lack CD4-8+cytolytic T cells
 β2-microglobulin deficient mice lack CD4-8+cytolytic T cells Mäuse mit einem Knock-out bezüglich ß-Microglobulin sind nicht in der Lage CD4-8+ cytotoxische T-Zellen zu bilden Nature,Vol 344, 19. April
β2-microglobulin deficient mice lack CD4-8+cytolytic T cells Mäuse mit einem Knock-out bezüglich ß-Microglobulin sind nicht in der Lage CD4-8+ cytotoxische T-Zellen zu bilden Nature,Vol 344, 19. April
Genregulation bei Eukaryoten
 Genregulation bei Eukaryoten 1. Allgemeines über Genregulation 2. Aufbau der DNA 3. Enhancer 4. Aktivierung und Repression 5. System, das Östrogene wahrnimmt und auf sie anspricht - DNA- Bindungsdomäne
Genregulation bei Eukaryoten 1. Allgemeines über Genregulation 2. Aufbau der DNA 3. Enhancer 4. Aktivierung und Repression 5. System, das Östrogene wahrnimmt und auf sie anspricht - DNA- Bindungsdomäne
Kohlenhydrate Aminosäuren Peptide
 Kohlenhydrate Aminosäuren Peptide Kohlenhydrate Kohlenhydrate = Saccharide Sie stellen zusammen mit Fetten und Proteinen den quantitativ größten verwertbaren (z.b. Stärke) und nicht-verwertbaren (Ballaststoffe)
Kohlenhydrate Aminosäuren Peptide Kohlenhydrate Kohlenhydrate = Saccharide Sie stellen zusammen mit Fetten und Proteinen den quantitativ größten verwertbaren (z.b. Stärke) und nicht-verwertbaren (Ballaststoffe)
Pathohysiologie und Androgendeprivationtstherapie(ADT) des kastrationsresistenten Prostatakarzinoms
 des kastrationsresistenten Prostatakarzinoms") Pathohysiologie und Androgendeprivationtstherapie(ADT) des kastrationsresistenten Prostatakarzinoms KASTRATIONSRESISTENTES PROSTATAKARZINOM (CRPC) Patienten sprechen im Allgemeinen gut auf eine ADT an.
Pathohysiologie und Androgendeprivationtstherapie(ADT) des kastrationsresistenten Prostatakarzinoms KASTRATIONSRESISTENTES PROSTATAKARZINOM (CRPC) Patienten sprechen im Allgemeinen gut auf eine ADT an.
Komponenten und Aufbau des Immunsystems Initiation von Immunantworten. lymphatische Organe. Erkennungsmechanismen. Lymphozytenentwicklung
 Komponenten und Aufbau des Immunsystems Initiation von Immunantworten lymphatische Organe Erkennungsmechanismen Lymphozytenentwicklung Entstehung und Verlauf adaptiver Immunantworten 1 Dendritische Zellen
Komponenten und Aufbau des Immunsystems Initiation von Immunantworten lymphatische Organe Erkennungsmechanismen Lymphozytenentwicklung Entstehung und Verlauf adaptiver Immunantworten 1 Dendritische Zellen
Inhalt 1 Das Immunsystem Rezeptoren des Immunsystems
 Inhalt 1 Das Immunsystem 1.1 Bedeutung des Immunsystems..................................... 1 1.2 Das Immunsystem unterscheidet zwischen körpereigen und körperfremd.................................................
Inhalt 1 Das Immunsystem 1.1 Bedeutung des Immunsystems..................................... 1 1.2 Das Immunsystem unterscheidet zwischen körpereigen und körperfremd.................................................
Diskussion 62 6 DISKUSSION
 Diskussion 62 6 DISKUSSION Kartierungsergebnisse Sowohl die Analyse von Deletionssyndromen, wie dem WAGR-Syndrom, als auch Beobachtungen von Allelverlusten der Region 11p13 14.1 in verschiedenen Tumoren
Diskussion 62 6 DISKUSSION Kartierungsergebnisse Sowohl die Analyse von Deletionssyndromen, wie dem WAGR-Syndrom, als auch Beobachtungen von Allelverlusten der Region 11p13 14.1 in verschiedenen Tumoren
Adoptive Immuntherapie dendritische Zellen, Killerzellen, T-Zellen
 Adoptive Immuntherapie dendritische Zellen, Killerzellen, T-Zellen Priv. Doz. Dr. med. Torsten Tonn Institut für Transfusionsmedizin und Immunhämatologie Johann Wolfgang Goethe Universitätsklinikum Frankfurt
Adoptive Immuntherapie dendritische Zellen, Killerzellen, T-Zellen Priv. Doz. Dr. med. Torsten Tonn Institut für Transfusionsmedizin und Immunhämatologie Johann Wolfgang Goethe Universitätsklinikum Frankfurt
1 Einleitung. 1.1 Rezeptortyrosinkinasen
 Einleitung 1 1 Einleitung In lebenden Zellen findet eine Vielzahl biochemischer Prozesse statt, die einer ständigen Kontrolle und Regulation bedürfen. Um intrazelluläre Prozesse aufeinander abstimmen zu
Einleitung 1 1 Einleitung In lebenden Zellen findet eine Vielzahl biochemischer Prozesse statt, die einer ständigen Kontrolle und Regulation bedürfen. Um intrazelluläre Prozesse aufeinander abstimmen zu